a result of the importance of the visual system in an animal's survival, the majority of previous studies have investigated how animals associate visual cues with spatial locations. Because the sexual behavior of the túngara frog (Physalaemus pustulosus) relies heavily on the use of acoustic signals, I hypothesized that they would be able to use acoustic cues to learn spatial locations. To test this hypothesis, I presented each individual with two arbitrary sounds, a tone and a recording of a mating call that had been reversed. The tone was initially associated with the correct door of the maze, which led to a shelter. Once an individual learned the association, I switched the sounds so that the reversed mating call was now associated with the correct door. My results indicate that túngara frogs are able to associate an arbitrary sound with a location and to reverse those associations. This study is an important contribution to research on anuran cognition because it is the first to demonstrate the use of acoustic signals during spatial learning, as opposed to the predominant visual studies.
INTRODUCTION
Learning is critical to an animal’s effective navigation of its environment, procurement of resources, parental care, predator avoidance, and ultimate survival. As a result, animals have evolved effective sensory systems that enable them to learn and navigate a rapidly changing environment, including visual, auditory, olfactory, tactile, and magnetic senses (Harrison, 1984). The importance of learning studies has long been appreciated, ranging from the common “rat in a maze” experiment, to the many complex learning experiments that are being explored today.
1985, Giurfa, 2004, Kelber, 2002, Brattstrom, 1990). Recently, studies have emerged focusing on other modes of learning. Rats have demonstrated an ability to use olfactory cues to learn a task (Devore, 2013), while pigs (and other mammals) have exhibited similar abilities using acoustic cues (Murphy, 2013).
Anuran learning studies were originally limited because researchers believed that these animals were behaviorally inflexible (Thorpe, 1963). However, further research demonstrated that it was necessary to use biologically relevant methodologies in order to produce effective motivation in individuals (Ellins, 1982). Today, limited studies exist on maze learning in amphibians, but many continue to emerge. Maze learning has been demonstrated in toads, including tasks in which both visual and auditory cues are associated with a reward (either escape, shelter, or food) (Schmajuk, 1980, Brattstrom, 1990). However, it has yet to be shown that amphibians can specifically associate a sound with a particular reward to learn a task. In this experiment, the acoustic learning abilities of the túngara frog, a well-studied model for acoustic communication, were explored.
associate an arbitrary cue with a reward, demonstrating higher learning capabilities than exhibited with the use of a visual cue.
In this experiment, we used a two-arm maze to assess an individuals’ ability to associate one of two sounds with the maze exit. We found that that the túngara frog can effectively associate one of two auditory stimuli with a reward, as well as the reversal of this task. The reversal of this particular task involved the frogs learning a new stimulus reward-pairing, in which the other, previously irrelevant, auditory stimulus was now associated with the reward. These results suggest that anurans, specifically the túngara frog, can exhibit learning capabilities that are equivalent to that of other animals. In addition, our results suggest that learning
capabilities in many animals are highly dependent on the behavioral modalities that are most applicable to their natural habitat and behavior.
MATERIALS AND METHODS Animals
We used six individuals in the experiment, four females and two males. We kept the animals in a temperature controlled room (27 ̊ C). We spritzed the animals with water every other day and fed them wingless fruit flies three times a week, along with calcium and multiple vitamin powder. We set their light cycle to approximately 7 AM – 7 PM, similar to where they originate in the tropical rainforest of Central America.
Maze Set-up
addition, we placed two 150 watt heating lamps directly above the maze, resulting in a temperature of 37-40̊C throughout the maze. We placed a camera between the two heating lamps to record individuals’ movements.
