*Correspondence to Author: Sayed, S.M.
Egypt- Animal Health Research In-stitute (Assiut Regional Lab., Bac-teriology Dept.).
E-mail: smhamuda @ yahoo.com, Phone: 01148278056
How to cite this article:
Sayed, S.M.. A contribution on Co-liforms causing mastitis in cows with reference to serotypes and virulence factors of E. coli isolates. American Journal of Microbiology and Immunology, 2016,1:2
Accepted 3 August 2016; published 3 August 2016.
eSciPub LLC, Houston, TX USA. Website: http://escipub.com/
American Journal of Microbiology and Immunology
(ISSN:2
474
-2
9
10)
Research Article AJMI (2016), 1:2
A contribution on Coliforms causing mastitis in cows with reference
to serotypes and virulence factors of E. coli
isolates
Escherichia coli (E. coli) is the predominant coliform species
causing intramammary infections. Where in the present study, E.
coli isolates were 18 strains (17.82%) followed by Enterobacter
aerogenes 3 strains (2.97%) and Klebsiella pneumoniae one
strain (0.99%) from 101 clinical mastitic milk samples of cows.
Eighteen E. coli isolates were serotyped to nine different sero
-groups; O111:H4 (3), O127:H6 (3), O26 (2), O126 (2), O119:H6
(1), O114:H21 (1), O55:H7 (1), O44:H18 (1), O124 (1) and (3)
untyped. Virulence tests were performed on the 18 isolated E.
coli, it was found that 15 isolates (83.3%) were serum resistant,
13 isolates (72.2%) had Congo Red binding activity, 6 isolates
(33.3%) were invasive and one isolate (5.6%) had haemolytic
activity. PCR was applied to detect the presence of Shiga like
toxin producing E. coli (stx1 and stx2 genes) on the nine differ
-ent strains (one strain for each serogroup), where stx1 and stx2
were found in 8 (88.9%) and 4 (44.4%) of the nine examined
strains, respectively. While stx1 and stx2 genes were found
to-gether in 3 strains (33.3%). Conclusions: E. coli isolates usually
posses one or more virulence factors that may help in
establish-ment at the infection site and subsequently causing clinical
bo-vine mastitis.
Keywords:
Coliforms, E. coli, Serotypes, Virulence Factors, Stx1, Stx 2.
Sayed, S. M.
Sayed, S.M., Egypt- Animal Health Research Institute (Assiut Regional Lab., Bacteriology Dept.)
Introduction
Mastitis is one of the major challenges of the dairy industry. Staphylococcus aureus [S. aureus] is one of the most important pathogens causing mastitis in dairy cattle [1,2]. Methicillin resistant Staphylococcus aureus [MRSA] has been recovered from dairy cat-tle in Korea [1-3]; Turkey [4]; Netherland [5]; Iran [6] and Uganda [7]. Several efforts to remove this pathogen from farms are hampered by some factors, where one of these factors is antibiotic resistance. One of the major mechanisms of resistance to β-lactam an -tibiotics is β-lactamase producing by staphylococci. This enzyme hydrolyzes the β- lactam ring and caus -es inactivation of β-lactams. In the early 1950s, it has been aware of the effectiveness of penicillin in treat -ment of S. aureus infections because of β-lactamase producing plasmids. In 1959, methicillin, synthetic, penicillinase –resistant penicillin, was introduced and solved problems in clinical practice, for a time. How-ever, by 1960, S. aureus strains were found to be resistant to the new semisynthetic β-lactams [methi -cillin, oxa-cillin, flucloxacillin], and became known as methicillin-resistant S. aureus [MRSA]. This type of resistance was termed “intrinsic resistance” because it was not due to destruction of the antibiotic by β-lac -tamase [8]. Methicillin resistance in S. aureus is medi-ated by the production of an altered penicillin-binding protein [PBP2a], a transpeptidase. mecA encodes this enzyme involved in cell wall peptidoglycan synthesis. Unlike conventional PBPs of S. aureus, PBP2a does not bind to β-lactam antibiotics with high affinity [9]. It is considered that the first step in mastitis progress is adhesion of S. aureus to mammary epithelial cells and slime factor plays an important role for adhesion and colonization [10]. Production of slime factor also plays an important role in antibiotic resistance and it has been reported that slime producing strains are more resistant to antibiotics than non-slime produc-ing strains [11]. Intercellular adhesion is encoded in the ica locus containing icaA, icaB, icaC, icaD genes in S. aureus strains [12]. icaA gene encodes N-acetyl-glucosaminyl transferase, further, icaD plays an im-portant role in expression of this enzyme. icaA and icaD were found to be in high prevalence among S. aureus mastitis isolates and this finding confirms that ica locus has a potential role as a virulence fac-tor in the pathogenesis of mastitis in ruminants [10]. This study was undertaken to determine the bovine mastitis Staph. Spp., their resistance to antimicrobi-al agents approved for its control and to determine
tion of S. aureus in bovine mastitis phenotypically and genotypically for mecA, icaA and icaD genes.
