Copyright © 2000, American Society for Microbiology. All Rights Reserved.
Sequence Analysis of the
ank
Gene of Granulocytic Ehrlichiae
ROBERT F. MASSUNG,1* JESSICA H. OWENS,1DAVID ROSS,1KURT D. REED,2MIROSLAV PETROVEC,3
ANNELI BJOERSDORFF,4RICHARD T. COUGHLIN,5GERALD A. BELTZ,5ANDCHERYL I. MURPHY5
Division of Viral and Rickettsial Diseases, Centers for Disease Control and Prevention, Atlanta, Georgia1; Marshfield Clinic and Marshfield Medical Research Foundation, Marshfield, Wisconsin2; Institute of Microbiology and Immunology, Medical Faculty,
University of Ljubljana, Ljubljana, Slovenia3; Department of Clinical Microbiology, Kalmar County Hospital, Kalmar, Sweden4; and Aquila Biopharmaceuticals, Inc., Framingham, Massachusetts5
Received 3 January 2000/Returned for modification 14 March 2000/Accepted 28 April 2000
Theankgene of the agent of human granulocytic ehrlichiosis (HGE) codes for a protein with a predicted
molecular size of 131.2 kDa that is recognized by serum from both dogs and humans infected with granulocytic ehrlichiae. As part of an effort to assess the phylogenetic relatedness of granulocytic ehrlichiae from different
geographic regions and in different host species, theankgene was PCR amplified and sequenced from a variety
of sources. These included 10 blood specimens from patients with confirmed human granulocytic ehrlichiosis (three from New York, four from Wisconsin, two from Slovenia, and one from Sweden). Also examined was a
canine granulocytic ehrlichia sample obtained from Minnesota, Ehrlichia equi from California, Ehrlichia
phagocytophilafrom Sweden, and the granulocytic ehrlichia isolate USG3. The sequences showed a high level of homology (>95.5% identity), with the lowest homology occurring between a New York HGE agent and the
SwedishE. phagocytophila. Several 3-bp deletions and a variable number of 51- and 81-bp direct repeats were
noted. Although the North American HGE sequences showed the highest conservation (>98.1% identity), phylogenetic analyses indicated that these samples represent two separate clades, one including the three New York HGE samples and the USG3 strain and another with the Wisconsin HGE and Minnesota canine sequences. Two of the New York samples and the USG3 strain showed 100% identity over the entire 3,696-bp product. Likewise, three of the Wisconsin human samples and the Minnesota dog sample were identical (3,693
bp). Whereas phylogenetic analysis showed that theE. equisequence was most closely related to the Upper
Midwest samples, analysis of the repeat structures showed it to be more similar to the European samples.
Overall, the genetic analysis based on theankgene showed that the granulocytic ehrlichiae are closely related,
appear to infect multiple species, and can be grouped into at least three different clades, two North American and one European.
The members of the genusEhrlichiaare obligate, intracel-lular bacteria within the orderRickettsiales. Although ehrlichia infections of veterinary importance were first described in 1935, the first case of human ehrlichiosis in the United States was reported in 1987 (17). The human pathogen was subse-quently identified asEhrlichia chaffeensis(1). In 1994, a second ehrlichia infection of humans was reported and has been re-ferred to as human granulocytic ehrlichiosis (HGE) owing to the proclivity of the agent to infect neutrophils (5). The ma-jority of HGE cases have been diagnosed in the Northeastern and Upper Midwestern areas of the United States, although a limited number of cases have been reported in Europe and in northern California (2, 4, 9–12, 22, 26).
The genusEhrlichiahas been divided into three genogroups based on analysis of 16S rRNA sequences, and the HGE agent is a member of theEhrlichia phagocytophilagenogroup (29).E. phagocytophila is the etiologic agent of tick-borne fever in ruminants in Europe, and the E. phagocytophila genogroup also includes Ehrlichia equi, the agent of equine granulocytic ehrlichiosis. These three granulocytic ehrlichiae (GE) are very closely related based on numerous criteria including the high degree of homology of the 16S rRNA and groE DNA se-quences and may represent strains of a single species (5, 14, 25). However, the analysis of genetic elements less conserved than the 16S andgroEsequences is needed to accurately assess
the phylogenetic relationship of these agents and the degree of variability at the subspecies level. Recently, several genes that may encode structural proteins were identified from an HGE agent (USG3 strain) expression library and were recognized by serum from humans and canines infected with GE (24). One of these is referred to as theankgene, owing to a series of repeats within the predicted 131.2-kDa protein product that are similar to the repeats within the human erythrocyte ankyrin protein (24). We have PCR amplified and sequenced the completeank gene open reading frame and 28 bp 5⬘of the initiating methi-onine and 13 bp downstream from the termination codon. Samples examined were the HGE agent from 10 confirmed cases (3 from New York, 4 from Wisconsin, 1 from Sweden, and 2 from Slovenia), a canine granulocytic ehrlichiosis agent from a Minnesota dog,E. equi, andE. phagocytophilafrom a Swedish cow.
