Copyright 0 1988 by the Genetics Society of America
Is
the
Suppressor-mutator
Element
Controlled
by a
Basic Developmental
Regulatory Mechanism?
N. V.
Fedoroff and
J.
A. Banks
Department of Embryology, Carnegie Institution of Washington, Baltimore, Maryland 21210 Manuscript received March 1 1, 1988
Accepted June 8, 1988
ABSTRACT
We report the results of genetic studies on derivatives of two different alleles of the maize a locus
with an insertion of the Suppressor-mutator (Sprn) transposable element in which the element is inactive, but can be reactivated readily. We present evidence that the mechanism that determines whether the element is in an active or inactive phase has two genetically distinguishable components. One determines whether or not the element is genetically active (the phase setting) and the other determines the stability of the setting in development, its heritability, and its phase in the next generation (the phase program). We show that the element’s phase can be reset in a reproducible pattern during plant development. We also show that the Spm element can be reprogrammed to undergo a subsequent phase change without a concomitant phase change. The capacity to reset and reprogram the Sprn element is differentially expressed within the plant in a pattern that is correlated with the develop- mental fate of apical and lateral meristems, suggesting the involvement of a basic developmental
determination mechanism.
I
N studies on the maize Suppressor-mutator (Sprn) transposable element, MCCLINTOCK (1 957-1 959, 1961a, b, 1962, 1965, 1967, 1971) discovered the existence of a genetic mechanism that determines the pattern of element expression in the maize plant. T h e mechanism is both heritable and reversible. It displays a startling versatility in the types of spatial and tem- poral patterns of element expression that it can estab- lish and heritably maintain. T h e Spm element can exist in one of three states. T h e first of these is a constitutively active state in which the element is expressed throughout development. T h e second is a cryptic state in which the element is genetically un- detectable and its presence can be inferred only from extremely rare incidents of spontaneous or induced activation (NEUFFER 1966; FEDOROFF 1986; FEDOROFFet al. 1988). In the third state, the element is pro- grammed to be expressed in any one of many different temporal or spatial patterns during the development of the plant (MCCLINTOCK 1959, 1961a, b, 1965,
197 1 ; PETERSON 1966; FOWLER and PETERSON 1978). T h e element’s program can specify that it is either in the active or inactive phase at the beginning of the plant’s developmental cycle and that it will later undergo changes in the phase of activity at specific times in development, at regular intervals, in certain plant parts or in certain areas of a given tissue (Mc-
CLINTOCK 1959; PETERSON 1966). MCCLINTOCK
(1 958, 1959, 197 1) described elements, for example, that were active early in development, but cycled between the active and inactive phases late in devel- opment, as well as others that remained inactive dur-
Genetics 1 4 0 559-577 (October, 1988)
ing most of the plant’s life cycle, returning to an active form infrequently and late in development. PETERSON (1 966) described a form of the cognate Enhancer ( E n ) element that was active only in the crown of the kernel or, conversely, active throughout the kernel, but not in the crown.
MCCLINTOCK (1959) noted that when an element underwent a change in its phase of activity during plant development, it could exhibit a heritable change in its pattern of expression. She concluded that when an element changed its phase of activity, it could also be reprogrammed, heritably altering the duration or spatial pattern of its subsequent expression. In the course of these studies, MCCLINTOCK further ob- served that the patterns of element expression were influenced by interactions between different Spm ele- ments in the genome (MCCLINTOCK 1958, 1971). When she combined an element undergoing frequent changes of activity phase with an element in a pro- longed inactive phase in the same plant, she found that the inactive element exhibited the genetic behav- ior of an active element. She further reported that the active and inactive elements segregate unchanged in progeny (MCCLINTOCK 1959).
560 N. V. Fedoroff and J
a.
A. Banks
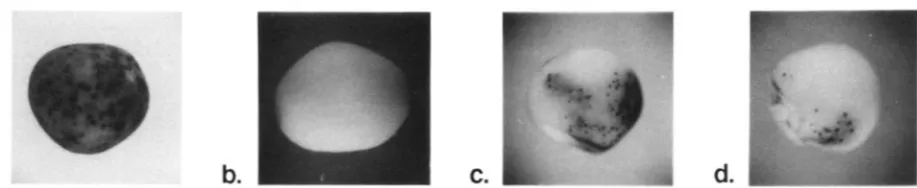
FIGURE 1 .-Phenotypes of kernels having thc genetic constitution u-m2-7991;\1 S h / a sh2 w i t h a , a fully active S p m ; I,. a t 1 in;tctivc Spm; c
and d , an inactive Spm returning to the active phase during kernel development.
by using probes homologous to the insertion site of the element under study. We chose the a locus of maize both because it has been cloned and because there are Spm insertions at different sites within the locus that give alleles with markedly different genetic properties (MCCLINTOCK, 196 1 a, b, 1962; O’REILLY et al. 1985; MASSON et al. 1987).
We used two different alleles of the a locus with Sprn insertion mutations in an effort to identify factors common to the inactivation mechanism. One was a derivative of the original a-m2 allele, designated a-m2- 7991Al (MCCLINTOCK 195 1, 1962; MASSON et al. 1987). It was chosen because it permits changes in element activity to be monitored not only by excision of the Sprn element, but also by changes in expression of the adjacent a gene, which is expressed under the control of the element in these alleles. A colorless derivative of MCCLINTOCK’S (1 96 1 b, 1962) original a- m5 allele was chosen because it is useful in monitoring changes in expression of the element’s truns-acting functions without interference from phenotypes re- sulting from element-promoted changes at the a locus. In the present communication, we describe the ge- netic properties of derivatives of both selected Spm insertion alleles in which the resident element is in- active, but can be reactivated readily. We present evidence that the mechanism that controls element activity during development has two components. One component determines whether or not the ele- ment is expressed (the phase setting) and the other determines the stability of the setting in development, its heritability, and its phase in the next generation (the phase program). We report that elements can be reset, undergoing reproducible, lineage-specific changes
in the phase of activity. We also show that they can be reprogrammed without being reset, altering the heritability of the setting. Thus, the maize plant can program the Sprn element in a regular, directed way. T h e programming mechanism is correlated with the developmental determination of the apical and lateral meristems, suggesting the involvement of a basic de- velopmental determination mechanism. We discuss the implications of the present observations, as well as the growing evidence that they reflect reversible mod- ifications of specific DNA sequences, for the mecha-
nisms that control Spm element activity in particular and the developmental determination of gene expres- sion in general.
MATERIALS A N D METHODS
Spm-s alleles of the a locus: The alleles with Spm-s
insertions used in the present study are the a-m2-7991AI
allele (MASON et al. 1987) and a derivative of McClintock’s ( 1 95 1, 196 1 b, 1962) a-m5 allele. Both have a standard, fully functional Sprn (Spm-s) element inserted at the a locus. The
a-m2-799IA I allele belongs to the a-m2 series of alleles that we have designated Spm-dependent because the a gene is co- expressed with the element (MASSON et al. 1987). lts phe- notype is shown in Figure la. The a-m2-7991AI allele ex- hibits background pigmentation due to co-expression of the a gene and Spm element, a moderate somatic excision fre- quency and a high germinal excision frequency (MASSON et al. 1987). The a-m2-7991AI allele has an 8.3-kb Spm-s
element inserted 0.1 kb upstream of the a gene’s transcrip tion start site (SCHWARZ-SOMMER et al. 1987; MASON et al.