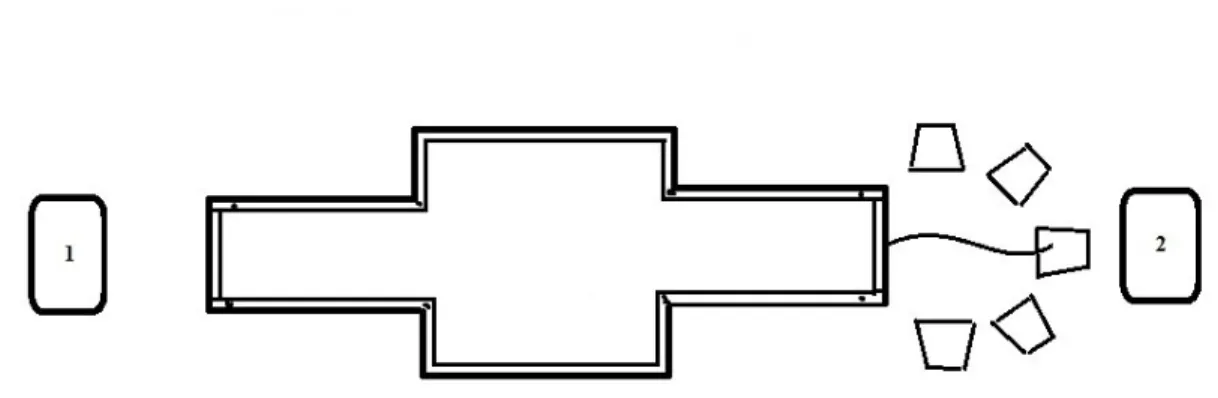
We surrounded the maze by white sheets on all four sides in order to eliminate additional visual cues. Between the maze and the sheet of Plexiglas we placed a piece of absorbent paper, which we replaced daily in order to eliminate olfactory cues. We placed five shelters outside of each door of the maze that were insulated and therefore cooler than the rest of the maze
environment. Behind these shelters on both sides we placed the speaker that played an auditory cue.
Figure 1: Maze Set-up. Sounds
auditory range of the túngara frog. In the literature, this species has no natural attraction to either sound (Wilczynski, 1995). In addition, we conducted a pilot study to confirm that individuals had no preference for either sound over the other. In this pilot study, we placed each individual in the maze with both doors open and the sounds alternating playing, identical to how the sounds would be played during the trials. We recorded the door used for escape for each individual over three days of pilot trials, and determined that the individuals did not have an inherent preference for either sound.
Procedure
Acquisition
In the acquisition period, we expected the frogs to associate the correct door, or escape, with the tone, not the reversed whine. We ran two trials per day, beginning between 8:30 and 9:30 AM. We removed both the male and female cages from the room and misted the
individuals. We misted the individuals with water so that the maze felt very hot and dry, thereby increasing the individual’s motivation to escape. At the beginning of each day, before the
In each trial, we captured the individual using a clay cup and a square sheet of paper to minimize human contact. We placed this cup face down in the center of the maze, with both maze doors closed. The cup remained over the frog for 15 seconds while the two sounds played, antiphonally (non-overlapping). After 15 seconds, we removed the cup and the frog was allowed to move freely within the maze. We started the timer upon the frog’s first movement. We stopped the timer either when the frog passed within approximately two centimeters from the correct door, or upon the conclusion of three minutes. If the individual did not reach the correct door in the span of three minutes, we stopped the timer and opened the correct door. Then, we gave the individual as much time as needed to exit the maze and enter a shelter. Upon entering the shelter, we gave the frog 45 seconds to reinforce the reward before being placed back in the cage.
We considered the trial a success if the frog crossed approximately two centimeters (marked in Figure 1) in front of the correct door before the termination of three minutes. Once the frog cross this two centimeter mark, we opened the door to allow the frog to exit. If the individual crossed approximately two centimeters in front of the incorrect door it was given an error. It is possible for individuals to achieve both an error and success in one trial. In addition, we recorded the latency (amount of time spent in the maze, maximum 180 seconds) for each individual in each trial. The criterion for learning the acquisition is that of five successes without error in six consecutive trials.
