Dorota Tichaczek-Goska
1, Danuta Witkowska
2, Agnieszka Cisowska
1,
Stanisław Jankowski
1, Andrzej B. Hendrich
1The Bactericidal Activity of Normal Human Serum
Against Enterobacteriaceae Rods with Lipopolysaccharides
Possessing O-Antigens Composed of Mannan*
Bakteriobójcza aktywność normalnej surowicy ludzkiej
wobec szczepów z rodziny Enterobacte
r
iaceae z lipopolisacharydami
zawierającymi O-antygeny typu mannanowego
1 Department of Biology and Medical Parasitology, Wroclaw Medical University, Wroclaw, Poland 2 Institute of Immunology and Experimental Therapy, Polish Academy of Sciences, Wroclaw, Poland
Abstract
Background. The susceptibility of bacteria to the bactericidal activity of serum depends on the structure and orga-nization of the bacterial outer membrane. It is known that the structure of the O-specific polysaccharide chain of lipopolysaccharide (LPS) plays an important role in the resistance of bacteria to host immune defenses.
Objectives. The susceptibility of rods belonging to Enterobacteriaceae family to the bactericidal activity of the nor-mal human serum (NHS) was examined. The mechanisms of complement activation were also investigated.
Material and Methods. The study was carried out on 15 strains containing LPSs with O-specific polysaccharides composed of mannan, belonging to the following species: Citrobacter freundii, C. werkmanii, C. braakii, C. youn-gae, Hafnia alvei, Escherichia coli and Klebsiella pneumoniae. The levels of C3 and C4 complement components, IgG and IgM immunoglobulin in NHS were examined using specific antibodies. The bactericidal activity of NHS and its preparations (HS50/20, HSMgEGTA) was determined. LPSs from E. coli O8 strains were analyzed by poly-acrylamide gel electrophoresis (PAGE) in the presence of sodium dodecyl sulphate (SDS).
Results. Eleven strains were sensitive to NHS bactericidal activity, and four were resistant. Only one group of strains was fully susceptible to NHS action. In three other groups, both sensitive and resistant strains were found. The majority of the strains remained susceptible to NHS activity irrespective of which pathway of serum activity was blocked. All E. coli O8 strains contained smooth-type LPSs.
Conclusions. Strains belonging to the same serotype showed variable susceptibility to the bactericidal action of normal human serum. Two mechanisms of the bactericidal activity of NHS have been identified (Adv Clin Exp Med 2012, 21, 3, 289–299).
Key words: mannan, O-antigen, human serum, bactericidal effect, complement system.
Streszczenie
Wprowadzenie. Podatność bakterii na bakteriobójcze działanie surowicy zależy od struktury i organizacji błony zewnętrznej drobnoustroju. Wiadomo, że struktura polisacharydowego łańcucha O-swoistego odgrywa istotną rolę w oporności bakterii na działanie układu odpornościowego organizmu gospodarza.
Cel pracy. Zbadano podatność pałeczek należących do rodziny Enterobacteriaceae na bakteriobójcze działanie normalnej surowicy ludzkiej (NSL). Określono również mechanizm aktywacji układu dopełniacza oraz jego rolę w procesie lizy komórek przez NSL.
Materiał i metody. Badaniami objęto grupę 15 szczepówmających O-antygeny typu mannanowego, należących do gatunków: Citrobacter freundii, C. werkmanii, C. braakii, C. youngae, Hafnia alvei, Escherichia coli i Klebsiella pneu-moniae. Oznaczano stężenia składowych dopełniacza C3 i C4 oraz przeciwciał klasy IgG i IgM w NSL. Określano
Adv Clin Exp Med 2012, 21, 3, 289–299
ISSN 1899–5276 © Copyright by Wroclaw Medical University
bakteriobójcze działanie NSL oraz jej preparatów (SL50/20, SLMgEGTA) w stosunku do testowanych pałeczek. Wyizolowano i charakteryzowano elektroforetycznie (SDS-PAGE) lipopolisacharydy (LPS) szczepów E. coli O8.
Wyniki. Jedenaście szczepów bakterii było podatnych, a cztery oporne na działanieNSL. Tylko jedna grupa pałe-czek była wrażliwa na działanie NSL. W pozostałych grupach drobnoustrojów, mimo że struktury O-antygenowe były identyczne, odnotowano zarówno podatne, jak i niewrażliwe na NSL szczepy. Większość bakterii nie zmie-niła swojego wzoru podatności na surowicę, niezależnie od tego, którą z dróg układu dopełniacza inaktywowano. Wszystkie szczepy E. coli O8 miały LPS typu gładkiego.
Wnioski. Szczepy bakteryjne należące do jednego gatunku i serotypu wykazują zróżnicowanie w podatności na lityczne działanie białek układu dopełniacza. Odnotowano dwa mechanizmy bakteriobójczego działania NSL wobec testowanych pałeczek (Adv Clin Exp Med 2012, 21, 3, 289–299).
Słowa kluczowe: mannan, O-antygen, ludzka surowica, działanie bakteriobójcze, układ dopełniacza.