1987).
The original a-m5 allele isolated by MCCLINTOCK ( 1 962) was highly unstable. I t had a colorless phenotype and a high somatic and germinal mutation frequency to full a gene expression, giving a deeply pigmented phenotype. We re- ceived two derivatives of the original a-m5 allele from MCCLINTOCK and have used both in the present study. The first of these is designated a-m5c and exhibits the extreme instability of the Spm insertion characteristic of the original
a-m5 allele, but has a stably colorless phenotype by virtue of a second mutation that the a locus acquired either de novo or by recombination with another null allele (B. MG CLINTOCK, personal communication). The a-m5c allele was chosen for the present study because its null phenotype permits the monitoring of the element’s trans-acting func- tions without interference from phenotypes associated with the excision of the element itself. The second, designated a- m5w, is described below.
Spm Control in Development 56 1
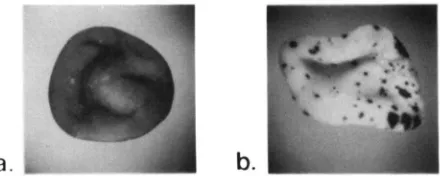
FIGURE 2.--Phenotvpes of kernels having thc gcnctic constitu- tion a-ml-5719AI Sh2 either a, without an Spm-s or b , with an
Spm-s.
element (Figure 2). The wx-m8 allele has a null phenotype and its dSpm insertion excises at a high frequency in the presence of an Sprn element (MCCLINTOCK 1961a). The a and wx genes are both expressed in endosperm and can be monitored separately. This makes it possible to distinguish genetic changes that affect the trans-acting Sprn from those that affect either dSpm element, since changes in the activity of the trans-acting element affect both dSpm elements simi- larly.
A stable recessive mutant allele and a wild-type allele of the nearby shrunken-2 (sh2) locus were used as segregation markers for the a locus because of their close linkage (0.1 - 0.2% recombination) and the distinctive endosperm phe- notypes associated with the Sh2 and sh2 alleles (COE and NEUFFER 1977). The Spm insertions were both linked to the
Sh2 allele, which gives full, rounded kernels (Figure 1). The
a-ml-5719AI and other tester alleles were linked to the sh2
allele (Figures 2 and 3), which gives shrunken endosperm. These allelic combinations made it possible to distinguish readily between Sprn elements that had undergone inacti- vation at the original insertion site from those that had become inactive concomitant with transposition away from the original insertion site at the a locus.
Trans-activation of an inactive Sprn element: The pres- ence of an inactive Spm was routinely verified by the ability of an active Spm to trans-activate an inactive element. Three different Spm elements with reduced activity, designated
Spm-w, were employed for this purpose. Two of the three are Spm-w elements derived from the original a-m2 (a-m2- 8 0 1 1 ) and a-m5 (a-m5w) alleles (MCCLINTOCK 1962, 1963). The a-m2-8011 allele has been cloned and shown to have a
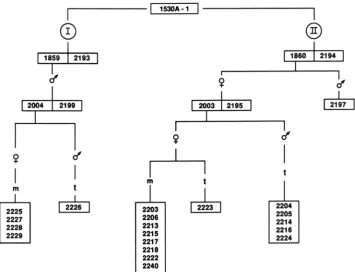
1.6-kb internal deletion in the Sprn element and to support the synthesis of IO-20% as much of the element's major transcript as an Spm-s element (MASSON et al. 1987). The a- m5w allele contains a structurally intact Spm element at a site within the a gene and its molecular defect is not known. Both alleles exhibit a sparse variegation pattern (Fig. 3, a and c) that becomes dense as a consequence of increased somatic reversion in the presence of an Spm-s element (Fig. 3, b and d). A third strain with an uncharacterized Spm-w
element linked to the p r locus on chromosome 5 , but not the a locus (chromosome 3 ) , was used to verify that trans- activation of an inactive element was independent of the location of the trans-activating element.
Screening for Spm-i at the a locus: T o identify inactive derivatives of the elements at the a locus, crosses were made in such a way that the activity of the single element could be followed and its location assessed readily in subsequent crosses. T h i s was done by crossing plants with the genetic constitution a-m2-7991Al Sh2/a sh2 or a-mI-5719A1 sh2, wx-m8 or wx/wx-m8 or that were homozygous for the a-m5c, Sh2 and wx-m8 alleles, to and by plants that were homozy- gous for the two dSpm tester alleles, a-ml-5719Al and wx- m8. The use of a dSpm tester favored recovery of inactive
rather than transposed elements. This is because it permits the detection of an active Spm element at any chromosomal location by its ability to excise the dSpm element of the tester allele and can therefore detect elements that have trans- posed away from the a locus.
Kernels that differed from the variegated phenotype as- sociated with the particular marker combination in one of two ways were selected for further analysis. The selected kernels either showed a marked reduction in Sprn activity throughout the kernel endosperm or exhibited Spm activity only in endosperm sectors. For the a-m2-7991AI allele, this was judged by the less frequent or sectorial excision of the resident element at the a locus (Figure 1). For the a-m5c
allele, this was judged by reduced suppression of the a-ml- 5719Al allele (Figure 2) and by infrequent excision of the
dSpm elements of the a-ml-5719AI and wx-m8 alleles. Ker- nels showing no evidence of transposable element activity were avoided in order to minimize the recovery of stable a
gene derivatives from which the element had excised. Plants grown from kernels showing reduced or sectorial transpos- able element activity were backcrossed to and by tester plants that were homozygous for the a-ml-5719A1, sh2, and
wx-m8 alleles. Ears were scored for the presence or absence of transposable element activity, as well as whether the transposable element activity co-segregated with the Sh2
allele, as it had in the parental plants. Ears showing evidence of an inactive element at the a locus returning to an active form were selected for further analysis.
Analysis of Spm-i derivatives: The progeny of two plants, one carrying the a-m2-7991A1 allele and the other carrying the a-m5c allele, and each showing evidence of element inactivation were chosen for further analysis. Of these, the derivatives of the a-m2-7991A1 allele have been subjected to the more extensive analysis. Nonetheless, enough crosses have been done with the inactive derivative of the a-m5c allele to verify that the inheritance patterns are similar for the two derivatives.