MATERIALS AND METHODS
Samples and sample preparation.All samples, except for the USG3 strain,
were obtained as EDTA blood specimens. Human samples were from HGE cases confirmed by seroconversion or by amplification and DNA sequencing of the 16S rRNA gene. The Swedish sample was from a 32-year-old previously healthy woman who presented at an outpatient clinic in Ronneby (Blekinge County, southern Sweden), with a 5-day history of fever (38 to 39°C), chills, headache, and myalgia. The patient worked part-time as a farmer, and she had recently been involved in the hunting and handling of a slaughtered roebuck. She also reported that she had sustained five or six tick bites during the month prior to admission. The patient’s acute-phase blood showed a reciprocal titer of 1:160 and was PCR positive when tested at the Centers for Disease Control and Prevention using the 16S rRNA gene as the target (data not shown).
DNA was extracted directly from the North American and Swedish blood samples by using a QIAamp blood extraction kit (Qiagen, Chatsworth, Calif.). * Corresponding author. Mailing address: Centers for Disease
Con-trol and Prevention, 1600 Clifton Rd., MS G-13, Atlanta, GA 30333. Phone: (404) 639-1082. Fax: (404) 639-4436. E-mail: [email protected].
2917
on May 15, 2020 by guest
http://jcm.asm.org/
The protocol followed was that suggested by the manufacturer. Briefly, detergent lysis was performed in the presence of proteinase K for 10 min at 70°C. The lysed material was applied to a spin column containing a silica gel-based membrane and washed twice. Purified DNA was eluted from the columns in 200l of Tris (10 mM, pH 8.0) and stored at 4°C until used as template for PCR amplification. DNA was extracted from the Slovenian EDTA blood samples using a modi-fication of the manufacturer’s protocol for the QIAamp tissue kit (Qiagen) as previously described (22). The USG3 strain preparation and the determination of the sequence of theank gene have been described previously (GenBank accession no. AF020521) (24). Horse blood infected withE. equiwas kindly provided by Richard Corstevet (Louisiana State University).
PCR analysis.Both nested and direct PCR protocols were used. Direct PCR
amplifications consisted of 40 cycles with each cycle including a 30-s denaturation at 94°C, a 30-s annealing at 55°C, and a 1-min extension at 72°C. The 40 cycles were preceded by a 2-min denaturation at 95°C and followed by a 5-min exten-sion at 72°C. PCR amplifications were performed in a Perkin-Elmer 9600 ther-mal cycler (Perkin-Elmer, Applied Biosystems Division, Foster City, Calif.), and reagents were from the GeneAmp PCR Kit with AmpliTaq DNA polymerase (Perkin-Elmer). Primary reactions used 5l of purified DNA as template in a total volume of 50l. Amplifications contained 200M (each) deoxynucleotide triphosphates (dATP, dCTP, dGTP, and dTTP), 1.25 U ofTaqpolymerase, and 0.5M (each) primer. Reaction products were subsequently maintained at 4°C until analyzed by agarose gel electrophoresis or used as template for nested reactions.
Nested amplifications used 1l of the primary PCR product as template in a total volume of 50l. Each nested amplification contained 200M (each) deoxynucleoside triphosphates (dATP, dCTP, dGTP, and dTTP), 1.25 U ofTaq polymerase, and 0.2M (each) primer. Nested cycling conditions were as de-scribed for the primary amplification, except that 30 cycles were used. Reaction mixtures were subsequently maintained at 4°C until analyzed by agarose gel electrophoresis or purified for DNA sequencing.
Primers used for PCR amplification and sequencing were as follows: 1F, ATGTTACGCTGTAATAGCATGGAC; 1R, TGCCCCAGCTTCTACAAC AC; 2F1, CTGATGTAAATGCGTCTCCA; 2R1, ACCATTTGCTTCTTGAG GAG; 3F, GTCTCGAAAGCATTTGTCAAAC; 3R, TTTCTCCCTTAGATGA CGCC; 4F1, GCTGCAATTACTTCCGAGGC; 4R1, GCGACCTCCTTTTAC AGACTTAG; U3, GAGGGCAATCGCGAGTGTGCAG; U5, GAACAAGCA CGTGAGAAGGCAGG; U7, GCGTCTGTAAGGCAGATTGTG; U8, TAAG ATAGGTTTAGTAAGACG; 1R1, TATACACCTGGAGTAGGAAC; 1R2, AATAACTACTCTTCCTTCC; 1R4, CATACTGTACTGCACTCATCC; 1R7, TGCATCGTCATTACGCACAAGGTC; 4F2, TGCTCCGGATTCTACCAA AG; 4F3, AAGGAACTAACAAAAGCTCC; D1, TATTGATCAAAGTAC CTCAGCG; and D2, GCCTAAATACTCAGAAGCGCG.
DNA sequencing and data analysis.DNA sequencing reactions used
fluores-cently labeled dideoxynucleotide technology (dye terminator cycle sequencing ready reaction kit; Perkin-Elmer). Sequencing reaction products were separated, and data were collected using an ABI 377 automated DNA sequencer (Perkin-Elmer). The sequence was fully determined for both strands of each DNA template to ensure maximum accuracy of the data. Sequences were edited and assembled using the Staden software programs (6) and analyzed using the Wis-consin Sequence Analysis Package (Genetics Computer Group, Madison, Wis.) (8).
Sequences were aligned using the Pileup program of the GCG package (8). Phylogenetic analysis was performed with the PAUP program (version 4.0.0d64) on a Power Macintosh 9500/132. The maximum parsimony optimality criterion was used for a heuristic search, and the resulting unrooted tree was the product of 100 bootstrap replicates.