The outer membrane is the first layer of the cell envelope of Gram-negative bacteria. The internal part is built of phospholipids, and the external part is built mainly of lipopolysaccharide (LPS) [1]. LPS contributes greatly to the structural integrity of the bacterial cell envelope. It is one of the virulence factors of bacteria. Lipopolysaccharides of some Enterobacteriaceae rods from the genera Citrobac-ter, Hafnia, Escherichia and Klebsiella possess the O-specific polysaccharide moieties consisting of mannan or including mannose [2–5].
Mannan is a polymer of monosaccharide man-nose. It is a storage product and component of many plant cells, and a major cell wall glycoprotein of Candida spp. [6]. The mycobacterial cell wall com-ponents lipomannan and lipoarabinomannan have been described as virulence factors of Mycobacte-rium tuberculosis[7]. The biological importance of mannan as a component of bacterial structures is relatively poorly described. It is known that man-nan strongly activates the complement system and the body’s immune response [8, 9].
As Podschun and Ullmann wrote, “As op-portunistic pathogens, Klebsiella spp. primar-ily attack immunocompromised individuals who are hospitalized and suffer from severe underly-ing diseases” [10]. Hafnia alvei strains have been shown to cause respiratory tract infections, diar-rhea and gastroenteritis [11]. Gupta et al. pointed out: “Citrobacter species cause a wide spectrum of infections in the urinary tract, blood, … peritone-um and several other normally sterile sites, most frequently in hospitalized and immunocompro-mised patients” [12]. Escherichia coli strains can cause urinary tract infections, and severe diarrhea in children and adults [13]. Escherichia coli and Klebsiella spp. bacilli are the most common bac-teria responsible for severe infections in humans, mainly in small children, e.g. in neonatal intensive care units [14]. rods belonging to the Enterobac-teriaceae family, especially those resistant to wide range of antibiotics, are an important epidemio-logical problem in Poland and elsewhere [15].
The bactericidal activity of serum is a natural barrier, and is a very important factor
protect-ing the macroorganism against various infec-tions caused by Gram-negative bacteria (among others) [16]. It is known that the structure of the O-specific polysaccharide chain of LPS plays an important role in the resistance of bacteria to host immune defenses, especially to complement protein deposition and complement lytic activ-ity [17–19]. There are three routes by which the complement system can be activated: the classical pathway, the alternative pathway and the lectin pathway. The lectin pathway is activated when mannan-binding lectin (MBL) present in the host serum attaches to mannose or to some other sug-ar residues (N-acetylglucosamine, fucose) on the pathogen surface. This activates the MBL- associ-ated serine proteases MASP-1 and MASP-2, and the latter cleaves C2 and C4 to generate a C3 con-vertase [20]. MBL also associates with MASP-3 and MAp19, although the function of them is still unknown. recent literature also describes other venues of complement activation: the “extrinsic protease” pathway, which involves direct cleavage of C3 and C5 by non-complement proteins such as kallikrein or thrombin [21] and the so-called “C2 bypass” pathway, entailing direct cleavage of C3 by MBL/MASP-2 [22].
A number of authors have emphasized the im-portance of LPS structure on the susceptibility of bacteria to serum bactericidal activity. These stud-ies were mainly focused on the role of sialic acid in this phenomenon [18, 23, 24]. Since mannan struc-tures are another component that is rarely found in the LPS of some bacterial strains, the authors of the present article deemed it worthwhile to inves-tigate how these structures affect the susceptibility of Gram-negative bacteria from the Enterobacte-riaceae family to serum bactericidal activity.
Material and Methods
Bacterial Strains
The study was carried out on 15 Enterobacte-riaceae strains that contain LPSs with mannan O-specific polysaccharides, which have been well de-scribed in world literature [2–5, 25–29] (Table 1). The bacterial strains were provided by the Polish Collection of Microorganisms (PCM) in Wroclaw and by the Veterinary research Institute in Pu-lawy (IW). The species affiliation of the examined strains was confirmed using the API-20E identifi-cation test kit (BioMérieux, Warsaw, Poland).
Serum
As described in a previous publication, normal human serum (NHS) was obtained from healthy volunteers untreated with any antimicrobial drug. The samples of NHS were collected, pooled and kept frozen at –70°C in 0.25 ml portions. A suit-able volume of serum was thawed immediately be-fore the experiment and used only once [18].
Determination of C3, C4, IgG
and IgM Levels in NHS
The levels of C3 and C4 complement com-ponents, IgG and IgM immunoglobulin were de-termined using specific antibodies. Nutrient agar plates with C3, C4, IgG and anti-IgM monospecific polyclonal antibodies (MEGA-TrADING, Gliwice, Poland) were used. The assay was carried out according to the instructions pro-vided by the supplier.