The bulk of the analysis has been done on two lineages derived from plant 1530A-1 (a-m2-7991A1 ShZla-rnl- 571 9A 1 sh2, wx-m8), which produced two ears, both of which were crossed by a-ml-5719Al sh2, wx-m8 tester plants (Fig- ure 4). Two plants, designated 1859A-1 and 1860-1, were grown in the summer of 1985 from kernels exhibiting reduced Spm expression, selected from ear I and ear 11 of plant 1530A-1. Additional kernels with a fully active Spm
from the same ears were grown during the summer of 1987 and comprise cultures 2193 and 2194. Kernels with an inactive Spm phenotype selected from the ear produced by plant 1860-1 were grown in the summer of 1986 (family 2003) and 1987 (family 2 195). Inactive Spm kernels selected from an ear pollinated with pollen collected from plant 1860-1 were grown in the summer of 1987 (family 2197). Kernels with an inactive or slightly active Sprn element selected from an ear on a tester plant crossed by plant 1859A-1 were grown in the summer of 1987 and designated families 2 199A and B, respectively, and a sibling family was grown from kernels with an inactive element in the summer of 1986 (family 2004). Kernels with either inactive or active
Spm phenotypes were selected from ears obtained in families 2003 and 2004 and grown as families 2203-2229 in the summer of 1987. I t should be noted that pollen was obtained only from tiller tassels in families 2003 and 2004 (Figure 4) because of prevailing weather conditions (heat and drought). The lineage with an inactive Sprn derivative of the a-m5c
562 N. V. Fedoroff and J. A. Banks
FIGURE 3,"Phenotypes of kernels having the genetic constitutions a, a-m5w sh2; b, a-m5w sh2/a-m2-7991AI Sh2; c, a-m2-801 I sh2; d, a- m2-8011 sh2/a-m2-7991AI Sh2. The Spm elements of the a-rn2-7991AI allele were inactive in the parent plant from which the illustrative kernels were taken. Their variegation pattern is attributable to trans-activation of the inactive Spm-s element of the a-m2-7991AI allele by the Spm-w element at the a locus of the a-m5w and a-m2-801 I alleles. The high density of the variegation pattern is due to the much higher frequency at which the elements residing in the Spm-w alleles excise to give the A phenotype in the presence of an Spm-s when compared with the a-m2-7991Al allele (Figure la).
I
I
0
I
I
rn t
2217
2222 2240
t
t
$J
FIGURE 4.-Relationships among the plant families with an inactive Spm element arising from the a-m2-7991AI allele. All of the kernels on both ears I and I1 of plant 1530A-1 had an active Spm element, although the two kernels initially selected and grown, giving plants 1859A-1 and 1860-1. showed evidence of reduced element activity. The sibling families 2193 and 2194 were grown from kernels exhibiting an Spm-s phenotype. All subsequent families, with the exception of families 2199B and 2240, were grown from kernels in which the Spm
element was inactive. Family 2199B comprised plants grown from kernels in which the element was almost completely inactive, while family 2240 consisted of plants grown from kernels in which the element was fully active. The figure indicates whether the line of descent was through the male or female gametes and whether the gametes were produced on the main stalk (m) or a tiller (t).
m8 tester in the summer of 1983. The kernel was grown (plant 1856A-1) in the summer of 1985 and crossed by and onto plants that had the genetic constitution A ShZ/a-ml- 5719A1 sh2, Wxlwx-m8. Among kernels that showed trans- posable element activity, many gave a mottled phenotype (poor suppression of the a-ml-5719Al allele) and one or more deeply pigmented and Wx sectors signaling transpos- able element activity. Kernels exhibiting such a mottled phenotype with reduced transposable element activity were grown in the summers of 1986 (family 1997) and 1987 (families 2280 and 2282). Kernels selected from an ear obtained by backcrossing a plant in family 1997 by an a-ml- 5719Al sh2, wx-m8 tester were grown in the summer of 1987 (family 2283).
563 Spm Control in Development
TABLE 1
Identification of plants with an inactive Sptn element at the a locus
Location and activity of Sprn Active Spma
Active Spm' Inactive Sprn Inactive Sprn not at a lo- Inactive Spm not at a lo- unknown lo- Source allele at a locus cus at a locus cus cation
a-m5cb 7 61 1 2 0
a-m2-7991Al' 3 3 2d 1 1
Some of the plants were used as pollen parents in crosses to the dSpm tester strains, as well as egg parents. In some of the progeny ears
Ninety-seven kernels selected from ten ears carrying a a-m5c allele gave 7 1 plants that were used to make one or more crosses to a dSpm
obtained from such crosses, marked sectorial inactivation of the Spm element was observed.
tester plant.
' Eleven kernels selected from 19 ears carrying the a-m2-7991AI allele gave ten plants that were used in crosses to dSpm tester plants. These plants were grown from two kernels selected from different ears of the same plant.
for a null allele of the a locus and the same alleles of the sh2 and wx loci (a sh2, wx-m8). One or more ears on each plant was also self-pollinated. Similar crosses were carried out with pollen collected from the tassel on the main stalk and with pollen collected from tiller tassels.
Ears were scored for the fraction of kernels showing transposable element activity in the Sh2 and sh2 kernel classes. The results are reported as a percentage of Sh2 kernels exhibiting the colorless phenotype expected for an inactive Spm in the a-m2-7991Al allele. The correspondence between aleurone and endosperm phenotypes was assessed by scoring the wx phenotype in some or all kernels, either visually (for Sh2 kernels) or by staining endosperm with I- KI solution.
Tests of the ability of an Spm element to reactivate the Spm-i element were carried out using one of the several Spm-w elements discussed above. Kernels produced on ears resulting from crosses in which one parent carried an Spm-i element and the other carried an Spm-w element were scored for the fraction of kernels exhibiting an Spm-s phenotype in both the aleurone ( a locus) and endosperm (wx locus). In two of the Spm-w testers, the Spm-w element was at the a locus and tightly linked to the sh2 allele (a-m2-8011 sh2 and a-m5w sh2), while in a third tester, it was linked to the pr locus. All testers were homozygous for the wx-m8 allele.
The activity of the a-rn5c allele was assessed in crosses by the a-ml-5719Al sh2, wx-m8 tester. Kernels with an inactive element have the palely pigmented phenotype of the tester allele. Ears were scored for the fraction of kernels that showed no Spm activity, Sprn activity in kernel sectors, and full Spm activity.
Statistical tests: T o evaluate differences between progeny ears receiving the Spm element through different gamete sources, paired comparisons were made between different progeny ears of individual plants. This was done by calcu- lating the difference in the percentage of progeny kernels with an inactive Spm element between different ears on the Spm-containing plant or between an ear on the plant and an ear on a tester plant receiving pollen from the plant. All comparisons were therefore made between ears that re- ceived the Spm element from the same parent plant. No effort was made to compare family means. This was because the number of ears obtained from crosses varied among the plants in a family and because probability plots showed that the data were not normally distributed around the mean values obtained for each gamete source. Thus, within-plant differences were calculated for the following categories: (1) first and second ears on the main stalk (Spm transmitted through the ear parent); (2) the first ear on the main stalk and the first tiller ear to be pollinated (Spm transmitted
through the ear parent) and (3) the first ear on the main stalk of the Spm-bearing plant and an ear on a tester receiv- ing pollen from the tassel on the main stalk of the Spm- bearing plant. Differences calculated for each category from the data in Tables 2-8 were subjected to Wilcoxon's signed- ranks test and found to be significant at or above the 99.5% confidence level.
RESULTS
Identifying an inactive Sprn element at the a locus: Active Spm elements can transpose premeiotically to new sites in the genome and it has been reported that changes in element activity can accompany transposi- tion of the element (PETERSON 1976a, b). The Spm-s
element transposes away from the a locus of both t h e
a-m2-7991Al a n d a-m5c alleles used here at a relatively high frequency, giving rise to stable alleles of t h e locus and independently segregating Sprn elements (Mc- CLINTOCK 1962, 1963). T h e initial crosses t o prepare material for screening were therefore structured to minimize the recovery of stable alleles and favor re- covery of inactive elements (see MATERIALS AND METH-
ODS). It was anticipated that two types of variants would be recovered from t h e initial screen. These are (1) stable derivatives of the two a alleles with a trans- posed, but inactive Sprn element elsewhere in the genome, and (2) derivatives with an inactive Spm
element at its original location. It was also anticipated that some fraction of the phenotypic variants would not be heritable. Only kernels showing reduced or sectorial element activity were selected t o avoid re- covering very stably inactivated and mutant elements, either at t h e a locus or elsewhere.