Materials and Methods
Milk samplesA total of 101 milk samples were collected from 101 cows, at various private farms in Assiut, Egypt, showing clinical signs of mastitis. All samples were taken under aseptic conditions and trans-ferred in ice box to laboratory as soon as possible.
Isolation and identification of bacterial isolates
Amount of 0.01 ml of each milk samples was cultured on blood agar with 5% sheep blood, Mannitol salt agar [BBL], Baird-Parker medium [Oxoid] and Mac-Conkey agar [Biomark Lab. India] which incubated at 37°c for 48 h. The suspected colonies were identi-fied: morphologically, by Gram’s stain and biochemi -cally confirmed by using catalase activity, coagulase test as well as Novobiocin [5 µg] and polymixin-ß sulphate [300 U] sensitivity tests, according to [13].
Phenotypic detection of methicillin resistance Disc diffusion sensitivity testing was performed ac -cording to the Kirby-Bauer method, as described in the guidelines of the National Committee for Labo-ratory Standards [14], using discs [Bioanalyse-Tur-key] containing Oxacillin [OX] 1 µg, Ampicilin [AM] 10 µg, Cefotaxime [CTX] 30 µg, Cloxacillin [CX] 1 µg, Doxycycline [DO] 30 µg, Enrofloxacin [ENR] 5 µg, Gentamicin [CN] 10 µg, Lincomycin [L] 2 µg, Oxytetracycline [T] 30 µg, Penicillin [P] 10 µ and Trimethoprim – Sulflamethaxzole [SXT] 25 µg. For Oxacillin susceptibility determinations, inhibi-tion zones around the disc were measured after 24 and 48 h using the following breakpoints: sus-ceptible [S] ≥ 18 mm; resistance [R] ≤ 17 mm [13].
Slime production assay
col-Table 1 The sequence of stx1
PRIMER NAME TARGET GENE OLIGONUCLEOTIDE SEQUENCE (5′ → 3′)
1Slt224 stx1 ATG TCA GAG GGA TAG ATC CA
1Slt385 stx1 TAT AGC TAC TGT CAC CAG ACA AT
Table 2 The sequence of stx2
PRIMER NAME TARGET GENE OLIGONUCLEOTIDE SEQUENCE (5′ → 3′)
2Slt537 stx2 AGT TCT GCG TTT TGT CAC TGT C
2Slt678b stx2 CGG AAG CAC ATT GCT GAT T
Table 3: Prevalence of coliforms isolated from 101 clinical mastitis cow’s milk samples.
E. coli Enterobacter aerogenes Klebsiella pneumoniae Total of coliforms
No. % No. % No. % No. %
18 17.82 3 2.97 1 0.99 22 21.78
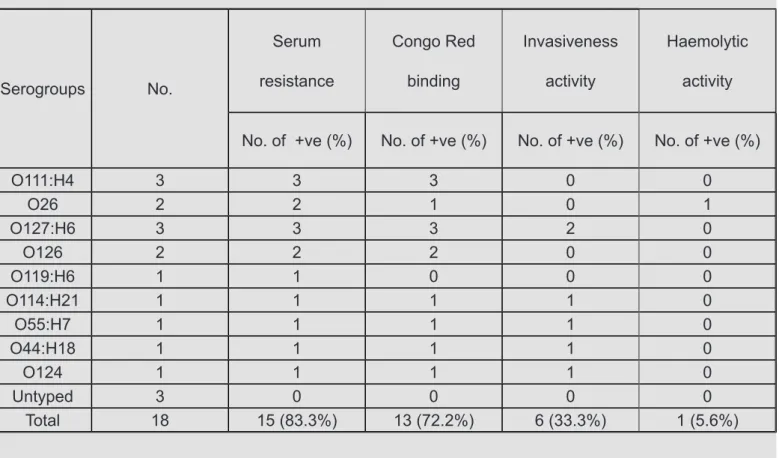
Table 4 Relationship between different serogroups and phenotypic virulence factors of E. coli isolated from clinical mastitic cow’s milk samples.