Nucleotide sequence accession numbers.GenBank accession numbers for the
ankgene sequences are as follows: EE (E. equi), AF100882; NY1, AF100883; NY2, AF100884; NY3, AF100885; Sl-HG1, AF100886; Sl-HG2, AF100887; Sw-HG, AF100888; EP (E. phagocytophila), AF100889; WI1, AF100890; WI2, AF100891; WI3, AF100892; WI4, AF100893; and MN-dog, AF100894.
RESULTS
PCR amplification.The completeankgene, including 28 bp
upstream of the start codon and 16 bp downstream from the stop codon, was amplified by using a combination of multiple primers and a nested PCR strategy. PCR primers were initially designed based on theankgene sequence of the USG3 strain (24). Additional sequencing of theankgene from this strain revealed that the open reading frame consists of 3,696 and not 2,244 bp as originally reported (24). Theankgene was divided into seven overlapping regions of 550 to 600 bp, and primer sets were designed to specifically amplify each of these regions. The locations of primers used for PCR and sequencing are shown in Fig. 1. A nested PCR strategy was used to amplify the ankgene for two reasons. First, the initial direct PCR
ampli-fication attempts resulted in little or no product with many of the templates (data not shown) because many of the clinical samples used in this study contained a low concentration of the ehrlichial agents. Second, several samples were limited by the volume of the sample. Therefore, a nested protocol that al-lowed both increased sensitivity and conservation of limited amounts of samples was developed. The primary reactions used primers U7 and 1R1 to amplify the 5⬘portion of the gene and primers 1F and 4R1 for the 3⬘ region. The products of these reactions were used as templates for each of the specific nested or heminested reactions for the seven regions. The downstream regions were subsequently amplified indepen-dently by using primary reactions with 4F2 and D2 followed by nested reactions using 4F3 and D1.
DNA sequence analysis. The samples that were used for
[image:2.612.316.550.70.192.2]PCR amplification and sequencing of theankgene are shown in Table 1. The total number of base pairs sequenced from each specimen and the predicted number of amino acids en-coded within the gene are also shown in Table 1. The 28 bp FIG. 1. Primers used for amplification of theankgene and topography of the deduced Ank protein. The rectangle represents the coding region in 5⬘-to-3⬘ orientation. The length indicated for the open reading frame (3,696 bp) is for the USG3 strain. The location and orientation of the primers used for PCR ampli-fication and DNA sequencing are shown relative to theankgene coding region. The products of each primer pair are shown as dashed lines. Superimposed on the coding region are the ankyrin-repeat region and the region of the coding sequence containing the three types of repetitive elements in the deduced Ank protein for the USG3 strain of the HGE agent.
TABLE 1. Sample identifiers, sources, collection locations, andankgene DNA and protein sequence data
Name Source Location ORF
bsize
bp aac
USG3 I. scapularis NY/PAa 3,696 1,232
NY1 Human Westchester County, N.Y. 3,696 1,232 NY2 Human Westchester County, N.Y. 3,696 1,232 NY3 Human Westchester County, N.Y. 3,696 1,232 WI1 Human Trempealeau County, Wis. 3,693 1,231 WI2 Human Barron County, Wis. 3,693 1,231 WI3 Human Clark County, Wis. 3,693 1,231 WI4 Human Marathon County, Wis. 3,693 1,231 MN-dog Canine Hennepin County, Minn. 3,693 1,231 EE Equine Northern California 3,615 1,205
EP Bovine Sweden 3,615 1,205
Sw-HG Human Sweden 3,618 1,206
Sl-HG1 Human Slovenia 3,669 1,223
Sl-HG2 Human Slovenia 3,720 1,240
aIsolate from ticks collected in Westchester County, N.Y., and Montgomery
County, Pa.
bORF, open reading frame. caa, amino acids.
on May 15, 2020 by guest
http://jcm.asm.org/
[image:2.612.311.552.534.694.2]upstream of the predicted initiating codon and 16 bp down-stream of the stop codon were identical in length for each sample used in this study. Each of the three human samples from New York showed anankcoding region equal in length to that of the USG3 reference strain (3,696 bp), and the nu-cleotide sequences for two of the New York samples (NY1 and NY2) were identical to the nucleotide sequence of the USG3 strain. Likewise, each of the samples from the Upper Midwest (WI1, WI2, WI3, WI4, and MN-dog) showed sequences of equal size (ankcoding region of 3,693 bp), and theE. equiand E. phagocytophilasequences were of equal size (3,615 bp). In contrast, each of the three European HGE sequences showed unique sizes: 3,618 bp for the Swedish HGE agent and 3,669 and 3,720 bp for the Slovenian samples. A feature unique to the Northeastern U.S. samples (New York and USG3) is a 3-bp insertion (TTT) at nucleotide position 2470 that is absent in all other samples examined in this study. This single codon insertion is also responsible for the 3-bp difference in size between the New York (3,696-bp) and the Upper Midwest (3,693-bp) samples.
