324
means of transport vesiclesthat shuttle between the various organelles. These transport vesicles carry membrane lipids and membrane-bound proteins to their proper destinations in the cell, and they also carry soluble materials destined for secretion. Thus, these organelles and the vesicles connecting them make up a single dynamic system of membranes and internal spaces. Currently, one of the most exciting questions in modern cell biology concerns endomembrane trafficking: How does each of the multitude of proteins and lipids in a cell manage to reach its proper destination at the proper time?
The Endoplasmic Reticulum
The endoplasmic reticulum (ER)is a continuous network of flattened sacs, tubules, and associated vesicles that stretches throughout the cytoplasm of the eukaryotic cell. Although the name sounds formidable, it is actually quite descriptive. Endoplasmicsimply means “within the (cyto)plasm,” and reticulum is a Latin word meaning “network.” The membrane-bounded sacs are called ER cisternae(singular: ER cisterna), and the space enclosed by them is called the ER lumen(Figure 12-1). Of the total membrane in a mammalian cell, up to 50–90% surrounds the ER lumen. Unlike more prominent organelles, such as the mitochondrion or chloroplast, however, the ER is not visible by light microscopy unless one or more of its com-ponents are stained with a dye or labeled with a fluorescent molecule.
The ER was first observed in the late nineteenth century, when it was noted that some eukaryotic cells, particularly those involved in secretion, contained regions that stained intensely with basic dyes. The signif-icance of these regions remained in doubt until the 1950s, when the resolving power of the electron micro-scope was improved dramatically. This allowed cell biologists to visualize for the first time the ER’s elaborate network of intracellular membranes and to investigate the role of the ER in cellular processes. This is a common
A
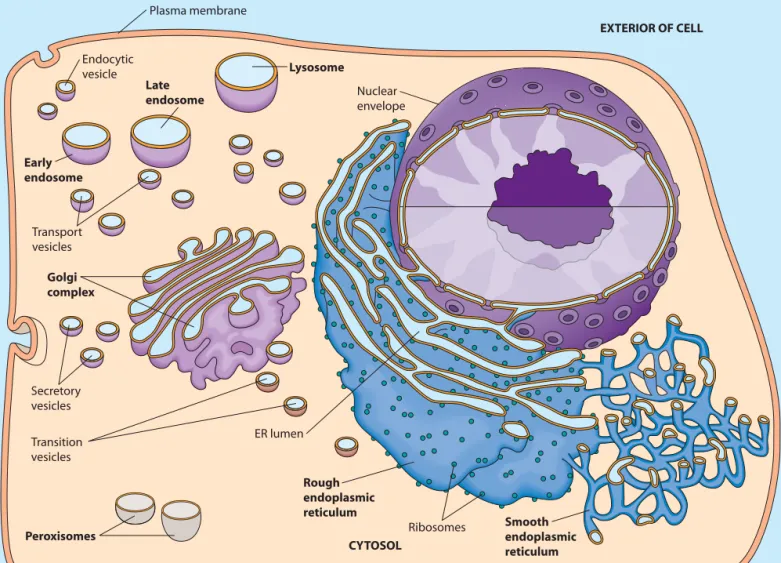
full appreciation of eukaryotic cells depends on an understanding of the prominent role of intracellular membranes and the com-partmentalization of function within organelles—intracellular membrane-bounded compart-ments that house various cellular activities. Whether we consider the storage and transcription of genetic informa-tion, the biosynthesis of secretory proteins, the breakdown of long-chain fatty acids, or any of the numerous other metabolic processes occurring within eukaryotic cells, many of the reactions of a particular pathway occur within a dis-tinct type of organelle. Also, the movement of molecules between organelles, known as trafficking,must be tightly regulated to ensure that each organelle has the correct com-ponents for its proper structure and function.We briefly encountered the major organelles found in eukaryotic cells in Chapter 4, and we then learned more about the mitochondrion and chloroplast in Chapters 10 and 11, respectively. We are now ready to consider several other individual organelles in more detail. We will begin with the rough endoplasmic reticulum, thesmooth endo-plasmic reticulum,and the Golgi complex,which are sites for protein synthesis, processing, and sorting. Next, we will look at endosomes,organelles that are important for carry-ing and sortcarry-ing material brought into the cell. Endosomes help to form lysosomes,which are organelles responsible for digestion of both ingested material and unneeded intracel-lular components. We will conclude with a look at peroxisomes,which house hydrogen peroxide–generating reactions and perform diverse metabolic functions.