Treatment of Sera
The alternative pathway of complement acti-vation was blocked by incubating the NHS samples at 50°C for 20 min (HS50/20) [30]. The classical and lectin complement pathways were inhibited by using EGTA (SIGMA-ALDrICH, USA) and MgCl2 (HSMgEGTA). Complement activation by these pathways requires Ca2+ and hence may be inhibited by EGTA, which preferentially chelates Ca2+. The final concentration of EGTA and MgCl2 in the serum was 10 mM. EGTA solution was pre-pared according to Fine et al. [31]. NHS samples decomplemented by heating at 56°C for 30 min (HS56/30) were used as a control [19]. In HS56/30 serum all complement activation pathways were inhibited.
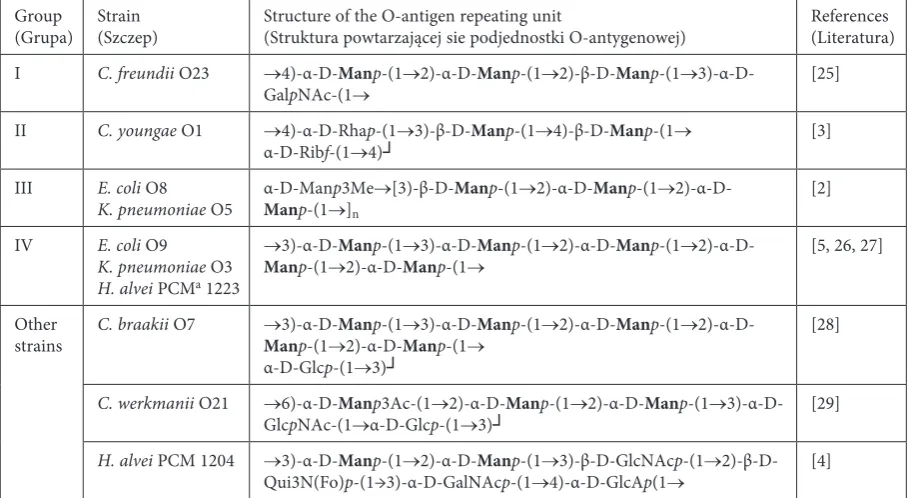
Table 1. The polysaccharide repeating units of O-antigenscomposed of mannan.amongrods of the Enterobacteriaceae family
Tabela 1. Wzory podjednostek polisacharydowych O-antygenów typu mannanowego w obrębie pałeczek z rodziny
Enterobacteriaceae
Group
(Grupa) Strain (Szczep) Structure of the O-antigen repeating unit (Struktura powtarzającej sie podjednostki O-antygenowej) references (Literatura) I C. freundii O23 →4)-α-D-Manp-(1→2)-α-D-Manp-(1→2)-β-D-Manp-(1→
3)-α-D-GalpNAc-(1→ [25]
II C. youngae O1 →4)-α-D-rhap-(1→3)-β-D-Manp-(1→4)-β-D-Manp-(1→
α-D-ribf-(1→4)┘ [3]
III E. coli O8
K. pneumoniae O5 α-D-ManManp-(1→p3Me]n →[3)-β-D-Manp-(1→2)-α-D-Manp-(1→
2)-α-D-[2]
IV E. coli O9
K. pneumoniae O3
H. alvei PCMa 1223
→3)-α-D-Manp-(1→3)-α-D-Manp-(1→2)-α-D-Manp-(1→
2)-α-D-Manp-(1→2)-α-D-Manp-(1→ [5, 26, 27]
Other
strains C. braakii O7 Man→3)-α-D-p-(1→Man2)-α-D-p-(1→Man3)-α-D-p-(1→Manp-(1→2)-α-D-Manp-(1→
2)-α-D-α-D-Glcp-(1→3)┘
[28]
C. werkmanii O21 →6)-α-D-Manp3Ac-(1→2)-α-D-Manp-(1→2)-α-D-Manp-(1→ 3)-α-D-GlcpNAc-(1→α-D-Glcp-(1→3)┘ [29]
H. alvei PCM 1204 →3)-α-D-Manp-(1→2)-α-D-Manp-(1→3)-β-D-GlcNAcp-(1→ 2)-β-D-Qui3N(Fo)p-(1→3)-α-D-GalNAcp-(1→4)-α-D-GlcAp(1→ [4]
Bactericidal Activity of NHS
The bactericidal activity of NHS was deter-mined as described by Cisowska et al. [18]. Briefly, the strains were grown overnight, and then bacte-rial cells were transferred to fresh nutrient broth (BIOMED, Warsaw, Poland) and incubated at 37°C for 30 min. After incubation the bacterial cells were centrifuged (4000 rpm for 20 min). Then the bac-terial suspensions were added to NHS, HS50/20, HSMgEGTA and HS56/30. To obtain various se-rum concentrations (25%, 50% and 75%), the NHS samples were diluted (v/v) with PBS. Then the bac-teria with sera were incubated in a water bath at 37°C. After 0, 60 and 180 min, samples were col-lected, diluted and cultured on nutrient agar plates (BIOMED, Warsaw, Poland) for 18 h at 37°C. The number of colony forming units (c.f.u.) at time 0 was taken as 100%. Strains that showed a sur-vival rate above 100% after 180 min of incubation in sera were regarded as resistant. The mean values from three separate experiments were calculated.Isolation and SDS-PAGE
Electrophoresis of LPS
The LPS of E. coli O8 strains (IW 728, IW 729, IW 928) were obtained using the phenol-water method described by Westphal and Jann [32] and analyzed by sodium dodecyl sulphate polyacryl-amide gel electrophoresis (SDS-PAGE) according to Laemmli [33]. Briefly, LPS samples were mixed with sample buffer containing 4% SDS and boiled for 5 min; 2.5 µl (1 µg) portions were applied to the gel. LPS bands were visualized after electrophore-sis by the silver staining method [34]. Smooth-type LPS from E. coli O111 strain (Sigma-Aldrich, Poznan, Poland) was used as a control reference in the process of electrophoretic separation [35].