Serogroups No.
Serum
resistance
Congo Red
binding
Invasiveness
activity
Haemolytic
activity
No. of +ve (%) No. of +ve (%) No. of +ve (%) No. of +ve (%)
O111:H4 3 3 3 0 0
O26 2 2 1 0 1
O127:H6 3 3 3 2 0
O126 2 2 2 0 0
O119:H6 1 1 0 0 0
O114:H21 1 1 1 1 0
O55:H7 1 1 1 1 0
O44:H18 1 1 1 1 0
O124 1 1 1 1 0
Untyped 3 0 0 0 0
PCR for detection of mecA, icaA and icaD genes
Detection of mecA, icaA and icaD genes was per-formed on those ten S. aureus isolates, which were methicillin resistant by phenotypic test, as follows:
I- DNA extraction: DNA extraction from samples was performed using the QIAamp DNA Mini kit [Qiagen, Germany, GmbH] with modifications from the manufacturer’s recommendations. Briefly, 200 µl of the sample suspension was incubated with 10 µl of proteinase K and 200 µl of lysis buffer at 56OC for 10 min. After incubation, 200 µl of 100%
ethanol was added to the lysate. The sample was then washed and centrifuged following the manu-facturer’s recommendations. Nucleic acid was elut -ed with 100 µl of elution buffer provid-ed in the kit. II- Oligonucleotide Primers: Primers encod-ing for mecA, icaA and icaD genes were supplied from [Metabion, Germany] are listed in Table [1].
III- PCR amplification: Primers were utilized in a 25- µl reaction containing 12.5 µl of Emerald Amp Max PCR Master Mix [Takara, Japan], 1 µl of each primer of 20 pmol concentration, 4.5 µl of water, and 6 µl of template. The reaction was performed in a Biometra thermal cycler. For mecA gene PCR, a primary dena-turation step was done at 95 OC for 5 min, followed by
35 cycles of 95OC for 45 sec., 50OC for 45 sec. and
72OC for 45 sec. A final extension step was done at
72OC for 10 min, according to[15]. However for the icaA
and icaD genes, the cycles consisted of 95ºC for 1 min, 49º C for 1 min and 72º C for 1 min, according to[4].
IV- Analysis of the PCR Products: The products of PCR were separated by electrophoresis on 1.5% agarose gel [Applichem, Germany, GmbH] in 1x TBE buffer at room temperature using gradients of 5V/cm. For gel analysis, 15 µl of the products was loaded in each gel slot. A 100 bp DNA Ladder [Qiagen, Germa -ny, GmbH] and 100 bp plus DNA Ladder [Fermentas, Cat.No. SM 0323] were used to determine the frag-ment sizes. The gel was photographed by a gel doc-umentation system [Alpha Innotech, Biometra] and the data was analyzed through computer software.
Results
Please check figures and tables.
Discussion
Table 5 Stx1 and stx2 genes profile of different nine E. coli strains isolated from clinical mastitic cow’s milk samples.
Virulence genes Positive serogroups Number of +ve serogroup (%)
stx1 O26, O44:H18, O55:H7, O111:H4, O114:H21, O119:H6, O126 and
O127:H7 8 (88.9%)
stx2 O26, O111:H4, O126 and O124 4 (44.4%)
stx1and stx2 O26, O111:H4 and O126 3 (33.3%)
Fig. 1 serum resistance test, A): control negative (green colour). B): positive serum re-sistance E. coli isolate (yellow colour).