The nucleotide sequences were aligned, and the calculated percent identities are shown in Table 2. The nucleotide ho-mology among the North American HGE samples was very high, with a range of 98.16 to 100%. The sequences for four of the five samples from the Upper Midwest (WI2, WI3, WI4, and MN-dog) were identical. The remaining Upper Midwest sam-ple (human WI1) showed 99.95% identity to each of the other Midwestern sample sequences because of a 2-bp difference. Regarding the samples from the Northeastern United States, the NY1, NY2, and USG3 samples were identical and differed from the NY3 sample by a single nucleotide (99.97% identity). Comparing the North American HGE sequences to the Euro-pean HGE sequences showed a range of 95.54% (for NY3 and Sl-HG2) to 97.22% (WI2 and Sw-HG) identity. The E. equi sequence showed highest homology to the sequences of the Wisconsin and Minnesota samples (99.18 to 99.24%), less ho-mology to the New York samples (97.90 to 97.92%), and least homology to the European HGE andE. phagocytophila sam-ples (96.70 to 97.46%). Conversely, the E. phagocytophila quence was most homologous to the European HGE se-quences (97.76 to 97.79%) and showed less homology to the sequences from the North American samples (95.90 to 96.56%).
Protein sequence analysis. The predicted amino acid
se-quences were also aligned, and the percent identities are shown in Table 2. Similar to the nucleotide sequence results, the amino acid sequences clearly could be classified as
belong-ing in one of two groups, either North American or European, and the North American group could be further subdivided into either Upper Midwest or Northeastern samples. Interest-ingly, the percent identities between the groups were lower for the amino acid sequences than for the nucleotide sequences, indicating that many of the nucleotide changes represent non-silent mutations that result in protein sequence variations. Amino acid homology between the Upper Midwest and North-eastern samples ranged from 96.34 to 96.43%. In contrast, the amino acid sequences of each of the four samples from the Northeast were identical, and only a single amino acid differ-ence was noted among the Upper Midwest samples (99.92% identity). The homology within the European HGE group was also very high and ranged from 99.92 to 100%. The SwedishE. phagocytophilasequence (EP) showed lower homology, with a range from 92.61 to 93.61% identity to members of the North American group and from 95.44 to 95.52% identity to the European HGE group. The highest percent identity noted between members of the European and North American groups was 95.27% (Sw-HG and WI2).
[image:3.612.51.553.83.200.2]The nucleotide and amino acid sequence analyses described above and shown in Table 2 suggest the existence of three distinct groups of the HGE agent (Northeast United States, Upper Midwest United States, and European), with a high degree of homology within each group and lesser homology between groups. The homologies among these three groups of the HGE agent were examined further by dividing the Ank protein into two regions: an 850-residue amino-terminal region (849 residues for Upper Midwest samples) that includes the ankyrin repeat elements and a carboxyl-terminal region rang-ing from 356 residues (Sw-HG) to 390 residues (Sl-HG2) that contains the 27-, 17-, and 11-residue repeats. In comparison of only the residues that were conserved among each member of a given group, the protein coding changes were relatively evenly distributed between the amino-terminal and carboxyl-terminal regions in comparing the European group to either of the North American groups. The corresponding N-terminal region comparison showed identities ranging from 92% (Eu-ropean to Northeastern United States) to 94.7% (Eu(Eu-ropean to Midwestern United States), while the C-terminal region ho-mology was 94.3% (European to either Northeastern or Mid-western United States). In contrast, the same comparison be-tween the two North American groups showed that the N-terminal regions of the proteins were considerably more variable (93.8% identity) than the C-terminal regions of the proteins (99.7% identity).
TABLE 2. ankgene nucleotide sequence and deduced amino acid residue homologya
Sample % Homology for nucleotide (roman) and deduced amino acid (italic) sequence
USG3b NY3 WI1 WI2c Sw-HG Sl-HG1 Sl-HG2 EE EP
USG3b 99.97 98.18 98.24 96.45 95.80 95.57 97.92 95.93
NY3 100 98.16 98.21 96.43 95.78 95.54 97.90 95.90
WI1 96.34 96.34 99.95 97.16 96.50 96.34 99.18 96.50
WI2c 96.43 96.43 99.92 97.22 96.56 96.39 99.24 96.56
Sw-HG 93.95 93.95 95.19 95.27 99.97 99.97 97.46 97.79
Sl-HG1 92.97 92.97 94.19 94.28 99.92 99.95 97.43 97.76
Sl-HG2 93.02 93.02 94.31 94.39 100 99.92 97.43 97.76
EE 95.93 95.93 98.42 98.51 94.11 94.52 95.02 96.70
EP 92.61 92.61 93.53 93.61 95.52 95.44 95.52 92.37
aSequences were aligned using the GCG Pileup program, and percent identity was determined by the Olddistances program using length of shorter sequence without
gaps as the denominator.
bUSG3 also represents NY1 and NY2. cWI2 also represents WI3, WI4, and MN-dog.
on May 15, 2020 by guest
http://jcm.asm.org/
Repetitive elements.The sequence of theankgene for the USG3 strain showed two copies of an 81-bp (27-amino-acid) repeat beginning at nucleotide position 2828 (23). This repeat was also found in theankgene sequence for each of the North American HGE agent samples and the Minnesota canine sam-ple. However, only a single copy of the 81-bp sequence was found in theE. equisequence, resulting in an 81-bp deletion in the E. equi ank gene relative to the other North American samples that were tested. Similarly, a single copy of the 81-bp element was present in each of the European samples. The Slovenianankgene sequences also showed a variable number of copies of a 51-bp (17-amino-acid) repetitive element located directly adjacent to and downstream from the 81-bp element. Whereas each of the North American, Swedish HGE (Sw-HG), andE. phagocytophila(EP) samples showed a single copy of this 51-bp sequence, the Slovenian HGE samples (Sl-HG1 and Sl-HG2) showed two and three copies of the element, respectively. Two 11-amino-acid repeats were conserved in all samples examined. The arrangement of these repetitive ele-ments (represented by the number of amino acids) within the ankgene for each sample examined is shown in Fig. 2.