Reversal
is the same as the criterion for acquisition. Only three individuals were run in the reversal trials because two individuals died after the acquisition.
Data Analysis
For the acquisition period, we determined the success rate of each individual by averaging the number of successes each day (across the two trials). Once these success rates were calculated and recorded (ranging from 0-1 for each day), we calculated the average success rate across individuals for each day (again ranging from 0-1). I ran a repeated measures ANOVA for the first 11 days of acquisition and the first 14 days of reversal. I only analyzed the first 11 and 14 days because the first individual reached criterion on days 11 and 14 for the acquisition and reversal, respectively – therefore, the program was unable to analyze the group with empty data cells for individuals that had already reached criterion.
I graphed the latency data in a similar fashion. I averaged the latency for each individual across the two trials for each day (maximum 180 seconds). After compiling this data for each individual, I averaged the daily average latency across all individuals for each day. I graphed this data against each day for both the acquisition and reversal (Figures 4 and 5). I ran a repeated measures ANOVA for the first 11 days of acquisition latency data, and the first 14 days of reversal latency data.
RESULTS
success rate for the reversal gradually increased from Day 1 to Day 17 (Figure 3). This also demonstrates that the individuals approached the criterion for reversal.
The increase in success rate across the first 11 days of acquisition demonstrates that the individuals were becoming more and more successful in their trials in the maze, reached the criterion, and therefore learned the task (P-value: 0.042). The increase in success rate across the first 14 days of reversal demonstrates that the individuals were able to reverse their stimulus-reward association over time, reached the criterion, and therefore successfully reversed the task (P-value 0.048).
The latency of each individual gradually decreases over the 17 days of acquisition (Figure 4). This decrease in latency demonstrates that the individuals learned the task, as they were spending less and less time completing the maze and becoming more adept at the task (P-value: 0.040). The latency of each individual additionally decreases over the 17 days of reversal, demonstrating that individuals learned the reversal task as well (Figure 5, P-value: 0.040).
Reaching criterion involves six consecutive successes across six trials without more than one error. Therefore, reaching criterion is defined by the mean success rate across six trials equaling minimum of .833, or 5/6. We were able to graph this approach to criterion by averaging the success rate across each increment of six trials (i.e. Trials 1-6, 2-7, 3-8, etc.) for each individual (Figure 6). For each individual, the mean of their success gradually increases towards .833 in their final increment of six trials, as each reaches criterion for the acquisition. DISCUSSION
been previously unexplored and unknown in anuran cognition and learning studies. These results confirm our hypothesis that the túngara frog is able to associate an arbitrary auditory cue with a reward and use this cue to complete a task.
As mentioned earlier, the behavioral flexibility of anurans has gone relatively unstudied in the literature, especially when considering primarily auditory tasks, as the majority of learning studies in anurans that currently exist focus primarily on visual tasks. It has been demonstrated that fire-bellied toads (Bombina orientalis), bullfrogs, newts, and túngara frogs have the ability (although occasionally limited) to associate a reward with a visual cue (Schmajuk, 1980, Jenkin, 2010, Ellins, 1982, Brattstrom 1990, Liu, 2014). However, the use of auditory learning in anurans is nearly absent. There are several studies revolving around auditory learning in mammals, including that of rats and pigs. Rats have effectively demonstrated the ability to associate a reward with a pulsation rate of a tone (Bao, 2004). Our study reflects an important movement away from the current focus on visual studies, opening the door to future auditory learning research in not only mammals, but also anuran species.
Another critical component of this study is its focus on reversal learning. Reversal learning involves the adaptation of behavior according to changes in the stimulus-rewards pairing, a learning capacity that is important to higher learning (Rolls, 1999). It is exemplified by a task in which subjects must learn to respond according to the opposite, previously
auditory cues in this study, including the reversal. This suggests that the túngara frog (and perhaps other amphibian species) has higher learning capabilities when considering an auditory modality as opposed to the visual modality, most likely as a result of their dependence on auditory communication and phonotaxis for survival and reproduction.