M
1
2
3
4
5
6
7
8
9
10
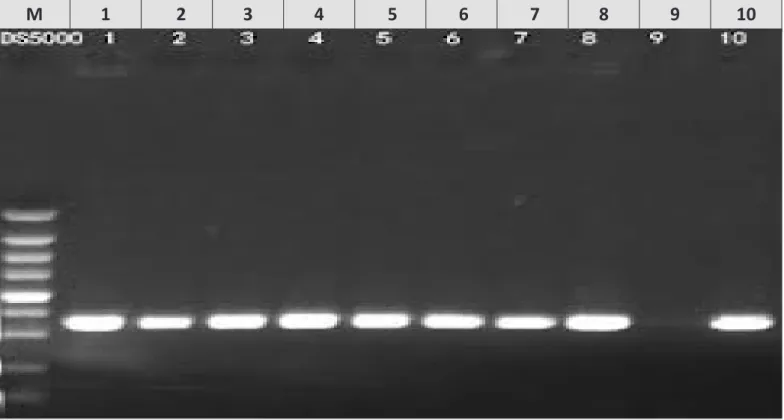
Fig. 3 Agarose gel electrophoresis of PCR amplification products using Shiga toxin 1 (stx1) primers of E. coli (1Slt224 and 1Slt385). Lane M: 185 bp ladder as molecular DNA marker. Lane 1: Control positive for stx1 producing E. coli. Lane 2 (E. coli O26), Lane 3 (E. coli O44), Lane 4 (E. coli O55), Lane 5 (E. coli O111), Lane 6 (E. coli O114), Lane 7 (E. coli O119), Lane 8 (E. coli O126) and Lane 10 (E. coli O127): Positive E. coli for stx1 production. Lane 9 (E. coli O124): Negative E. coli for stx1 production.
M
1
2
3
4
5
6
7
8
9
10
4], respectively. MRSA resistance to methicillin and other ß-lactam antibiotics is caused by the action of mecA gene [29]. The discrepant results between disc diffusion methods and PCR for detection of methicillin resistance may be due to another resistance mech-anism such as hyperproduction of beta-lactamase, also MRSA strains show a heterogeneous charac-ter with the level of resistance varying according to the culture conditions and β-lactam antibiotic being used. Because of this heterogeneous resistance, and time consuming the detection of MRSA by phe-notypic methods becomes problematic [4]. However, PCR-based methods have shown to be a rapid and reliable approach for the identification and genotypic characterization of MRSA. Detection of mecA- based PCR methods has accepted as “gold standard” [30]. The present work found that six isolates [60%] of the tested MRSA strains were slime producing positive on CRA plates in vitro, Table [4]. Slime-producing S. aureus isolates from different clinical origins such as bovine mastitis [4, 10], wound infection [12] and clinical cases [31] has been detected in vitro by using Con-go Red Agar plates in percentages of 37.2, 91.4, 52 and 53.3%, respectively. Phenotype on CRA was found to be an unreliable indicator of slime-form-ing capacity among clinical isolates of S. aureus [32]. Therefore, although CRA methods may be easier to perform than a molecular analysis of the genes impli-cated in biofilm production and could be performed easily in a diagnostic laboratory, it may be a poor method for determining the slime producing capac-ity of clinical isolates in the diagnostic laboratory [33]. PCR methods provided a direct evidence of the ge-netic basis of slime production complementary to the CRA test. The ica locus consists ica A, D, B, C genes. Slime synthesis is controlled by the ica [inter-cellular adhesion] operon. Coexpression of icaA with icaD leads to a significant increase in activity and is related to phenotypic expression of the capsular polysaccharide [34]. In this study, slime factor produc-tion of MRSA isolates were detected by PCR target-ing icaA and icaD genes and found that 5 [50%] of the tested MRSA strains were positive for both icaA and icaD genes. Six [60%] and eight [80%] isolates were positive for icaA gene and icaD gene, respec-tively as shown in Table [4] and Fig. [2 & 3]. Fifteen [25.42%] out of 59 S. aureus strains were positive for both icaA and icaD genes, in addition 16 [27.12%] and 38 [64.41%] out of the 59 strains were positive for icaA and icaD gene, respectively [4]. Also [31] found that 75% of MRSA carried ica operon. The icaAD