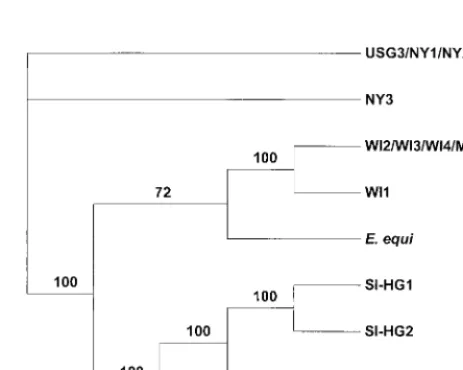
Phylogenetic analysis.Theankgene sequences were used as
a phylogenetic tool to assess the relationship of the GE that were examined in this study. Samples with sequences that were identical were removed from the analysis but are shown in the phylogram in Fig. 3. These results separate the North Ameri-can samples into two clades, one including the New York HGE and USG3 samples and another with the Wisconsin HGE and Minnesota dog samples. Likewise, the phylogram effectively positions the four European samples in a separate clade. The position of theE. equisequence in the Upper Midwest clade is supported by 72 bootstrap replicates.
DISCUSSION
Members of the GE genogroup have been shown to cause disease in numerous vertebrate species including dogs, cattle, sheep, horses, and humans. To date, analysis of the members of this group at the molecular level has consisted primarily of examination of the DNA sequence of the 16S rRNA gene. These analyses have shown very little difference within the 16S rRNA sequences, suggesting that the HGE agent,E. equi, and E. phagocytophila represent strains of a single species. Addi-tionally, the 16S rRNA sequences determined for the HGE agent from confirmed human cases from both North America (Upper Midwest and Northeast) and Europe, including those used in this study, have been identical, suggesting that the agent causing human disease in these areas may represent a
single strain. Recently, two human cases were reported from northern California that showed 16S rRNA sequences differing from the HGE agent by 1 bp and identical to the E. equi sequence, suggesting that anE. equi-like strain may be causing infections in northern California (10). In contrast to the 16S rRNA data, studies using DNA sequence and Western blot analyses of the major antigenic proteins have suggested a high degree of variability among HGE isolates from both the North-eastern and Upper Midwestern United States (13, 32, 33). Although analysis of the 16S rRNA gene sequence is a pow-erful tool for identifying novel agents and for determining relatedness at the genus and species level, the strong conser-vation of the 16S rRNA makes it less than ideal for differen-tiation of closely related species or subspecies. Sequences of genetic elements more variable than the highly conserved ri-bosomal subunits and housekeeping genes, such as genes en-coding structural proteins, are often needed to make these determinations. Recently, the sequences of several genes en-coded by the GE strain USG3 were determined, one of these being theankgene. Analysis of the predicted peptide sequence suggested that theankgene may encode a structural protein, and in this study the complete sequence of theankgene was determined for 13 members of the E. phagocytophila geno-group including samples from 10 confirmed HGE cases.
[image:4.612.54.293.72.188.2]Phylogenetic analysis of theankgene sequences separated the HGE agent into three distinct clades representing the U.S. Northeast and Upper Midwest and Europe. Analysis of the ankgene sequence thereby represents the first epidemiologic tool able to differentiate the HGE agent based on geographic origin of the sample and the first to indicate that the etiologic agents of HGE present in each of these locations are evolving independently. The sequence conservation noted for the HGE agent sequences within each of the three clades was remark-ably high, with both nucleotide and amino acid identities of ⬎99.9%. This was in contrast to nucleotide and amino acid sequence homologies among members of the three clades that were significantly lower (nucleotide sequence identity of ⬍98.2%; amino acid identity of⬍96.5%). Numerous identical sequences were noted for members within the same clade, including three of the four samples from the U.S. Northeast.
FIG. 2. Variable number of repetitive elements within theankgene. The number of 81-bp (27-amino-acid) and 51-bp (17-residue) repeats and the ar-rangement of the repeats are shown for the GE examined. N.A., North Amer-ican;E. phag.,E. phagocytophila.
FIG. 3. Phylogenetic analysis of the GE based on theankgene DNA se-quences using maximum parsimony as the optimality criterion. The number of bootstrap replicates (from a total of 100) that were in agreement are shown on each branch. Branches without bootstrap values represent polytomies that could not be resolved because the sequences were too similar.
on May 15, 2020 by guest
http://jcm.asm.org/
[image:4.612.316.548.498.683.2]Likewise, three of the four HGE agent samples from the Up-per Midwest were identical. The samples from the Northeast were also collected in three different years, 1993 (USG3), 1994 (NY2), and 1996 (NY1 and NY3), suggesting that there is minimal year-to-year variation within this population. Further-more, PCR amplification and DNA sequencing of a 500-bp region of theankgene from additional samples collected from confirmed HGE cases in Minnesota, Wisconsin, and Connect-icut support the results described herein and the conclusion that two distinct clonal populations of the HGE agent exist in the United States, defined by geographic location, Northeast and Upper Midwest (data not shown).