Results
The levels of C3, C4, IgG and IgM in NHS were found to be: 1.15 mg/ml (standard: 0.9–1.8 mg/ml), 0.11 mg/ml (standard: 0.1–1.8 mg/ml), 16.3 mg/ml (standard: 7.0–16.0 mg/ml) and 1.77 mg/ml (stan-dard: 0.4–2.3 mg/ml), respectively [36].
The current study focused on bacterial strains for which the O-polysaccharide chain structures have already been determined and published (ap-propriate references are given in Table 1). An analysis of these structures showed that some of the studied strains possess the same formula of O-antigen repeating units. Therefore, the tested strains were divided into four groups (see Table 1).
Strains possessing other O-antigen subunit struc-tures were considered separately.
In the first part of experiments the suscepti-bility of all the studied strains to 50% NHS was determined. The results of these experiments are summarized in Table 2. The susceptibility to NHS among the examined strains was diverse: Eleven strains were sensitive to the bactericidal action of 50% NHS, and four were resistant. It is worth not-ing that among the four groups of strains only one was fully susceptible to the bactericidal action of 50% NHS, while in the three other groups both sensitive and resistant strains were found.
To more accurately characterize the suscepti-bility of the studied bacterial strains to NHS, ex-periments in which the bacteria were treated with 25% or 75% NHS were also performed. Strains sensitive to 50% NHS were incubated with 25% NHS, while the ones that were resistant to 50% NHS were treated with 75% NHS. The results of these experiments are shown in Tables 3 and 4, respectively.
As seen in Table 3, most of the strains that were sensitive to 50% NHS were also sensitive to 25% NHS. Only the strains belonging to the Group III became resistant to the reduced concentration of NHS.
The strains resistant to 50% NHS remained re-sistant to 75% NHS (Table 4). The exception was one of the strains belonging to Group III, which became susceptible to 75% NHS. Comparing Ta-bles 3 and 4 it is noticeable that the susceptibility to different NHS concentrations changed exclu-sively among the strains belonging to Group III.
Table 2. The bactericidal activity of 50% NHS against bacteria with O-antigens composed of mannan Tabela 2. Bakteriobójcza aktywność 50% NSL wobec bakterii z O-antygenami typu mannanowego
Group (Grupa) Strain (Szczep)
Table 3. Bactericidal activity of 25% NHS against bacteria with O-antigens composed of mannan Tabela 3. Bakteriobójcza aktywność 25% NSL wobec bakterii z O-antygenami typu mannanowego
Group (Grupa)
Strain (Szczep) Time of incubation [min] (Czas inkubacji [min]) 0 60 180
Susceptibility (Podatność)
c.f.u c.f.u. percent survival ± SD c.f.u. percent survival ± SD I C. freundii O23 PCM 1556 7.1·10 6 1.3·10 1 0.0002 ± 0.0001 7.0·10 0 0.0001 ± 0.0000 s PCM 2352 1.6·10 7 1.3·10 2 0.0008 ± 0.0003 1.0·10 1 0.0001 ± 0.0000 s II C. youngae O1 PCM 1492 8.8·10 6 2.3·10 3 0.03 ± 0.02 1.1·10 2 0.001 ± 0.01 s PCM 1493 5.4·10 6 2.6·10 2 0.005 ± 0.001 1.4·10 3 0.03 ± 0.01 s III E. coli O8 IW 728 1.3·10 7 3.1·10 6 23.8 ± 14.5 3.4·10 7 261.5 ± 7.8 r IW 729 6.2·10 6 1.0·10 7 161.3 ± 25.0 2.0·10 7 322.6 ± 33.7 r K. pneumoniae O5 PCM 57 3.5·10 7 6.9·10 6 19.7 ± 0.8 1.2·10 8 342.9 ± 78.7 r IV K. pneumoniae O3 PCM 11 1.5·10 7 5.2·10 6 34.7 ± 4.1 2.1·10 6 14.0 ± 2.3 s Other strains (Inne szczepy) H. alvei PCM 1204 1.4·10 7 3.8·10 3 0.03 ± 0.002 3.6·10 2 0.003 ± 0.001 s C. braakii O7 PCM 1532 8.3·10 6 1.6·10 6 19.2 ± 4.1 3.9·10 6 47.0 ± 16.0 s C. werkmanii O21 PCM 1554 1.5·10 7 1.9·10 6 12.7 ± 4.3 3.4·10 6 22.7 ± 4.2 s Table 4. Bactericidal activity of 75% NHS against bacteria with O-antigens composed of mannan Tabela 4. Bakteriobójcza aktywność 75% NSL wobec bakterii z O-antygenami typu mannanowego