gene was detected in 32% of Staphylococcal spp. [35]. While [34] found that all strains which were positive for icaA gene were also positive for icaD gene. In ad-dition [10, 12] have reported that all S. aureus isolates possessed the ica locus, icaA and icaD genes. icaA and icaD genes were not be together in some isolates may due to some mutations on icaA gene, although coexpression on icaA and icaD is necessary for slime production it was considered that other genes in ica locus play role in controlling slime expression [4]. In present study, among four isolates which were negative for slime production on CRA plate in vitro, one isolate was positive for both icaA and icaD genes, two isolates were positive for icaD gene and the last one was negative for both icaA and icaD genes, Ta-ble [4]. Among 37 S. aureus strains which did not pro-duce slime factor on CRA plate in vitro, only 7 strains [18.9%] were positive for both icaA and icaD genes [4], they suggest that some environmental conditions or presence of accessory genes can influence the phe -notypic behavior on the Congo red agar plate, giving colonies which did not fully express the ica genes. In this work, six isolates [60%] were positive for both methicillin resistance and slime production pheno-typically and three strains [30%] were positive for all mecA, icaA and icaD genes, Table [4]. Only 2 [3.39%] of 59 S. aureus strains were positive for both methicil-lin resistance and slime producing, phenotypically [4]. In conclusion, it was showed that detection of mecA gene in S. aureus isolates indicating that sever-al cases suffering from S. aureus mastitis have an MRSA problem. Genotypic determination of mecA gene proved the most reliable method for detection of methicillin resistance. The present work paid an attention to the 3 MRSA strains [30%] were posi-tive to all tested genes rather than slime produc-tion as the worst isolated stains all over this study [multidrug resistant, slime producing as well as carrying mecA, icaA and icaD genes]. In vitro En-rofloxacin, Gentamicin and Doxycycline are the most effective drugs for Staph. spp. clinical mastitis.
Conflict of interest
NoneAll members shared in the work were entitled as authors but we want to acknowledge the molecular biotechnol-ogy department of AHRI - Dokky , reference Lab, Egypt.
References
1. Koneman, E. W.; Allen, S. D.; Dowell V. R. and Som-mer H. M. The Enterobacteriaceae, in: Color Atlas and Textbook of Diagnostic Microbiology, J.B. Lippin-cott Company, New York, USA, 1983; pp. 57– 124.
2. Hogan, J. S and Smith, K. L. Coliform mastitis. Vet. Res., 2003; 34:507-519.
3. Burvenich, C.; Van Merris, V.; Mehrzad, J. et al. Se-verity of E. coli mastitis is mainly determined by cow factors. Vet. Res., 2003; 34(5):521- 564.
4. Lehtolainen, T. (2004): E. coli mastitis: Bacterial fac-tors and host response, Ph. D. thesis Department of clinical veterinary sciences, Faculty of veterinary Medicine, Helsinki, University, Finland, 2004.
5. Quesnell, R. R.; Klaessig, S.; Watts, J. L. et al. Bovine intramammary Escherichia coli challenge infections in late gestation demonstrate a dominant anti-inflam -matory immunological response. J. Dairy Sci., 2012; 95(1):117-126.
6. Fernandes, JB. C.; Zanardo, L. G.; Galvão, N.N. et al. Escherichia coli from clinical mastitis: serotypes and virulence factors. J. of Vet. Diagnostic Investigation, 2011; 23(6) 1146–1152.
7. Kaper, J. B.; Nataro, J. P. and Mobley, H. L. Patho-genic E. coli. Nat. Rev. Microbiol., 2004; 2:123–140.
8. Moussa, I. M.; Mostafa, M. and Mohamed, K. F. De-termination of phylogenetic relationships among E. coli isolates recovered from bovine fecal and milk samples. J. Egypt. Vet. Med. Ass., 2006; 66: 7-25.
9. Amira, El-Sayed Lamey; Ammar, A. M.; Zaki, E. R.; et al. Virulence factors of Escherichia coli isolated from recurrent cases of clinical and subclinical
masti-tis in buffaloes. International J. of Microbiological Re -search, 2013; 4 (1): 86-94.
10. Dalia, M. Mohsen and Amany, N. Dapgh. Detection of gene sequence of E. coli toxins isolated from mastitic buffalo and cows milk. Vet. Med. J. Giza, 2007; 55(2): 451-467.