The USG3 strain was isolated from a canine infected by allowing Ixodes scapularis ticks collected from Westchester County, N.Y., and Montgomery County, Pa., to feed. The isolation of this strain has raised questions concerning the authenticity of the USG3 strain as a representative of the HGE agent and the efficacy of using this strain as an antigen for serodiagnostic purposes (19, 31). However, the sequence de-termined for theankgene of the USG3 strain was identical to that of two of the samples from confirmed HGE cases in New York, suggesting that the USG3 strain likely represents a strain of the HGE agent. Similarly, the sequence determined for the Minnesota dog sample was identical to three of the four Wis-consin HGE samples, suggesting that a single strain, or a very closely related one, is capable of infecting and causing clinical illness in both human and canine species. In addition, portions of at least six genes other than the 16S rRNA,groE, andp44 have been sequenced from the USG3 strain and were identical to those from a New York human isolate (C. I. Murphy et al., unpublished observations).
Theankgene sequences were determined and analyzed for single representatives of E. equi and E. phagocytophila and were found to be no more closely related to each other than to the North American and European HGE agent sequences. In fact, the lowest homology found between any two amino acid sequences (92.37%) was between the predictedE. equiandE. phagocytophila proteins. While theE. phagocytophila nucleo-tide and amino acid sequences were clearly more closely re-lated to the European HGE agent sequences than to those of any of the North American samples, the E. equi sequences showed highest homology to those from the Upper Midwest samples. Additional representatives of both of these species need to be examined before drawing any conclusions, particu-larly forE. phagocytophila, where previous studies of thegroE heat shock operon sequences showed significant variability be-tween E. phagocytophilastrains relative to the high conserva-tion noted for the 16S rRNA gene (25). However, a partial sequence of theankgene from a SwissE. phagocytophila sam-ple showed ⬎99% identity to the Swedish E. phagocytophila examined in this study and suggests a close relationship be-tween European strains ofE. phagocytophila(data not shown). One of the more distinctive features within the ank gene sequence is a region containing a variable number of direct repeats with no ankyrin homology located in the last third of the gene. The result of these repeats at the protein level is a 27-amino-acid sequence that is repeated twice in each of the North American samples, with the exception of E. equi. The European samples andE. equishow a single copy of the 27-amino-acid element, suggesting that this organism may be more closely related to European GE than are the other North American GE. In fact, the number and arrangement of repet-itive elements forE. equiare identical to those noted for both the Swedish HGE agent andE. phagocytophila. However, the partial sequence of theankgene for the BDS strain ofE. equi (GenBank accession no. AF047897) shows a repeat structure
identical to that of the North American HGE agents and suggests that the number of copies of the 27-amino-acid repeat in E. equi strains can be variable. Directly adjacent to and downstream from the 27-amino-acid unit is a 17-amino-acid element that is present as a single copy in each of the North American HGE samples,E. equi, the Swedish HGE agent, and E. phagocytophila. Only the European HGE samples from Slo-venia show multiple copies of the 17-amino-acid element, with either two or three copies present, and thereby appear to represent a feature unique to the HGE agent in Slovenia.
A comparison of the members of the three clades of the HGE agent (U.S. Northeast and Upper Midwest and Europe) showed that the degree of homology is not uniform across the length of theankgene or the predicted protein. Analysis of the amino-terminal region of the protein sequence for the mem-bers of each clade showed a similar degree of identity among the three clades (92.0 to 94.7%). In contrast, the carboxyl-terminal region of the protein is very highly conserved between the members of the two North American clades (⬎99.7% iden-tity), while the European clade members show only 94.3% identity to each of the North American clades. These data suggest that there are constraints on the evolution of the car-boxyl-terminal portion of the protein in the North American strains that are not being exerted on the more variable amino-terminal region. It is the amino-amino-terminal region of the protein that contains the ankyrin-like repeats from which the name of the gene is derived. Ankyrin-related proteins have been de-scribed for bacteria, plants, and animals, and the most common function attributed to ankyrin-like repeats involves protein-protein interactions (3, 16). The diversity within the amino-terminal region of the HGE agent Ank protein indicates that, although the length of the amino-terminal region is conserved, the content is flexible. The fact that there are between 14 and 18 copies of ankyrin-like repeats may allow for minor varia-tions within the repeats to have little or no effect on the overall function of the protein, thereby allowing more diversity within this region of the protein.