Group (Grupa)
Strain (Szczep) Time of incubation [min] (Czas inkubacji [min]) 0 60 180
Susceptibility (Podatność)
Table 5. The mechanism of the NHS bactericidal activity against bacteria with O-antigens composed of mannan
Tabela 5. Mechanizm bakteriobójczej aktywności NSL wobec bakterii z O-antygenami typu mannanowego
Group
(Grupa) Strain (Szczep) Percent of surviving bacteria after 180 min of incubation (±SD) (Odsetek przeżywających bakterii po 180 min inkubacji (±SD)) HS50/20e (SL50/20) HSMgEGTAf (SLMgEGTA)
I C. freundii O23 PCM 1556 0.0001 ± 0.00003 s 0.01 ± 0.004 s PCM 2352 0.1 ± 0.03 s 0.2 ± 0.2 s II C. youngae O1 PCM 1492 0.006 ± 0.004 s 0.03 ± 0.01 s PCM 1493 1.1 ± 0.2 s 0.04 ± 0.02 s III E. coli O8 IW 728 1880.0 ± 521.2 r 0.7 ± 0.2 s IW 729 1041.7 ± 187.6 r 1.2 ± 0.9 s
K. pneumoniae O5 PCM 57 814.8 ± 210.8 r 39.3 ± 9.2 s IV K. pneumoniae O3 PCM 11 5.5 ± 2.0 s 10.8 ± 3.5 s Other
strains (Inne szczepy)
H. alvei PCM 1204 2.0 ± 1.1 s 0.01 ± 0.001 s
C. braakii O7 PCM 1532 0.4 ± 0.1 s 0.4 ± 0.01 s
C. werkmanii O21 PCM 1554 2.6 ± 1.0 s 0.4 ± 0.02 s
e HS50/20 – human serum with inactivated alternative complement pathway. eSL50/20 – ludzka surowica z inaktywowaną alternatywną drogą układu dopełniacza. fHSMgEGTA – human serum with inactivated classical and lectin complement pathways. fSLMgEGTA – ludzka surowica z inaktywowaną klasyczną i lektynową drogą układu dopełniacza
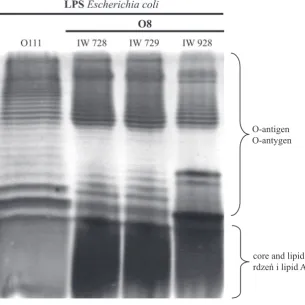
Fig. 1. Silver-stained SDS–PAGE of the LPS from E. coli O8 strains
Ryc. 1. Obraz elektroforetyczny LPS ze szczepów E. coli O8
O-antigen O-antygen
Heat inactivation of NHS (HS56/30) com-pletely abolished its bactericidal activity against all examined strains. The percent of surviving bacte-rial cells, depending on the strain, ranged from 1111.1% to 10181.8%.
The data concerning the susceptibility of bac-teria to NHS and the mechanism of the bacteri-cidal activity of NHS showed that the strains in Group III significantly differ from the others. This inspired the authors to isolate the LPSs from three E. coli O8 strains in Group III. The LPSs were pu-rified and subjected to SDS-PAGE. The electro-phoretic separation of LPS, when observed by sil-ver staining, exhibited a profile with characteristic ladder-like bands, which unambiguously indicates the presence of the O-antigen part of the LPS. As shown in Figure 1, all the tested E. coli O8 strains (IW 728, IW 729 and IW 928), as well as the con-trol E. coli O111 strain [35], possess such ladder-like bands and, consequentl,y smooth-type LPSs.
Discussion
The impact of LPS on the susceptibility of bac-teria to the bactericidal activity of serum has been investigated by many scientists [17–19, 23]. These studies include, among other things, analyses of the length of LPS [37] and its composition [23, 24], but up to now there have been no reported analyses of the role of mannan in the susceptibil-ity of bacterial strains to the bactericidal activsusceptibil-ity of serum. Therefore, the aim of the present study was to check and elucidate the susceptibility of bacte-rial strains with the same O-antigen repeating unit structures to NHS bactericidal activity. The au-thors also decided to study and compare which of the complement activation pathways plays a key role in serum bactericidal activity against these strains.
Considering the results of the tests with 50% NHS, it is not possible to state that there is a close correlation between the structure of O-antigen mannan repeating units and the tested strains’ susceptibility to NHS. As shown in Table 2, differ-ent strains with the same type of O-antigen varied in their susceptibility to 50% NHS. In three stud-ied groups of bacteria both sensitive and resistant strains were found.