Plants grown from the selected kernels were ana- lyzed for the presence and location of the Sprn element
contained an active element and in a majority (79%), the element was either unlinked to the Sh2 locus or less tightly linked, indicating that the element had transposed (Table 1). Among the plants in which the
Spm was active, t h e sectorial phenotype for which t h e original kernel was selected was sometimes observed
564 N. V. Fedoroff and J. A. Banks
when the plant was used as a pollen parent (see Table 1, first footnote). T h e marked instability of the a-m5c allele is apparent in the high frequency at which progeny proved to have a transposed element.
A total of seven plants gave ears with the phenotypes anticipated for an inactive Sprn element that is undergoing relatively frequent reactivation (Table 1). In three plants, the reactivating element was not at the a locus and in a fourth plant, the reactivation frequency was too low to assess the location of the Spm reliably. Three plants gave evidence that the Spm element at the a locus was inactive and undergoing frequent reactivation (Table 1). One of the plants carried the a-m5c allele, and the other two, which were grown from kernels selected from different ears produced by the same plant, carried the a-m2-7991Al allele.
Detailed genetic analysis has been carried out for the a-m2-7991Al derivative and the results are pre- sented in the following sections. T h e behavior of the analogous a-m5c derivative has been analyzed in less, but sufficient detail to show that the inheritance pat- terns are similar for both inactive derivatives of the active Spm element.
Inactivation occurs during plant development: T h e two kernels that were initially selected from different ears of a plant that carried the a-m2-7991Al allele exhibited a small number of deeply pigmented revertant sectors (plant 1530A-1; Figure 4). Ears pro- duced on the plants grown from these kernels and on tester ears crossed by the plants transmitted an active Spm to some, but not all progeny kernels (Table 2A). In all three ears obtained from these crosses, varie- gation was observed only in the Sh2 class of kernels, indicating that the Spm element had not moved away from the a locus (not shown). In the ear that received pollen from plant 1859A-1, 90% of the Sh2 kernels had the variegated phenotype associated with an ac- tive Sprn, while only half of the Sh2 kernels on the ear produced by plant 1860-1 were variegated. Moreover, among kernels exhibiting variegation, many showed relatively small areas with deeply pigmented sectors and some had only one or two deeply pigmented spots. T h e results obtained when plant 1860-1 served as pollen parent were similar to those obtained when it served as egg parent (Table 2A).
T o determine whether genetic factors in the paren- tal plant play a role in the inactivation of the element, a number of sibling kernels showing no evidence of Spm inactivation were selected from each ear of the same plant that gave rise to plants 1859-1 and 1860-
1 of Table 1. Plants were grown from the kernels and subjected to the same test crosses (see MATERIALS AND METHODS). T h e results are shown in part B of Table 2. Of 12 plants grown from such kernels, 7 showed little o r no evidence of Spm inactivation and 5 gave ears on which the Sprn was inactive in a significant
TABLE 2
Percent kernels with an inactive Sprn in progeny of plant 1530A-1"
Ear o n plant: Outcrosses using pollen from:
Plant NO.^ Ear I Ear 11 Tiller Main stalk Tiller
Part A'
1859A-1 10.1
1860-1 46.5 52.8 Part Bd
2193B-1 3.4
2193C-1 1.3 1.3 7.0
2193C-2 1.2
2193C-3 0.0 0.0
2193C-4 85.6 84.1
2193C-6 77.3
2194-1 11.5 0.7 41.0 20.0
2194-2 27.4 19.6 75.7 71.0
2 194-3 0.0 52.3 7.1
2 194-4 2.2 9.7 0.0
2194-5 13.4 5.5 57.7
2 194-6 0.0 0.0 0.0
* Ears were examined for kernels exhibiting one or more deeply pigmented spots resulting from excision of the Spm element from the a locus of the a-m2-7YYlAI allele. Only completely colorless kernels were judged to have received an inactive Sprn that remained in an inactive phase throughout development. The percentage of Sh2 kernels receiving an inactive Sprn element is recorded in the table.
'
The number is that of the plant carrying the Spm element. All plants were intercrossed with the dSpm tester plants described in MATERIALS A N D METHODS. These had the genetic constitution a sh2, wx-m8 or a-ml-571YA1 sh2, wx-m8.'Plants 1859A-1 and 1860-1 were grown from kernels with reduced Sprn activity selected from ears I and 11, respectively, on plant 1530A-1.
Plants in part B of the table were grown from kernels taken from the same ears as those in part A, but showing no apparent reduction in the activity of the Sprn at the a locus. Family 2193 comprises siblings of plant 1859A-1 and family 2194 comprises siblings of plant 1860-1.
fraction of kernels (1 1.5-85.6%). Moreover, the frac- tion of progeny Sh2 kernels showing no transposable element activity was markedly higher when some of the plants were used as the pollen parent than when they served as egg parents (see family 2 194, plants 1, 2, 3 and 5).
These observations indicate that the tendency to- ward element inactivation is not confined to plants grown from kernels in which the element exhibits reduced activity. T h e extent of element inactivation, as judged by the fraction of progeny kernels with an inactive element, varies markedly from plant to plant. Moreover, in some plants the Spm element appears to be inactivated more frequently on transmission through the male than through female gametes (Table 2B).
Spm Control in Development 565
TABLE 3
Percent kernels with an inactive Spm transmitted through ear I of plant 1860-1"
Ear on plant: Outcrosses using pollen from:
Main
Plant No. Ear I Ear 11 Tiller 1 Tiller 2 stalk Tillers
2195-1 96.1 100.0 65.7 100.0 95.7 96.2
2 195-2 92.9 0 95.8 85.8 1.6 17.5
2 195-3 49.4 51.9 98.6 99.1 70.5
2 195-4 96.6 80.4 95.1 98.0
2 195-5 99.5 86.4 97.1 54.9
2 195-6 95.9 98.1 24.1 98.3 96.6 69.4 2 195-7 99.6 98.3 97.1
2 195-8 97.4 97.8 28.9 93.5 87.7 74.4 94.5
2 195-9 100.0 99.3 98.9
2195-10 96.0 99.0 100.0 99.4
2195-11 80.2 59.8 92.3 16.7
2195-12 98.7 100.0 5.9 99.6
2195-13 82.3 67.7 86.1 1.1
2195-14 99.5 100.0 97.1 100.0 99.6
2195-15 100.0 100.0 99.2
2195-16 100.0 100.0 88.5 99.5
2195-17 100.0 97.3 98.3 92.3 2195-18 97.3 12.4 58.6 2195-19 100.0 88.6 97.0 78.5
2 195-20 100.0 99.6 100.0 98.9
2195-21 97.3 80.2 97.3
2 195-22 89.8 93.0 98.4 6.7
2 195-23 98.9 98.2 43.3 44.4 62.4
2 195-24 96.7 91.5 3.0
2 195-25 97.6 99.5 99.5 94.7
2 195-26 99.6 100.0 100.0 97.3
* The percentage of Sh2 kernels receiving an inactive Spm element is recorded in the table.
kernels had the same genotype (a-m2-7991AI Sh2/a- ml-5719AI sh2) and had the palely pigmented phe- notype expected if the element at the locus is inactive and unable to suppress expression of the a-ml-5719AI allele on the homolog. T w o of the families (2195 and 2197) represent progeny of the same Spm-containing plant (plant 1860-1, Table 2A), differing only by transmission of the a-m2-7991AI allele through fe- male and male gametes, respectively. Family 2199 comprises plants grown from kernels on the single ear obtained using plant 1859A-1 as pollen parent (Table 2A). Part A of the family consists of plants grown from kernels exhibiting a uniformly palely pigmented phenotype (inactive Spm) and part B, of plants grown from kernels showing one or a very few deeply pig- mented spots, signaling Spm activity in areas of the aleurone layer.