The life cycle of the HGE agent involves a complex inter-action between the natural host(s) or reservoirs and vectors that progress through multiple life stages and may transmit infections to humans. Additionally, tick species that rarely bite humans, including nidicolous members of the genus Ixodes, such as Ixodes trianguliceps in Europe, are likely involved as vectors for maintaining enzootic infectious cycles (20). This requires that the agent have the ability to adapt to these mul-tiple environments and environmental pressures. The differ-ences noted for the Ank protein among members of the three clades may reflect adaptation to differences between the vec-tors and/or reservoirs of the agent. While the presumed vector, I. scapularisticks, and the major reservoir,Peromyscus leuco-pus, are present in both the Upper Midwestern and Northeast-ern regions of the United States, the genetic and biologic diversity between these populations living in distinct geo-graphic locations has not been fully explored. There are also potentially significant differences between the vectors and res-ervoirs that are involved in the life cycles of the agents in North America and Europe. Whereas I. scapularis is the primary vector in the United States, a different species of Ixodid tick, Ixodes ricinus, has been suggested as the vector in Europe (7, 15, 18, 20–23, 28). WhereasP. leucopushas been shown to be a competent natural reservoir in the United States (27, 30), the corresponding reservoir in Europe has not been defined al-though Ogden et al. (20) identified GE closely related to the HGE agent inApodemus sylvaticuswood mice and Clethriono-mys glareolusbank voles in the United Kingdom, and Liz et al. (15) found the highest prevalence of GE in C. glareolus in
on May 15, 2020 by guest
http://jcm.asm.org/
Switzerland. Resolution of the mechanisms that are driving the evolution of theankgene will require additional studies ad-dressing the expression of the gene and characterization of the functional properties of the encoded protein, as well as a better understanding of the vectors and reservoirs of the HGE agent in the United States and Europe. Whereas the present study focused on the examination of theank gene amplified from confirmed human cases and suggests that this gene provides an excellent epidemiologic tool for differentiating HGE agent iso-lates, the diversity noted within the gene should prove useful for examining the heterogeneity of GE in veterinary and ar-thropod populations.
ACKNOWLEDGMENTS
We are grateful to the Biotechnology Core Facility of the National Center for Infectious Diseases for the synthesis of oligonucleotides and to Dana Jones for assistance with the phylogenetic analysis.
REFERENCES
1.Anderson, B. E., J. E. Dawson, D. C. Jones, and K. H. Wilson.1991.Ehrlichia
chaffeensis, a new species associated with human ehrlichiosis. J. Clin. Micro-biol.29:2838–2842.
2.Bakken, J. S., J. S. Dumler, S.-M. Chen, M. R. Eckman, L. L. Van Etta, and
D. H. Walker.1994. Human granulocytic ehrlichiosis in Upper Midwest
United States. A new species emerging? JAMA272:212–218.
3.Bork, P.1993. Hundreds of ankyrin-like repeats in functionally diverse
pro-teins: mobile modules that cross phyla horizontally. Proteins Struct. Funct. Genet.17:363–374.
4.Broqui, P., J. S. Dumler, R. Lienhard, M. Brossard, and D. Raoult.1995.
Human granulocytic ehrlichiosis in Europe. Lancet346:782–783.
5.Chen, S.-M., J. S. Dumler, J. S. Bakken, and D. H. Walker.1994.
Identifi-cation of a granulocytotropicEhrlichia species as the etiologic agent of human disease. J. Clin. Microbiol.32:589–595.
6.Dear, S., and R. Staden.1991. A sequence assembly and editing program for
efficient management of large projects. Nucleic Acids Res.19:3907–3911.
7.Des Vignes, F., and D. Fish.1997. Transmission of the agent of human
granulocytic ehrlichiosis by host-seekingIxodes scapularis(Acari: Ixodidae) in southern New York state. J. Med. Entomol.34:379–382.
8.Devereux, J., P. Haeberli, and O. Smithies.1984. A comprehensive set of
sequence analysis programs for the VAX. Nucleic Acids Res.12:387–395.
9.Dumler, J. S., and J. S. Bakken.1995. Ehrlichial diseases of humans:
emerg-ing tick-borne infections. Clin. Infect. Dis.20:1102–1110.
10. Foley, J. E., L. Crawford-Miksza, J. S. Dumler, C. Glaser, J.-S. Chae, E. Yeh,
D. Schnurr, R. Hood, W. Hunter, and J. E. Madigan.1999. Human
granu-locytic ehrlichiosis in Northern California: two case descriptions with genetic analysis of the ehrlichiae. Clin. Infect. Dis.29:388–392.
11. Gewirtz, A. S., P. J. Cornbleet, D. J. Vugia, C. Traver, J. Niederhuber, C. P.
Kolbert, and D. H. Persing.1996. Human granulocytic ehrlichiosis: report of
a case in northern California. Clin. Infect. Dis.23:653–654.
12. Hardalo, C. J., V. Quagliarello, and J. S. Dumler.1995. Human granulocytic
ehrlichiosis in Connecticut: report of a fatal case. Clin. Infect. Dis.21:910– 914.
13. Kim, H.-Y., and Y. Rikihisa.1998. Characterization of monoclonal
antibod-ies to the 44-kilodalton major outer membrane protein of the human gran-ulocytic ehrlichiosis agent. J. Clin. Microbiol.36:3278–3284.
14. Kolbert, C. P., E. S. Bruinsma, A. S. Abdulkarim, E. K. Hofmeister, R. B.
Tompkins, S. R. Telford III, P. D. Mitchell, J. Adams-Stich, and D. H.
Persing.1997. Characterization of an immunoreactive protein from the
agent of human granulocytic ehrlichiosis. J. Clin. Microbiol.35:1172–1178.
15. Liz, J. S., L. Anderes, J. W. Sumner, R. F. Massung, L. Gern, B. Rutti, and
M. Brossard.2000. PCR detection of granulocytic ehrlichiae inIxodes ricinus
ticks and wild small mammals in western Switzerland. J. Clin. Microbiol.