Looking only at the results obtained for 50% NHS, one might draw the (not fully reasonable) conclusion, that the structure of O-antigen mannan does not affect bacterial susceptibility to NHS at all. The reliability of such a hypothesis can be refuted, however, when one considers the results obtained for NHS concentrations other than 50%t. In par-ticular, a comparison of the behavior of the strains
in Group III with other groups suggests that some kind of association might be found between the O-antigen mannan structure and the strains’ sus-ceptibility to NHS. With the exception of the Group III, the strains studied kept the same susceptibil-ity pattern to 25% and 75% serum concentrations (Tables 3 and 4). Moreover, in additional experi-ments (data not shown), the strains in Groups I, II and IV also showed the same susceptibility pat-tern when analyzed with 12.5% (those susceptible to 25% NHS) and 87.5% (those resistant to 75% NHS) concentrations of NHS. Only the strains in Group III “changed” their behavior when the NHS concentration was altered: The strains that were sensitive to 50% NHS were resistant to a 25% con-centration, while the resistant strains at 50% were sensitive when exposed to a 75% serum concentra-tion. These results were unlike the results observed in all of the other groups of bacteria tested. As discussed below, the exceptional behavior of the strains in Group III seems to be confirmed by the results of the experiments aimed at elucidating the complement activation pathways.
The discovery of the fact that bacterial strains with the same O-antigen differ in their susceptibil-ity to various NHS concentrations led the authors to investigate which of the complement activation pathways plays a key role in serum bactericidal ac-tivity against the tested strains.
The biological significance of mannan as a component of the O-specific chain is relatively well documented. Its importance in the activation of the immune response has been demonstrated by many scientific scientists [8, 9, 37, 38]. However, its role in the resistance of bacteria to serum bacte-ricidal activity has not yet been well described.
It is known that E. coli O8 LPS has a linear mannose homopolysaccharide chain and that the chemical structure of its O-specific chain are identical to that of K. pneumoniae O5. The K. pneumoniae O3 LPS and LPSs from E. coli O9 and H. alvei PCM 1223 strains also possess man-nose homopolysaccharide [2, 5, 26, 27], as shown in Table 1. It could therefore be assumed that the main mechanism for serum bactericidal activity against these strains is based on activation of the lectin pathway, which is initiated by the binding of MBL to carbohydrates, e.g. mannose, on microbial surfaces.
strains to serum activity. For these strains, all pathways of complement activation are probably equivalent. They were activated independently of each other, and this activation was sufficient to mediate bacterial lysis.
As shown in studies carried out on bacteria with mannose-rich O-antigens [9], LPSs possess-ing the mannose homopolysaccharide as an O-specific chain (K. pneumoniae O3, K. pneumoniae O5, E. coli O8, and E. coli O9) strongly activated the complement system, and there was no notice-able difference in the intensity of the anticomple-mentary activity among these LPSs. However, LPSs possessing heteropolysaccharide moiety showed much lower activity. The ability of LPS from K. pneumoniae O3 to activate human complement was more than 100 times higher than that of LPS from E. coli O111, E. coli O55 or Salmonella enter-itidis, strains without mannose in the O-antigens. Moreover, Yokochi et al. [9] established that K. pneumoniae O3 LPS probably activates the com-plement system by using the alternative pathway. For the strains in Group III, inactivation of the alternative complement pathway (by using HS50/20 serum preparation) led to these strains becoming resistant to serum. This result clearly demonstrates the crucial role of the alternative route in the mechanism of serum bactericidal ac-tivity against strains in Group III. This observation seems to be important, since these strains possess mannose-containing homopolysaccharides as the O-antigens.
In contrast to the results presented above, Ji-ang et al. [38] showed that LPSs possessing a man-nose-rich polysaccharide structure (isolated from K. pneumoniae O3, K. pneumoniae O5, E. coli O8, and E. coli O9 strains) activate the complement sys-tem via the lectin pathway. Other reports, however, emphasize the role of the alternative pathway or demonstrate the equal role of all three pathways of complement activation (ie, the classical, lectin and alternative pathways). Schweinle et al. [8] showed that MBL enhances complement deposition via the alternative pathway on Salmonella montevideo
strains possessing a mannose-rich LPS and results in serum killing these organisms, which are resis-tant to complement lysis in the absence of MBL. They emphasized that MBL and the complement system cooperate in the host organism’s first line of defense. On the other hand, a study carried out by Fernandez-Prada et al. [39] showed that both the classical and lectin complement pathways are involved in killing Brucella abortus and Brucella melitensis. This result is interesting, because these strains do not possess mannose in the O-antigens.
Thus, in light of the results of the current study and those obtained in other laboratories, one can conclude that the presence of the mannose O-antigen homopolymer is not a sufficient factor to determine lectin complement pathway activation.
The differences in susceptibility to NHS and complement pathway activation among the E. coli O8 rods in Group III led the current authors to isolate the LPSs from these strains. Contrary to expectations, all strains contained smooth-type LPS, regardless of whether they were derived from serum-sensitive or serum-resistant strains. This means that although the presence of mannan structures does not directly correlate with the sus-ceptibility of bacteria to NHS, mannan O-antigen structures can in some cases affect the mechanism of serum bactericidal activity against certain bacte-rial strains.