Congo red dye test as an indicator test for detection of invasive bovine Escherichia coli – short communi-cation. Veterinarski Arhiv, 2006; 76(4):363-366.
12. Taylor, P. w. Bacteriocidal and bacteriolytic activity of serum against Gram negative bacteria. Microbial. Rev., 1983; 47: 46-83.
13. John, M.; Jacques, M. and Lariviere, S. Pathogenicity of E. coli O115:K V165 strains isolated from pigs with diarrohea. Am. J. Vet. Res., 1989; 50(1): 1029-1036.
14. Gad EL-Said, W. A.; El-Jakee, J. K.; Xandel, M. M. et al. Presence of E. coli O157:H7 in raw milk and meat samples. J. Egypt Vet. Med. Ass., 2005; 65(3): 341-350.
15. Quinn, P. J.; Carter, M. E.; Markey, B. et al. Clinical veterinary microbiology. 6th ed., 2004; Mosby,
Edin-burgh, London, New York, Philadelphia, St. Louis, Sydney, Toronto.
16. Kok, T.; Worswich, D. and Gowans, E. (1996). Some serological techniques for microbial and viral infec-tions. In Practical Medical Microbiology (Collee, J.; Fraser, A.; Marmion, B. and Simmons, A., eds.) 14th
ed.,1996, Edinburgh, Churchill Livingstone, UK.
17. Beutin, L.; Montenegro, M. A.; Orskov, I. et al. Close association of verotoxin (Shiga like toxin) production with enterohaemolysin production in strains of E. coli. J. Clin. Microbiol. 1989; 27(11):2559-2564.
18. Panigrahy, B. and Yushen, L. Differentiation of patho -genic and non patho-genic Escherichia coli isolated from poultry. Avian Dis., 1990; 34: 941-943.
19. Timmis, K. N. Plasmid gene that specifies resistance to the bactericidal activity of serum. In Plasmids of medical, environmental and commercial importance. Edited by K. N. Timmis & Puhler, A. (Elsevier, North Holland Biomedical Press, New York,1979;145-153.
20. Sereny, B. Experimental shigella keratoconjunctivitis. Acta Microbiol. Acad. Sci. Hung., 1955; 2: 293-296.
21. Fagan, P.; Hornitzky, M.; Bettelheim, K. et al. Detec-tion of Shiga-like toxin (stx1 and stx2), intimin (eaeA), and enterohemorrhagic E. coli (EHEC) hemolysin (EHEC hlyA) genes in animal feces by multiplex PCR. Appl. Environ. Microbiol. 1999; 65:868-872.
23. Pass, M., Odedra, R. and Batt, R. Multiplex PCR for identification of Escherichia coli virulence genes. J. Clin. Microbiol., 2000; 38:2001-2004.
24. Oliver, S.P. Frequency of isolation of environmental mastitis-causing pathogens and incidence of new in-tramammary infection during the non-lactating peri-od. American Journal of Veterinary Research, 1988; 49(11): 1789-1793.
25. El-Khodery, S. A. and Osman, S. A. Acute coliform mastitis in buffaloes (Bubalus bubalis): Clinical find -ings and treatment outcomes. Trop. Animal Health and Production, 2008; 40(2): 93-99.
26. Wenz, J. R.; Barrington, G. M.; Garry, F. B. et al. E. coli isolates, serotypes, genotypes and virulence genes and clinical coliform mastitis severity. Journal of Dairy Science, 2006; 89(9): 3408-3412.
27. Blum, S. E. and Leitner, G. Genotyping and virulence factors assessment of bovine mastitis Escherichia coli. Vet. Microbiol., 2013;163(3-4):305-312.
28. Schukken, Y.H,; Bennett, G.J.; Zurakowski, M.J. et al. Randomized clinical trial to evaluate the efficacy of a 5-day ceftiofur hydrochloride intramammary treat-ment on non severe gram-negative clinical mastitis. J. Dairy Sci., 2011; 94(12):6203-6215.
29. Ericsson, U. H.; Lindberg, A.; Persson, W. K. et al. Microbial aetiology of acute clinical mastitis and agent-specific risk factors. Vet. Microbiol., 2009; 137(1-2):90-97.