CONCLUSION
In the 1960’s, amphibians were believed to be behaviorally inflexible and incapable of learning (Hodos, 1969). However, the aforementioned visual studies have demonstrated the need for biologically relevant methodologies in order to reflect learning capabilities in anurans. Our study uses the biologically deterring factors of heat and lack of humidity to produce an
environment conducive to learning in amphibians, and demonstrates their ability to learn a task (and the reverse) using an auditory cue instead of the typical visual cue. Our study challenges the previously held conception of anurans as a behaviorally inflexible order of species and opens the door to further studies regarding their behavioral learning capabilities outside of the visual sense.
Figure 2: Success Rate (Averaged Across Individuals) Across Days for the Acquisition Period.
Figure 4: Latency (Averaged Across Individuals) Across Days in the Acquisition Period.
Figure 6: Approach to Criterion (Success Rate = .833) for Each Individual. REFERENCES
1. Bao, S., Chang, E.F., Woods, J., Merzenich, M. (2004). Temporal Plasticity in the Primary Auditory Cortex Induced by Operant Perceptual Learning. Nature: Neuroscience, 7(9), 974-981.
2. Brattstrom, B.H. (1990). Maze Learning in the Fire-Bellied Toad, Bombina orientalis.
Journal of Herpetology, 24(1), 44-47.
3. Colwill, R.M., Raymond, M.P., Ferreira, L., Escuerdo, H. (2005). Visual Discrimination Learning in Zebrafish (Danio rerio). Behavioral Processes, 70 (1), 19-31.
4. Devore, S., Lee, J., Linster, C. (2013). Odor Preferences Shape Discrimination Learning in Rats. Behavioral Neuroscience, 127(4), 498-504.
5. Ellins, S.R., Cramer, R.E., Martin, G.C. (1982). Discrimination Reversal Learning in Newts.
6. Giachetti, P., Orsini, C., Fogassi, L., Francesconi, W., Musumeci, D. (1985). An Automated Apparatus for the Study of Visual Discrimination in Pigeons. Behavioral Brain Research, 15(3), 255-258.
7. Giurfa, M. (2004). Conditioning Procedure and Color Discrimination in the Honeybee Apis
mellifera. Naturwissenschaften, 91, 228-231.
8. Harrison, J.M. (1984). The Functional Analysis of Auditory Discrimination . The Journal of
the Acoustical Society of America, 75(6), 1848-1854.
9. Hodos, W., Campbell, C.B.G. (1969). Scala Naturae: Why There is No Theory in Comparative Psychology. Psychological Review, 76(4), 337-350.
10. Kelber, A. (2002). Pattern Discrimination in the Hawkmoth: Innate Preferences, Learning Performance and Ecology. Proceedings of the Royal Society B: Biological Sciences, 269(1509), 2573-2577.
11. Liu, Y., Burmeister, S. (2014). The Females are Smarter: Sex Differences in Behavioral Flexibility in the Túngara Frog. Animal Behaviour (Under Review).
12. Murphy, E., Kraak, L., Nordquist, R.E., van der Staay, F.J. (2013). Successive and Conditional Learning in Pigs. Animal Cognition, 16(6), 883-893.
13. Rolls, E.T., Everitt, B.J., Roberts, A. (1996). The Orbitofrontal Cortex [and Discussion].
Philosophical Transactions: Biological Sciences, 351(1346), 1433-1444.
14. Schmajuk, N.A., Segura, E.T., Reboreda, J.C. (1980). Appetitive Conditioning and Discriminatory Learning in Toads. Behavioral Neural Biology, 28(4), 392-397. 15. Sinsch, U. (2006). Orientation and Navigation in Amphibia. Marine and Freshwater
Behavior and Physiology, 39(1), 65-71.