Each plant was used as both egg and pollen parent in several types of crosses to assess the activity and location of the Spm element (see MATERIALS AND METHODS). T h e purpose of crosses to and by plants carrying dSpm alleles and no active Spm element was to assess the heritability of the inactive state of the Spm element. Kernels receiving an Spm that remains inactive are colorless because the a gene is not ex- pressed when the element is not and because the
inactive element does not excise (Figure 1, a and b). Kernels that commenced development with an inac- tive element that returns to an active phase vary in phenotype, depending on the timing and frequency of element reactivation. They may have one or just a few small, deeply pigmented spots representing re- vertant cell clones produced when the element returns to an active phase and promotes its own excision. Or the kernel aleurone may have one or more larger sectors, in some cases comprising half or more of the kernel, which exhibit the palely pigmented, variegated phenotype observed with an active element (Figure 1, c and d). Each ear was scored for the number of Sh2 kernels exhibiting each of the phenotypic classes illus- trated in Figure 1. T h e results are expressed as the percentage of the Sh2 kernels that are colorless, the phenotype expected if the Spm is completely inactive throughout the kernel (Tables 3-5).
566 N. V. Fedoroff and J. A. Banks
TABLE 4
Percent kernels with an inactive Sprn transmitted through pollen of plant 1860-1"
Far on plant: Outcrosses using pollen from:
Plant No. Far I Far I1 Tiller 1 Tiller 2 stalk Tillers
2 197-3 96.7 97.7 94.8 99.2 100.0
Main
2 197-4 100.0 34.1 99.3 76.6
2197-5 98.6 98.0 57.1 93.5
2 197-6 95.9 96.5 51.9
2 197-8 100.0 68.6 90.3
2 197-9 99.5 99.6 90.3
2197-10 89.2 17.9 7.0
2197-11 86.3 63.8 17.4 9.6 11.9
2197-12 100.0 99.5 99.3
2197-13 100.0 99.2 98.4 94.6
2197-14 100.0 100.0 100.0 100.0 98.1 99.7
2197-15 98.1 51.4 39.2 20.9 94.8
2197-16 94.9 93.5 11.4 1.7
2197-17 40.8
2197-18 100.0 99.6 85.3 79.4
2197-19 100.0 100.0 100.0 100.0
2 197-20 100.0 99.5 100.0 88.4 95.7 96.4
2197-21 99.6 99.5 100.0 93.0
2 197-22 97.4 93.8 25.6 7.5 16.0
2 197-23 100.0 100.0 100.0 99.2
2 197-24 100.0 95.1 100.0
2 197-25 99.6 100.0 100.0
2 197-26 100.0 98.7 99.1 96.6
2197-27 100.0 100.0 67.3 80.0
a The percentage of Sh2 kernels receiving an inactive Spm element is recorded in t h e table.
trans-activating Spm-w element was at the a locus on the homolog and when it was located elsewhere in the genome. In the absence of a trans-activating element, the fraction of kernels on progeny ears exhibiting no Sprn activity differed between the families, among the plants in a family and, in many cases, among the ears on a single plant. In order to facilitate discussion of the observed inheritance patterns, some of the data for each of the families in Tables 2-5 have been grouped by ear and pollen source in Figure 5.
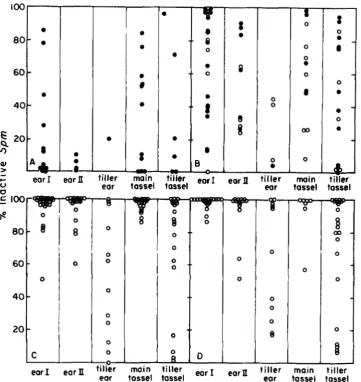
Considering only the data obtained for the first ear on plants with the a-m2-7991A1 allele (column 1 in each panel of Figure 5), it can be seen that the herit- ability of the inactive state is pronounced, but not complete. Among the progeny of plant 1860-1 (Table 2A), 90% or more of the kernels had an inactive element on 87% of the first ears. T h e results were comparable in the two families 2195 and 2 197, which differed only by the sex of the gamete through which the inactive element had been transmitted in the preceding generation (Figure 5, panels C and D). All but 1 of the 47 first ears produced among the 50 plants in the two families showed a significantly higher fraction of inactive kernels than the two ears from which the inactive kernels were selected (Table 2A, 46.5% and 52.8% inactive kernels). T h e correspond- ing fraction of first ears with more than 90% of kernels having an inactive element was lower in family
2199A (37.5%). This family comprised plants grown from kernels taken from the small fraction (1 0.1 %) of kernels with an inactive element appearing on the ear that was pollinated by plant 1859A-1 (Tables 2A and 5; Figure 5, panel B, open circles). A similar fraction of ears with a highly inactive element (31.3%) was obtained from plants grown from kernels exhibiting one or a few deeply pigmented spots signaling occa- sional reactivation of the element in endosperm tissue (family 2 199B; Figure 5, panel B, filled circles). T h e heritability of the inactive state was therefore different in the progeny of plants 1859A-1 and 1860-1 (Figure; 5, panels B, C and D) and it was markedly higher for the progeny of the plant that exhibited more extensive inactivation in the previous generation (Figure 6, panels B and D).
Spm Control in Development 567
TABLE 5
Percent kernels with an inactive Spm transmitted through pollen of plant 1859A-1’
Ear on plant: Outcrosses using pollen from:
Plant No. Ear I Ear I1 Tiller 1 Tiller 2 stalk Tillers Main
2199A-1 83.3 59.4
2 199A-2 97.1 2 199A-3 74.6 90.5 85.9
2 199A-4 0 23.8 25.5 1.4
2 199A-5 96.8 32.1 41.0 1.9 57.9
2 199A-6 80.6 27.4 7.4 10.2 76.1 32.8 1.2
2 199A-7 99.4 45.0 55.0
2 199A-8 25.5
2 199A-9 60.0 64.0 69.3
2 199A-10 8.3
2199B-1 39.4 49.2 37.4
2 199B-2 36.9 4.3 48.1 4.1
2 199B-3 45.8 0 3.1
2 199B-5 99.3 98.1 94.2
2 199B-6 85.1
2 199B-7 66.7 75.2 82.1
2 199B-8 97.1 90.5
2 199B-9 40.6
2199B-10 96.3 95.9
2199B-11 35.6 26.4 49.0 45.6
2199B-12 93.6 62.5 76.8 82.1 88.4
2 199B-13 85.5 87.8 82.1
2 199B-14 13.8 32.8 2.9
2199B-15 64.7 27.1 5.8
2199B-16 13.0 2199B-17 79.1
2199B-18 99.4 83.3 94.0 95.2
The percentage of Sh2 kernels receiving an inactive Spm element is recorded in the table.
of the families descended through the first ear from plant 1860-1 (Table 6). However, among the first ears produced on plants grown from kernels selected from the ears with the most inactive element (99-loo%), 95% had more than 90% inactive kernels (Figure 6, panel C). T h e progeny of siblings of family 2 199 gave fewer ears with a large fraction of kernels with an inactive Spm (Table 7). But once again, those families of plants grown from kernels taken from ears exhib- iting the most extensive inactivation of the Spm ele- ment give ears on which most of the kernels have an inactive element (Figure 6, panel E).