It should be noted that mannan is a substance rarely found in structures of Gram-negative bacte-ria. Therefore, the small number of strains and the wide diversity of species used in this study does not allow clear conclusions to be formed regarding the importance of mannan in protecting bacteria against the host’s defense mechanisms. It should be also borne in mind that the bactericidal activity of serum is a very complex phenomenon, and still not fully understood. Many components of the bacterial membrane are involved in this process, but LPS plays the most important role. Therefore, it seems advisable carry out further studies to check how mannan-containing LPSs isolated from the E. coli O8 strains in Group III affect this activity.
References
[1] Raetz CRH: Biochemistry of endotoxin. Annu rev Biochem 1990, 59, 129–170.
[2] Jansson P, Lonngren J, Widmalm G: Structural studies of the O-antigen polysaccharides of Klebsiella pneumoniae O5 and Escherichia coli O8. Carbohydr res 1985, 145, 59–66.
[3] Kocharova NA, Mieszała M, Zatonsky GV, Staniszewska M, Shashkov AS, Gamian A, Knirel YA: Structure of the O-specific polysaccharide of Citrobacter O1 containing an α-D-ribofuranosyl group. Carbohydr res 2004, 339, 321–325.
[4] Katzenellenbogen E, Romanowska E, Kocharova NA, Shashkov AS, Knirel YA, Kochetkov NK: Structure of the polysaccharide of Hafnia alvei 1204 containing 3,6-dideoxy-3-formamido-D-glucose. Carbohydr res 1995, 273, 187–195.
[6] Nelson RD, Shibata N, Podzorski RP, Herron MJ: Candida mannan: chemistry, suppression of cell-mediated immunity, and possible mechanisms of action. Clin Microbiol rev 1991, 4, 1–19.
[7] Dao DN, Kremer L, Guerardel Y, Molano A, Jacobs WR, Porcelli SA, Briken V:Mycobacterium tuberculosis lipomannan induces apoptosis and interleukin-12 production in macrophages. Infect Immun 2004, 72, 2067– 2074.
[8] Schweinle JE, Ezekowitz RA, Tenner AJ, Kuhlman M, Joiner KA: Human mannose-binding protein activates the alternative complement pathway and enhances serum bactericidal activity on a mannose-rich isolate of Sal-monella. J Clin Invest 1989, 84, 1821–1829.
[9] Yokochi T, Inoue Y, Kimura Y, Kato N: Strong interaction of lipopolysaccharides possessing the mannose ho-mopolysaccharides with complement and its relation to adjuvant action. J Immunol 1990, 144, 3106–3110.
[10] Podschun R, Ullmann U: Klebsiella spp. as nosocomial pathogens: epidemiology, taxonomy, typing methods, and pathogenicity factors. Clin Microbiol rev 1998, 11, 589–603.
[11] Ramos A, Damaso D: Extraintestinal infection due to Hafnia alvei. Eur J Clin Microbiol Infect Dis 2000, 19, 708–710.
[12] Gupta N, Yadav A, Choudhary U, Arora D: Citrobacter bacteremia in a tertiary care hospital. Scand J Infect Dis 2003, 35, 765–768.
[13] Farmer JJ. III. Enterobacteriaceae: introduction and identification. In: Murray Pr, Baron EJ, Pfaller MA, Tenover FC, Yolken rH. Eds. Manual of clinical microbiology. Washington D.C.: American Society for Microbiology 1999, pp. 442–458.
[14] Castro B, Montesinos I, Fuster-Jorge P, Delgado T, Miguel-Gómez MA, Sierra A: Epidemiology of Enterobac-teriaceae causing bloodstream infections in neonatal intensive care unit patients. Enferm Infecc Microbiol Clin 2010, 28, 227–232.
[15] Empel J, Baraniak A, Literacka E, Mrówka A, Fiett J, Sadowy E, Hryniewicz W, Gniadkowski M; Beta-PL Study Group. Molecular survey of beta-lactamases conferring resistance to newer beta-lactams in Enterobacteria-ceae isolates from Polish hospitals. Antimicrob Agents Chemother 2008, 52, 2449–2454.
[16] Frank MM: Complement deficiencies. Pediatr Clin North Am 2000, 47, 1339–1354.
[17] Kłak M, Jankowski S: The influence of lipopolysaccharides isolated from Enterobacteriaceae strains on the bacte-ricidal activity of normal cord serum. Adv Clin Exp Med 2010, 19, 57–64.
[18] Cisowska A, Bugla-Płoskońska G, Tichaczek-Goska D, Doroszkiewicz W, Jankowski S: The susceptibility of Escherichia colistrains with sialic acid-containing lipopolysaccharides or capsules to the bactericidal action of normal human serum. 7th conference: Molecular biology in diagnostics of infectious diseases and biotechnology.
Warsaw: SGGW, 2004, 41–47.