As you study the role of each organelle, keep in mind that the endoplasmic reticulum, the Golgi complex, endo-somes, and lysosomes (but not peroxisomes) comprise the endomembrane systemof the eukaryotic cell, as shown in
Figure 12-1. (The nuclear envelope,which we will study in Chapter 18, is closely associated with the endomembrane system.) Material flows from the endoplasmic reticulum to and from the Golgi complex, endosomes, and lysosomes by
12
The Endomembrane System
and Peroxisomes
The Endoplasmic Reticulum 325 theme in scientific discovery—conceptual advances in
one field often follow technological advances in a related (or even an unrelated) field.
We now know that enzymes associated with the ER are responsible for the biosynthesis of proteins destined for incorporation into the plasma membrane or into organelles of the endomembrane system and for synthesis of proteins destined for export from the cell. The ER also plays a central role in the biosynthesis of lipids, including triacylglycerols, cholesterol, and related compounds. The ER is the source of most of the lipids that are assembled to form intracellular membranes and the plasma membrane. The Two Basic Kinds of Endoplasmic Reticulum Differ in Structure and Function
The two basic kinds of endoplasmic reticulum typically found in eukaryotic cells are distinguished from one
another by the presence or absence of ribosomes attached to the ER membrane (Figure 12-2). Rough endoplasmic reticulum (rough ER) is characterized by ribosomes attached to the cytosolic side of the membrane (the side that faces away from the ER lumen; Figure 12-2a). Trans-lation by these ribosomes occurs in the cytosol, but the newly-synthesized proteins will enter the ER lumen shortly. Because ribosomes contain RNA, it was this RNA that reacted strongly with the basic dyes originally used to identify the rough ER. A subdomain of rough ER, the transitional elements (TEs), plays an important role in the formation of transition vesiclesthat shuttle lipids and proteins from the ER to the Golgi complex. In contrast, smooth endoplasmic reticulum (smooth ER) appears smooth due to the absence of ribosomes attached to the membrane (Figure 12-2b) and has other roles in the cell.
Rough and smooth ER are easily distinguished mor-phologically. As illustrated in Figure 12-1, rough ER
Plasma membrane
Endocytic vesicle
ER lumen
EXTERIOR OF CELL
Nuclear envelope
Transport vesicles
Transition vesicles
Lysosome
Secretory vesicles
CYTOSOL Late
endosome
Early endosome
Golgi complex
Smooth endoplasmic reticulum Rough
endoplasmic reticulum
Peroxisomes Ribosomes
FIGURE 12-1 The Endomembrane System. The endomembrane system of the eukaryotic cell consists of the endoplasmic reticulum (ER), the Golgi complex, endosomes, and lysosomes (but not peroxisomes). It is associated with both the nuclear envelope and the plasma membrane. The ER lumen is linked to the interiors of the Golgi complex, endosomes, and lysosomes by transport vesicles that shuttle material between organelles, as well as to and from the plasma membrane.
326 Chapter 12 The Endomembrane System and Peroxisomes
(a) Rough endoplasmic reticulum 0.25μm (b) Smooth endoplasmic reticulum 0.25μm
FIGURE 12-2 Rough and Smooth Endoplasmic Reticulum. (a)Electron micrograph of endoplasmic reticulum. The rough ER is studded with ribosomes. (b)Electron micrograph of smooth endoplasmic retic-ulum. The dark spots near the smooth ER appear to be glycogen granules (TEMs).
membranes usually form large flattened sheets, whereas smooth ER membranes generally form tubular structures. The transitional elements of the rough ER are an excep-tion to this rule; they often resemble smooth ER. However, the rough and smooth ER are not separate organelles—electron micrographs and studies in living cells show that their lumenal spaces are continuous. Thus, material can travel between the rough and smooth ER without the aid of vesicles.