Thus the heritability of the inactive state differs in the two lines descending from the two initial ears in which Sprn inactivation was detected (Table 2A). Moreover, the difference in the heritability of the inactive state reflects the degree of Sprn inactivation in the two plants that gave rise to the two lineages. Despite the persistence of the differences through two generations, the heritability of the Spm’s inactive state approaches 100% in the progeny of plants grown from kernels of either lineage selected from ears showing >99% inactivity of the Spm element.
A n
inactive Sprn element can be reactivated dur-ing development: When a comparison is made be- tween the fraction of progeny kernels receiving an inactive Spm element through gametes produced on main stalks and tillers, it becomes immediately appar- ent that the inactive state is far less frequently trans- mitted through tiller gametes than main stalk gametes of either sex (Figure 5). T h e difference between tiller and main stalk gametes is especially striking in family 2195 (Figure 5, panel C), but is evident in all of the families for which sufficient data exist. T h e Sprn ele- ment is inactive in more than 90% of kernels receiving it through 80% of the 46 ears produced on the main stalk of the plants in family 2195 and 90% of the ears produced by using the main stalk tassels in the same family as pollen parents. By contrast, comparable lev- els of element inactivity were observed in only 29% of the 14 tiller ears in which the element’s activity could be assessed and 52% of the ears resulting from pollination using tiller tassels of family 2195 (Figure 5, panel C).
568 N . V. Fedoroff and J. A . Banks
FIGURE 5,”Percentage of kernels per ear with an inactive Sprn element. Data from Tables 2-5 are represented in panels A (families 1859, 1860, 2193, 2194); B (family 2199); C (family 2195) and D (family 2197). Each circle represents the percent of kernels with an inactive Sprn
o n one ear. Filled and open circles correspond to ears produced on plants grown from kernels with an active Sprn (0) and an inactive Sprn (0), respectively. The first two columns of each panel display the data from the first (ear I ) and second (ear 11) ears produced on the main stalk of the plant carrying the a-m2-7991AI allele. Data obtained from one tiller ear on each plant are given in the third column. The data given in the fourth and fifth columns were obtained from ears on tester plants (see MATERIALS AND METHODS) pollinated with pollen collected from the tassel produced on t h e main stalk and one of the tillers, respectively.
ears are compared in family 2 199, it appears that the fraction of kernels with an inactive Sprn element is lower in the population of second ears, than in the first ears produced on the main stalks. Similar differ- ences are apparent in the other families when the first and second ears are compared for individual plants (see for example, Table 3, plants 4, 5, 1 1 , 13 and 19). Although the magnitude of the difference between first and second ear is generally not as great as that of the difference between the first and tiller ears, the differences between first and second ears within the present lineage are nonetheless statistically significant ( P
<
0.005). Thus there is a marked systematic differ- ence in the inheritance of the inactive state through the main stalk and the tillers, and a small but signifi- cant difference upon transmission through the first and second ears produced on the main stalk. An inactive element has the highest probability of beingpropagated in the inactive phase in the first ear on the main stalk. The probability that it will be reacti- vated is higher for the second ear on the main stalk and very much higher for gametes produced by tillers.
An inactive element can be reprogrammed during development: An inactive element transmitted
through a tiller is frequently reactivated in the next generation. Table 8 gives the fraction of kernels with an inactive Spm obtained in crosses with plants grown from inactive Spm kernels selected from tiller ears or ears that received the inactive Spm through pollen collected from tiller tassels. Since element reactivation is more frequent in tillers than in the main stalk, the inactive kernels giving rise to the plants comprising the families for which data are given in Table 8 were taken from ears that had 26 to 95% of kernels with an inactive element.
Spm Control in Development 569
u) L
0
W
20 40 60 80 l o o
100
60
P:99-IOO%i
. .
20 40 60 80 1 0 0 20 40 60 80 1 0 0
% Inactive Spm
FIGURE 6,"Relationship between the Spm activity phase in parents and progeny. Each bar represents the percentage of first ears in each of several families of plants (data from Tables 2-7) that fall into the indicated intervals with respect to the fraction of kernels with an inactive
Spm. The percentage of inactive kernels (% i) on the parent ear (P) from which the kernels were selected is given in each panel. Open and shaded bars correspond to data obtained from ears on plants grown from kernels in which the Spm was active and inactive, respectively. Panel A, All first ears in families 1860, 2193 and 2194 of Table 2 (grown from kernels with an active Spm selected from 2 ears produced by plant 1530A-1). Panel B, All first ears in families 2195 and 2197 of Tables 3 and 4 (grown from kernels with an inactive Spm selected from the two parental ears produced from crosses with plant 1860-1; the data for each family are represented separately and the percentage of kernels with an inactive Spm is given for the parent ear from which the kernels were selected). Panel C, All first ears in families 2203, 2206, 221 3, 221 7, and 2218 in Table 6 (grown from kernels with an inactive Spm selected from those parental first ears in family 2003 that had 99-100% of kernels with an inactive Spm). Panel D, All first ears in family 2199 A of Table 5 (grown from kernels with an inactive Spm
selected from the single ear produced from a cross with plant 1859A-1). Panel E, All first ears in families 2225 and 2228 of Table 7 (grown from kernels with an inactive Sprn selected from those parental first ears in family 2004 that had 99-100% of kernels with an inactive S p m ) .
Panel A shows the frequency at which the Sprn element was transmitted in an inactive form by the plant that gave rise to both of the a-m2-
799141 derivatives with an inactive Spm. Panels B and D show the frequency at which the Spm was transmitted in an inactive form by the two plants (1859A-1 and 1860-1) initially giving evidence of extensive element inactivity and represent the generation following that for which data are given in panel A. Panels C and E show the frequency at which Spm was transmitted in an inactive form by plants grown from kernels selected from first ears that had received a highly inactive Spm (99-100% inactive) and represent the generation following that for which data are given in panels B and D, respectively.
inactive element is compared pairwise for plants grown from kernels that received the inactive element from the same parent, but through the main stalk or through a tiller (Figure 7). In three of the four proge- nies in which the comparison can be made, it is appar- ent that the inactive kernels taken from tiller ears or from ears that received the inactive element through pollen produced in a tiller tassel are very much more likely to produce plants in which more kernels receive an active element than those grown from inactive kernels on the first ear of the main stalk. This com- parison reveals clearly that the inactive elements had already been reprogrammed in the tillers to favor reactivation of the element during development of the next generation. Thus even in those kernels re- ceiving the Spm through a tiller gamete in which the element has not been reactivated either premeiotically or during kernel development, it has undergone a modification that predisposes it to be reactivated in the subsequent generation.