38:1002–1007.
16. Lux, S. E., K. M. John, and V. Bennett.1990. Analysis of cDNA for human
erythrocyte ankyrin indicates a repeated structure with homology to tissue-differentiation and cell-cycle control proteins. Nature (London)344:36–42.
17. Maeda, K., N. Markowitz, R. C. Hawley, M. Ristic, D. Cox, and J. E.
McDade.1987. Human infection withEhrlichia canis, a leukocytic rickettsia.
N. Engl. J. Med.316:853–856.
18. Magnarelli, L. A., K. C. Stafford III, T. N. Mather, M.-T. Yeh, K. D. Horn,
and J. S. Dumler.1995. Hemocytic rickettsia-like organisms in ticks:
sero-logic reactivity with antisera to ehrlichiae and detection of DNA of agent of human granulocytic ehrlichiosis by PCR. J. Clin. Microbiol.33:2710–2714.
19. Nicholson, W. L., J. A. Comer, J. W. Sumner, C. Gingrich-Baker, R. T.
Coughlin, L. A. Magnarelli, J. G. Olson, and J. E. Childs.1997. An indirect
immunofluorescence assay using a cell culture-derived antigen for detection of antibodies to the agent of human granulocytic ehrlichiosis. J. Clin. Mi-crobiol.35:1510–1516.
20. Ogden, N. H., K. Brown, B. K. Horrocks, Z. Woldehiwet, and M. Bennett.
1998. Granulocytic Ehrlichiaeinfection in Ixodid ticks and mammals in woodlands and uplands of the U.K. Med. Vet. Entomol.12:423–429.
21. Pancholi, P., C. P. Kolbert, P. D. Mitchell, K. D. Reed, J. S. Dumler, J. S.
Bakken, S. R. Telford, and D. H. Persing.1995.Ixodes damminias a
poten-tial vector of human granulocytic ehrlichiosis. J. Infect. Dis.172:1007–1012.
22. Petrovec, M., S. L. Furlan, T. A. Zupanc, F. Strle, P. Broqui, V. Roux, and
J. S. Dumler.1997. Human disease in Europe caused by a granulocytic
Ehrlichiaspecies. J. Clin. Microbiol.35:1556–1559.
23. Pusterla, N., J. B. Huder, C. M. Leutenegger, U. Braun, J. E. Madigan, and
H. Lutz.1999. Quantitative real-time PCR for detection of members of the
Ehrlichia phagocytophilagenogroup in host animals andIxodes ricinusticks. J. Clin. Microbiol.37:1329–1331.
24. Storey, J. R., L. A. Doros-Richert, C. Gingrich-Baker, K. Munroe, T. N.
Mather, R. T. Coughlin, G. A. Beltz, and C. I. Murphy.1998. Molecular
cloning and sequencing of three granulocyticEhrlichiagenes encoding high-molecular-weight immunoreactive proteins. Infect. Immun.66:1356–1363.
25. Sumner, J. W., W. L. Nicholson, and R. F. Massung.1997. PCR
amplifica-tion and comparison of nucleotide sequences from thegroESLheat shock operon ofEhrlichiaspecies. J. Clin. Microbiol.35:2087–2092.
26. Sumption, K. J., D. J. M. Wright, S. J. Cutler, and B. A. S. Dale.1995.
Human ehrlichiosis in the UK. Lancet346:1487–1488.
27. Telford, S. R., III, J. E. Dawson, P. Katavolos, C. K. Warner, C. P. Kolbert,
and D. H. Persing.1996. Perpetuation of the agent of human granulocytic
ehrlichiosis in a deer tick-rodent cycle. Proc. Natl. Acad. Sci. USA93:6209– 6214.
28. von Stedingk, L. V., M. Gu¨rtelschmid, H. S. Hanson, R. Gustafson, L.
Dotevall, E. O. Engvall, and M. Granstro¨m.1997. The human granulocytic
ehrlichiosis (HGE) agent in Swedish ticks. Clin. Microbiol. Infect.3:573–574.
29. Walker, D. H., and J. S. Dumler.1996. Emergence of human ehrlichioses as
human health problems. Emerg. Infect. Dis.2:18–29.
30. Walls, J. J., B. Greig, D. F. Neitzel, and J. S. Dumler.1997. Natural infection
of small mammal species in Minnesota with the agent of human granulocytic ehrlichiosis. J. Clin. Microbiol.35:853–855.
31. Yeh, M.-T., T. N. Mather, R. T. Coughlin, C. Gingrich-Baker, J. W. Sumner,
and R. F. Massung.1997. Serologic and molecular detection of granulocytic
ehrlichiosis in Rhode Island. J. Clin. Microbiol.35:944–947.
32. Zhi, N., N. Ohashi, and Y. Rikihisa.1999. Multiplep44genes encoding
major outer membrane proteins are expressed in the human granulocytic ehrlichiosis agent. J. Biol. Chem.274:17828–17836.
33. Zhi, N., Y. Rikihisa, H. Y. Kim, G. P. Wormser, and H. W. Horowitz.1997.
Comparison of major antigenic proteins of six strains of the human granu-locytic ehrlichiosis agent by Western immunoblot analysis. J. Clin. Microbiol.
35:2606–2611.