Both types of ER are present in most eukaryotic cells, but there is considerable variation in the relative amounts of each type, depending on the activities of the particular cell. Cells involved in the biosynthesis of secretory proteins, such as liver cells and cells producing digestive enzymes, tend to have very prominent rough ER networks. On the other hand, cells producing steroid hormones, such as in the testis or ovary, contain extensive networks of smooth ER.
When tissue is homogenized for subcellular fraction-ation, the ER membranes often break into smaller fragments that spontaneously close to form sealed vesicles known as microsomes.Fractions can be isolated with and without attached ribosomes, depending on whether the membrane originated from rough or smooth ER, respec-tively. Such preparations are tremendously useful for exploring both types of ER. Keep in mind, however, that microsomes do not exist in the cell; they are simply an artifact of the fractionation process. Box 12A presents more detailed information about subcellular fractionation by differential centrifugation.
Rough ER Is Involved in the Biosynthesis and Processing of Proteins
The ribosomes attached to the cytosolic side of the rough ER membrane are responsible for synthesizing both membrane-bound and soluble proteins for the endomem-brane system. So how do proteins enter the ER lumen and
the endomembrane system from their site of synthesis on the opposite (cytosolic) side of the rough ER membrane? Synthesis of proteins destined for the endomembrane system begins on cytoplasmic ribosomes, which attach to the rough ER via receptor proteins in the ER membrane shortly after translation initiation. Newly-synthesized pro-teins enter the endomembrane system cotranslationally— that is, they are inserted through a pore complex in the ER membrane into the rough ER lumen as the polypeptide is synthesized by the ER-bound ribosome (see Figure 22-16). After biosynthesis, membrane-spanning proteins remain anchored to the ER membrane either by hydrophobic regions of the polypeptide or by covalent attachment to membrane lipids. Soluble proteins, including secretory proteins, are released into the ER lumen.
In addition to its role in the biosynthesis of polypep-tide chains, the rough ER is the site for several other processes, including the initial steps of addition and processing of carbohydrate groups to glycoproteins, the folding of polypeptides, the recognition and removal of misfolded polypeptides, and the assembly of multimeric proteins. Thus, ER-specific proteins include a host of enzymes that catalyze cotranslational and posttranslational modifications. These modifications include glycosylation, which is important for sorting of proteins to their proper destinations, and disulfide bond formation, which is essential for proper protein folding. We will discuss the topics of protein biosynthesis, targeting, and folding in more detail in Chapter 22.
The ER is also a site for quality control. In ER-associated degradation (ERAD),proteins improperly modified, folded, or assembled are exported from the ER for degradation by cytosolic proteasomes before they can move on to the Golgi complex. Several human diseases, including cystic fibrosis and familial hyperc-holesterolemia, are associated with defects in these processes.
330 Chapter 12 The Endomembrane System and Peroxisomes Smooth ER Is Involved in Drug Detoxification, Carbohydrate Metabolism, Calcium Storage, and Steroid Biosynthesis
Drug Detoxification.Drug detoxification often involves enzyme-catalyzed hydroxylation because the addition of hydroxyl groups to hydrophobic drugs makes them more soluble and easier to excrete from the body. Hydroxylation of organic acceptor molecules is typically catalyzed by a member of the cytochrome P-450family of proteins. These proteins are especially prevalent in the smooth ER of hepato-cytes (liver cells), in which many drugs are detoxified.