570 N. V. Fedoroff and J. A. Banks
TABLE 6
Percent kernels with inactive Spm transmitted through ear I”
Ear on plant: Outcrosses:
Percent Plant No.’ Ear I Ear I1 Tiller stalk Tiller (parent)
Spm-i
Main
2203-1 99.4 98.9 100.0
2203-2 100.0 100.0
2203-3 100.0 99.5
2203-4 100.0 100.0
2203-5 99.6 96.7 100.0
2203-6 100.0 100.0
2203-7 99.5 100.0 100.0 2203-8 2203-9 2203-10 2203-1 1 2203-12 2203-1 3 2203-14 2203-15 100.0 100.0 100.0 100.0 92.5 100.0 97.8 100.0 100.0 100.0 100.0 100.0 100.0 99.5 99.3
2206-1 100.0 100.0 100.0
2206-2 100.0 100.0
2206-3 100.0 99.6
2206-4 100.0 100.0
2206-5 100.0 100.0
2206-6 100.0 99.5
2206-7 100.0
2213-1 98.4 100.0
2213-2 100.0 100.0
22 13-3 99.5 93.3
2213-4 100.0 92.1 98.5
22 13-5 100.0 96.0
22 13-6 100.0 99.5
Far on plant: Plant No.’ Ear 1 Ear I1 Tiller
Outcrosses:
Percent
Main Spm-i
stalk Tiller (parent) 22 15-1 22 15-2 22 15-3 22 15-4 22 15-5 22 15-6 22 15-8 2217-1 221 7-2 22 17-3 22 17-4 22 17-5 22 17-6 22 17-7 22 17-8 98.3
90.7 28.4
97.2 89.2 98.6 99.6 97.4
99.5 99.0
65.0 98.1 99.0 99.0
99.1 93.2 99.4 100.0 98.3
93.4 80.4 94.9
88.4 100.0 99.7
90.1 100.0 100.0
98.5 100.0
97.6 78.4 100.0
2218-1 97.7 100.0 95.2 99.4
22 18-2 100.0
22 18-3 100.0
2218-4 98.1 99.4
2218-5 98.6 93.1
2218-6 98.7 100.0
22 18-7 100.0 100.0
2222-1 96.6 51.7 94.2 91.6
2222-2 67.2 26.6 97.1
2222-3 50.0 99.2
2222-4 89.8 98.8
2222-5 75.1 2222-6 96.4 82.2 2222-7 82.8 97.7 2222-8 62.6 83.3
a All families for which data appear in this table are progeny of family 2003, which comprises siblings of family 2195 (Figure 4). The table
*
Each family consists of plants grown from kernels taken from a single ear produced on a main stalk. The fraction of Sh2 kernels with an shows the percentage of Sh2 kernels receiving an inactive Spm element.inactive Spm on the ear from which the kernels were taken is given in the column headed “Percent Spm-i (parent).” reprogramming and the actual change in activity
phase are genetically separable: without an immediate phase change, the element can be programmed to undergo a later phase change. Therefore, the program and the phase setting must be effected by different molecular events.
The Spm element is inactivated upon transmission
through male gametes: There are systematic differ- ences in the transmission of the element’s previous state of activity through male and female gametes. It was noted early in the present analysis that some plants grown from kernels that commenced development with an active Spm element appeared to transmit the element in an inactive phase more frequently through male than through female gametes (Table 2B). One difficulty in interpreting this observation is that the endosperm is triploid by virtue of receiving two copies of the maternal haploid genome. Because elements present in the same genome can trans-activate each other, it is possible that element activity is more readily
maintained in kernels receiving two copies of the active element in the maternal genome than in kernels receiving a single copy in the paternal genome.
Spm Control in Development 57 1
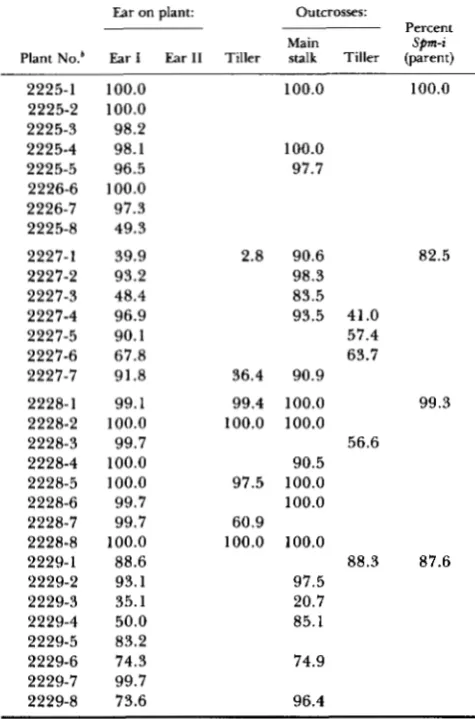
TABLE 7
Percent kernels with inactive Sprn transmitted through ear I”
Ear on plant: Outcrosses:
Percent
Plant No.’ Ear I Ear I1 Tiller stalk Tiller (parent)
Spm-i
Main
2225-1 100.0
2225-2 100.0 2225-3 98.2 2225-4 98.1 2225-5 96.5
2226-7 97.3 2226-6 100.0
2225-8 49.3
2227-1 39.9
2227-2 93.2 2227-3 48.4 2227-4 96.9 2227-5 90.1 2227-6 67.8 2227-7 91.8
2228-1 99.1 2228-2 100.0 2228-3 99.7 2228-4 100.0 2228-5 100.0 2228-6 99.7 2228-7 99.7 2228-8 100.0 2229-1 88.6 2229-2 93.1 2229-3 35.1 2229-4 50.0 2229-5 83.2 2229-6 74.3 2229-7 99.7 2229-8 73.6
100.0 100.0
100.0 97.7
2.8 90.6 82.5
98.3 83.5 93.5 41.0
57.4 63.7 36.4 90.9
99.4 100.0 99.3
100.0 100.0
90.5 97.5 100.0 100.0 60.9 100.0 100.0
97.5 20.7 85.1
56.6
88.3 87.6
74.9
96.4
All families for which data appear in this table are progeny of family 2004, which comprises siblings of family 2199 (Figure 4). The table shows the percentage of S h 2 kernels receiving an inactive Spm element.
Each family consists of plants grown from kernels taken from a single ear produced on a main stalk. The fraction of S h 2 kernels with an inactive Spm on the ear from which the kernels were taken is given in the column headed “Percent Spm-i (parent).”
male and female transmission exceeded 5% are shown in Figure 8 (panels A, B and C). It is evident that the initially inactive element is almost invariably transmit- ted in an inactive form to a larger fraction of progeny kernels through male, than through female gametes. Moreover, panel D of Figure 8 shows that similar results are obtained when the kernels commenced development with either an inactive or an active ele- ment, except that more of the active Sprn kernels grew into plants that gave ears with a high fraction of active Sprn kernels than did inactive Sprn kernels (compare Figure 8, C and D). Although Figure 8 displays data only for the plants exhibiting large differences in the transmissibility of the element’s activity phase, statis- tical analyses have been carried out for all plants in Tables 2-8 for which the relevant data are available
and show the differences in transmission through male and female gametes to be significant at the 99.9% confidence level.