[19] Jankowski S, Rowiński S, Cisowska A, Gamian A: The sensitivity of Hafnia alveistrains to the bactericidal effect of serum. FEMS Immunol Med Microbiol 1996, 13, 59–64.
[20] Fujita T, Matsushita M, Endo Y: The lectin-complement pathway – its role in innate immunity and evolution. Immunol rev 2004, 198, 185–202.
[21] Markiewski MM, Lambris JD: The role of complement in inflammatory diseases from behind the scenes into the spotlight. Am J Pathol 2007, 171, 715–727.
[22] Atkinson JP, Frank MM: Bypassing complement: evolutionary lessons and future implications. J Clin Invest 2006, 116, 1215–1218.
[23] Bugla-Płoskońska G, Doroszkiewicz W: Bactericidal activity of normal bovine serum (NBS) directed against some Enterobacteriaceaewith sialic acid-containing lipopolysaccharides (LPS) as a component of cell wall. Pol J Microbiol 2006, 55, 169–174.
[24] Mielnik G, Gamian A, Doroszkiewicz W: Bactericidal activity of normal cord serum (NCS) against Gram-negative rods with sialic acid-containing lipopolysaccharides (LPS). FEMS Immunol Med Microbiol 2001, 31, 169–173.
[25] Katzenellenbogen E, Ekiel I, Romanowska E: The structure of the O-specific polysaccharide chain from Citro-bacterO23-lipopolysaccharide. Carbohydr res 1988, 179, 349–357.
[26] Curvall M, Lindberg B, Lonngren J, Nimmich W: Structural studies on the KlebsiellaO group 3 lipopolysaccha-ride. Acta Chem Scand 1973, 27, 2645–2649.
[27] Katzenellenbogen E, Kocharova NA, Zatonsky G.V Kübler-Kiełb J, Gamian A, Shashkov AS, Knirel YA, Ro-manowska E: Structural and serological studies on Hafnia alveiO-specific polysaccharide of alpha-D-mannan type isolated from the lipopolysaccharide of strain PCM 1223. FEMS Immunol Med Microbiol 2001, 30, 223– 227.
[28] Kocharova NA, Zatonsky GV, Bystrova OV Shashkov AS, Knirel YA, Kholodkova EV, Stanislavsky ES: Struc-ture of the O-specific polysaccharide of Citrobacter braakiiO7a, 3b, 1c. Carbohydr res 2001, 333, 335–338.
[29] Knirel YA, Kocharova NA, Bystrova OV, Katzenellenbogen E, Gamian A: Structures and serology of the O-specific polysaccharides of bacteria of the genus Citrobacter. Arch Immunol Ther Exp 2002, 50, 379–391.
[30] Eidinger D, Bello E, Mates A: The heterocytotoxicity of human serum. I. Activation of the alternative comple-ment pathway by heterologous target cells. Cell Immunol 1977, 29, 174–186.
[31] Fine DP, Marney SR, Colley DG, Sergent JS, Des Prez RM: C3 shunt activation in human serum chelated with EGTA. J Immunol 1972, 109, 807–809.
[32] Westphal O, Jann K: Bacterial lipopolysaccharides: extraction with phenol-water and further application of the procedure. Meth Carbohydr Chem 1965, 5, 83–91.
[34] Tsai CM, Frasch CE: A sensitive silver stain for detecting lipopolysaccharides in polyacrylamide gels. Anal Bio-chem 1982, 119, 115–119.
[35] Gupta RK, Egan W, Bryla DA, Robbins JB, Szu SC: Comparative immunogenicity of conjugates composed of Escherichia coliO111 O-specific polysaccharide, prepared by treatment with acetic acid or hydrazine, bound to tetanus toxoid by two synthetic schemes. Infect Immun 1995, 63, 2805–2810.
[36] Herold G: Medycyna wewnętrzna 4th ed. Warsaw: PZWL, 2006, pp. 1131–1142.
[37] Murray GL, Attridge SR, Morona R: Altering the length of the lipopolysaccharide O-antigen has an impact on the interaction of Salmonella entericaserovar Typhimurium with macrophages and complement. J Bacteriol 2006, 188, 2735–2739.
[38] Jiang G, Sugiyama T, Kato Y, Koide N, Yokochi T: Binding of mannose-binding protein to Klebsiella O3 lipo-polysaccharide possessing the mannose homolipo-polysaccharide as the O-specific lipo-polysaccharide and its relation to complement activation. Infect Immun 1995, 63, 2537–2540.
[39] Fernandez-Prada CM, Nikolich M, Vemulapalli R Sriranganathan N, Boyle SM, Schurig GG, Hadfield TL, Hoover DL: Deletion of wboAenhances activation of the lectin pathway of complement in Brucella abortusand Brucella melitensis.Infect Immun 2001, 69, 4407–4410.
Address for correspondence:
Dorota Tichaczek-GoskaDepartment of Biology and Medical Parasitology Wroclaw Medical University
Mikulicza-radeckiego 9 50-367 Wrocław Poland
Tel.: +48 71 784 1523
Email: [email protected]