In the hepatocytes, an electron transport system trans-fers electrons from NADPH or NADH to a heme group in a cytochrome P-450 protein, which then donates an electron to molecular oxygen. One atom of molecular oxygen gains two electrons and two , forming . The other oxygen atom is added to the organic substrate molecule as part of a hydroxyl group. Because one of the two oxygen atoms of is incorporated into the reaction product, these cytochrome P-450 enzymes are often called monooxygenases. The net reaction is shown below, where R represents the organic hydroxyl acceptor:
(12-1) The elimination of hydrophobic barbiturate drugs, for example, is enhanced by hydroxylation enzymes in the smooth ER. Injection of the sedative phenobarbital into a rat causes a rapid increase in the level of barbiturate-detoxifying enzymes in the liver, accompanied by a dramatic prolifera-tion of smooth ER. However, this means that increasingly higher doses of the drug are necessary to achieve the same sedative effect, an effect known as tolerancethat is seen in habitual users of phenobarbital. Furthermore, the enzyme induced by phenobarbital can hydroxylate and therefore sol-ubilize a variety of other drugs, including such useful agents as antibiotics, anticoagulants, and steroids. As a result, the chronic use of barbiturates decreases the effectiveness of many other clinically useful drugs.
Another cytochrome P-450 protein found in the smooth ER is part of an enzyme complex called aryl hydrocarbon hydroxylase. This complex is involved in metabolizing polycyclic hydrocarbons, organic molecules composed of two or more linked benzene rings that are often toxic. Hydroxylation of such molecules is important for increasing their solubility in water, but the oxidized products are often more toxic than the original com-pounds. Aryl hydrocarbon hydroxylase converts some potential carcinogens into their chemically active forms. Mice synthesizing high levels of this hydroxylase have a higher incidence of spontaneous cancer than normal mice do, whereas mice treated with an inhibitor of aryl hydrocarbon hydrolase develop few tumors. Significantly, cigarette smoke is a potent inducer of aryl hydrocarbon hydroxylase.
ROH + NAD(P)+ + H2O
RH + NAD(P)H + H+ + O2 ¡
O2
H2O
H+
Recent work shows that differences in activities and side effects of certain medications can result from differ-ences in the presence or activity of particular cytochrome P-450 genes in different patients. This has led to a new field of study known as pharmacogenetics(also called pharmaco-genomics), which investigates how inherited differences in genes (and their resulting protein products) can lead to dif-ferential responses to drugs and medications.
Carbohydrate Metabolism.The smooth ER of hepato-cytes (liver cells) is also involved in the enzymatic breakdown of stored glycogen, as evidenced by the pres-ence of glucose-6-phosphatase,a membrane-bound enzyme that is unique to the ER. Thus, its presence is used as a marker to identify the ER during subcellular fractionation or to visualize the ER using fluorescent antibodies. Glucose-6-phosphatase hydrolyzes the phosphate group from glucose-6-phosphate to form free glucose and inor-ganic phosphate :
(12-2) This enzyme is abundant in the liver because a major role of the liver is to keep the level of glucose in the blood relatively constant. The liver stores glucose as glycogen in granules associated with smooth ER (Figure 12-3a). When glucose is needed by the body, especially between meals and in response to increased muscular activity, liver glycogen is broken down by phosphorolysis (see Figure 9-10), producing glucose-6-phosphate (Figure 12-3b). Because membranes are generally impermeable to phos-phorylated sugars, the glucose-6-phosphate must be converted to free glucose by glucose-6-phosphatase in order to leave the cell and enter the bloodstream. Free glucose then leaves the liver cell via a glucose transporter (GLUT2) and moves into the blood for transport to other cells that need energy. Significantly, glucose-6-phos-phatase activity is present in liver, kidney, and intestinal cells but not in muscle or brain cells. Muscle and brain cells retain glucose-6-phosphate and use it to meet their own substantial energy needs.
Calcium Storage.The sarcoplasmic reticulumfound in muscle cells is an example of smooth ER that specializes in the storage of calcium. In these cells, the ER lumen contains high concentrations of calcium-binding proteins. Calcium ions are pumped into the ER by ATP-dependent calcium ATPasesand are released in response to extracellular signals to aid in muscle contraction (see Figure 14-12). Binding of neurotransmitter molecules to receptors on the surface of the muscle cell triggers a signal cascade that leads to the release of calcium from the sarcoplasmic reticulum and causes the contraction of muscle fibers. We will discuss nerve impulse transmis-sion and muscle contraction in more detail in Chapters 13 and 16.