30. Kalmus, P.; Aasmäe, B.; Kärssin, A. et al. Udder pathogens and their resistance to antimicrobial agents in dairy cows in Estonia. Acta Vet Scand., 2011; 53: 4.
31. Bradley, A.J. and Green, M.J. Aetiology of clinical mastitis in six Somerest dairy herds. Vet. Rec., 2001; 148(22):683-686.
32. Longo, F.; Salat, O. and Gool, V.F. Incidence of clin -ical mastitis in French dairy herds: epidemiolog-ical data and economic costs. Folia Veterinaria, 2001; 45: 45-46.
33. El-Mahrouki, A. M.; Nevine, M. Sobhy and Aggour, M. G. Detection of coliform mastitis in cattle with special reference to molecular characterization of enterotoxi-genic E. coli using polymerase chain reaction (PCR). J. Egypt. Vet. Med. Ass., 2006; 66(1): 47-58.
34. Rangel, P. and Marin, J. M. Analysis of
Escherich-ia coli isolated from bovine mastitic milk. Pesq. Vet. Bras., 2009; 29(5):363-368.
35. Breen, J. E.; Green, M. J. and Bradley, A. J. Quarter and cow risk factors associated with the occurrence of clinical mastitis in dairy cows in the United King-dom. J. Dairy Sci., 2009; 92(6): 2551–2561.
36. Gyles, C. L. Escherichia coli. In: Gyles, C. L., Thoen, C.O. (Eds.). Pathogenesis of bacterial infections in animals. Iowa State University Press, Ames, Iowa, 1993; pp. 164-187.
37. DaRocha, A.C.; DaSilva, A.B.; DeBrito, A.B. et al. Virulence factors of avian pathogenic E. coli isolat-ed from broilers from the South of Brazil. Avian Dis., 2002; 46(3):749-753.
38. Kaipainen, T.; Pohjanvitra, T.; Sphigel, N. Y. et al. Vir-ulence factors of E. coli isolated from bovine clinical mastitis. Vet. Microbiol., 2002; 85(1):37-46.
39. Zaki, E.R.; Riad, E. M. and Sobhy, N. M. Correlation between E. coli serotypes isolated from buffalo mas -titic milk with different virulence patterns. J. Egypt Vet. Med. Assoc., 2004; 64: 53-63.
40. Berkhoff, H. A. and Vinal, C. A. Congo red medium to distinguish between invasive and noninvasive Esch-erichia coli pathogenic for poultry. Avian Diseases, 1986; 30(1): 117-121.
41. Salman, A. M. Serological and bacteriological studies on Escherichia coli in chicken. Ph. D. Thesis (Microbi-ology), Fac. Vet. Med., Cairo Univ., 1999.
42. Mosherf, B. E. (2004): Studies on microbial causes of mastitis in buffaloes, Ph. D. thesis (Microbiology), Fac. Vet. Med. Cairo Univ. 2004.
43. Beutin, L.; Krause, G.; Zimmermann, S, et al. Char -acterization of Shiga toxin-producing Escherichia coli strains isolated from human patients in Germany over a 3-year period. J. Clin. Microbiol. 2004; 42:1099-1108.
44. Zschock, M.; Hamann, H. P.; Kloppert, B. et al. Shi-ga-toxin producing E.coli in faeces of healthy dairy cows, sheep and goats: prevalence and virulence properties. Lett. Appl. Microbiol., 2000; 31:203-208.
46. Lira, W. M.; Macedo, C. and Marin, J. M. The in-cidence of Shiga toxin-producing Escherichia coli in cattle with mastitis in Brazil. J. of Applied Microbiolo-gy, 2004; 9: 861–866.
47. Galal, H. M.; Hakim, A. S. and Sohad, M. D. Phe -notypic and virulence genes screening of Escherich-ia coli strains isolated from different sources in delta Egypt. Life Science Journal, 2013; 10(2):352-361.
48. Murinda, S.E.; Nquyen, L.T.; Landers, T.L.; et al. Comparison of E. coli isolates from humans, food and farm and companion animals for presence of Shiga toxin-producing E. coli virulence markers. Foodborne Pathogens and Disease, 2004; 1(3): 178-184.