glucose-6-phosphate +H2O ¡glucose +Pi
(Pi)
The Endoplasmic Reticulum 331 (a)Proximity of glycogen
to smooth ER
0.5μm
Glycogen granules
Smooth ER Mitochondrion
Process of glycogen breakdown in liver P
P
Glucose-1-P
Glucose Glycogen phosphorylase
LIVER CELL
Glycogen granule
Glucose transporter
Smooth ER
Plasma membrane
In blood Phosphoglucomutase
Glucose-6-P
Glucose Glucose-6-phosphatase
Pi
(b) FIGURE 12-3 The Role of the Smooth ER in the Catabolism
of Liver Glycogen. (a)This electron micrograph of a monkey liver cell shows numerous granules of glycogen closely associated with smooth ER (TEM). (b)The breakdown of liver glycogen involves the stepwise removal of glucose units as glucose-1-phosphate, followed by the conversion of 1-phosphate to glucose-6-phosphate by enzymes in the cytosol. Removal of the phosphate group depends on glucose-6-phosphatase, an enzyme associated with the smooth ER membrane. Free glucose is then transported out of the liver cell into the blood by a glucose transporter in the plasma membrane.
Steroid Biosynthesis.The smooth ER in certain cells is the site of biosynthesis of cholesterol and steroid hor-mones such as cortisol, testosterone, and estrogen. Large amounts of smooth ER are found in the cortisol-producing cells of the adrenal gland; the Leydig cells of the testes, which produce testosterone; the cholesterol-producing cells of the liver; and the follicular cells of the ovary, which produce estrogen. Smooth ER has also been found in close association with plastids in some plants, where it may be involved in phytohormone synthesis.
Cholesterol, cortisol, and the male and female steroid hormones just described share a common four-ring struc-ture but differ in the number and arrangement of carbon side chains and hydroxyl groups (see Figure 3-27e and Figure 3-30). Hydroxymethylglutaryl-CoA reductase (HMG-CoA reductase), the committed step in cholesterol biosynthesis, is present in large amounts in the smooth ER of liver cells. This enzyme is targeted for inhibition by a class of cholesterol-lowering drugs known as statins. In addition, the smooth ER contains a number of P-450 monooxygenases that are important not only in the syn-thesis of cholesterol but also in its conversion into steroid hormones by hydroxlyation.
The ER Plays a Central Role in the Biosynthesis of Membranes
In eukaryotic cells, the ER is the primary source of mem-brane lipids, including phospholipids and cholesterol. Indeed, most of the enzymes required for the biosynthesis of
membrane phospholipids are found nowhere else in the cell. There are, however, important exceptions. Mitochondria synthesize phosphatidylethanolamine by decarboxylating imported phosphatidylserine. Peroxisomes have enzymes to synthesize cholesterol, and chloroplasts contain enzymes for the synthesis of chloroplast-specific lipids.
Biosynthesis of fatty acids for membrane phospholipid molecules occurs in the cytoplasm and incorporation is restricted to the monolayer of the ER membrane facing the cytosol. Cellular membranes, of course, are phospholipid bilayers,with phospholipids distributed to both sides. Thus, there must be a mechanism for transferring phospholipids from one layer of the membrane to the other. Because it is thermodynamically unfavorable for phospholipids to flip spontaneously at a significant rate from one side of a bilayer to the other, transfer depends on phospholipid transloca-tors,also called flippases,which catalyze the translocation of phospholipids through ER membranes (see Figure 7-10). Phospholipid translocators, like other enzymes, are quite specific and affect only the rate of a process. Therefore, the type of phospholipid molecules transferred across a membrane depends on the particular translocators present, contributing to the membrane asymmetry described in Chapter 7. For example, the ER membrane contains a translocator for phosphatidylcholine, and thus it is found in both monolayers of the ER membrane. In contrast, there is no translocator for phosphatidylethanolamine, phos-phatidylinositol, or phosphatidylserine, which are therefore confined to the cytosolic monolayer. When vesicles from the ER membrane fuse with other organelles of the