The differences in the transmissibility of the inac- tive form of the Spm element through male and female gametes, as well as through the main stalk and tiller ears are illustrated in Figure 9. The four ears were produced as a result of crosses in which a single plant served as the source of all of the gametes with an inactive Sprn element. The ears show that the overall pattern of differential inheritance of the inactive Sprn element is evident in the progeny of a single plant.
An independently selected inactive Sprn element also shows differential transmission of the inactive state: Although the independently selected inactive derivative of the a-m5c allele has not been analyzed as extensively as the a - m 2 - 7 9 9 1 A l derivatives described above, enough crosses have been done to reveal a similar inheritance pattern of the inactive state of the element. As shown by the data given in Tables 9 and
10, the inactive Sprn element in this derivative is reactivated upon transmission through tiller gametes and the element is more likely to be either inactivated or maintained in an inactive form upon transmission through male than female gametes. Thus the regular alterations in the inheritance of the element’s state of activity are not idiosyncratic to either a particular insertion allele or form of the element, but appear to be characteristic of the plant’s normal developmental cycle.
DISCUSSION
572 N. V. Fedoroff and J. A. Banks
TABLE 8
Percent kernels with inactive Sprn transmitted through tiller gametes”
Ear on plant: Outcrosses: Perrenr
2204-1 2204-2 2204-3 2204-4 2204-5 2204-6 2204-7 2204-8 2204-9 2204-10 2204-1 1 2204-12 2205-1 2205-2 2205-4 2205-5 2205-6 2205-7 2214-1 22 14-2 2214-3 22 14-4 22 14-5 2214-6 22 14-7 _ _ 100.1 60.1 91.9 41.5 75.9 97.7 0.0 53.8 7.2 95.4 88.7 58.9 99.6 100.0 100.0 97.1 98.9 100.0 96.4 100.0 100.0 100.0 95.9 99.1 99.6 76.1 86.6
98.9 96.6
98.9 89.9 98.1 6.1 92.1 67.5 100.0 84.9 56.3 91.3 93.2
99.3 48.2
91.7 100.0
99.4
100.0
100.0 100.0
100.0 99.6 100.0
96.6 93.6 80.9 94.9
Plant I
No.’
Ear on plant: Outcrosses: Percent
Ear 11 Tiller Tiller (parent)
Spm-i - 2216-1 221 6-2 2216-3 22 16-4 2216-5 2216-6 2216-7 2223-1 2223-2 2223-3 2223-4 2223-5 2223-6 2223-7 2224-1 2224-2 2224-3 2224-4 2224-5 2224-6 2224-7 2224-8 2226-1 2226-2 2226-3 2226-4 2226-5 2226-6 2226-7 2226-8 1.8 3.4 8.4 6.1 1.9 36.8 11.7 91.1 55.5 54.6 47.8 29.7 69.8 42.3 33.3 7.6 85.4 39.9 49.4 1.3 0.0 57.6 13.3 83.1 41.1 33.6 92.2 56.1 22.8 55.6 10.0 1.6 62.1
6.3 9.2 4.5 6.5
94.3 58.3 86.5 91.6 85.3 18.9 87.7 83.3
44.7 87.5
0.0 17.8
98.1 83.3 67.7 98.8 87.2 26.0 1.4 80.6 78.8 57.1 43.8
Family 2226 comprises progeny of family 2004 (siblings of 2199), while the remainder comprise progeny of family 2003 (siblings of 2195). The table shows the percentage of Sh2 kernels receiving an inactive Sprn element.
’
Each family consists of plants grown from kernels taken from a single ear either produced on a tiller of a plant carrying the a-rn2-7991AI allele or which was pollinated using pollen collected from a tiller tassel of such a plant. The fraction of Sh2 kernels with an inactive Sprn on the ear from which the kernels were taken is given in the column headed “Percent Spm-i (parent).”phenotype on one or more of the tiller ears (MC- CLINTOCK 1958).
In later studies on the En element, now known to be virtually identical in sequence to the Sprn element (MASSON et al. 1987), PETERSON identified several lines with elements that exhibited novel spatial and temporal patterns of expression. Some resembled Spm isolates in which the element activity is confined to part of the kernel (MCCLINTOCK 1965; PETERSON
1966) and appear to be attributable to the phase reversal mechanism. In other cases, the pattern of element activity depended on its location, suggesting that it may have been influenced by the structure and function of the chromatin in the vicinity of the inser- tion site (PETERSON 1976a, b). FOWLER and PETERSON (1 978) described one interesting En isolate that arose in a tiller and subsequently exhibited differential expression of the element in the main stalk and tiller ears. Although they did not attribute the change to the phase reversal mechanism, the genetic properties
of the altered En element are similar to those of the Spm derivatives described here, suggesting the in- volvement of the same genetic mechanism.
T h e objective of the present experiments was to identify an Spm element, located at an insertion site for which molecular probes are available, that is undergoing the types of cyclic alterations in activity described for Sprn elements in other locations. Our results indicate that the genetic properties of the two new derivatives of Spm insertions at the a locus are similar and fully compatible with the properties of previous isolates of this type. That is, each of the derivatives arose from an element that was active by a reversible genetic change that resulted in its trans- mission to a fraction of progeny in a genetically inac- tive state. T h e present analyses of the Sprn inheritance patterns in the parent plants and several generations of progeny plants have revealed new facets of the mechanism controlling the Spm’s activity phase.
Spm Control in Development 573
ear1 tilier
2003A- I
ear1 tiller 2004A- 2
FIGURE 7,”Differential transmission of an inactive Spm element through gametes produced on the main stalk and tillers. The percentage of kernels appearing on first ears of plants that received an inactive Spm element from one of four parental plants (2003A- 1 , 2003A-4, 2003A-5 and 2004A-2) through gametes produced on the main stalk and a tiller are given in the left and right columns of each panel, respectively. The data are taken from Tables 6-8 and represent families 2203 and 2204 (panel A); families 221 3 and 2214 (panel B); families 2215 and 2216 (panel C); and families 2225 and 2226 (panel D).
MCCLINTOCK (1958) and FOWLER and PETERSON (1978), is that an inactive Sprn element frequently undergoes reactivation when it is transmitted through gametes produced on tillers. Whether the element in any particular plant will be reactivated in the tillers appears to be a function of how stably inactive it is. Although MCCLINTOCK reported that each change in activity phase was accompanied by a reprogramming of the element to give a new and heritable pattern of phase reversals, o u r results suggest that the phase setting and the program that governs subsequent phase changes are distinguishable.
Two lines of evidence indicate that an Sprn element can be reprogrammed before the phase is reset. T h e a-rn2-7991AI lines with inactive Spm elements arose from two kernels on a single plant, both of which had a weakly active Spm. These kernels gave plants that produced ears in which a significant fraction (10-
50%) of kernels received an inactive Sprn element. Some of the sibling kernels that showed no alteration in Sprn activity likewise produced plants that yielded ears with up to 85% of kernels in which the Spm element showed no activity. Conversely, some kernels which received an inactive Sprn element, produced on tillers of plants with a highly inactive element, grew into plants that transmitted the element to a majority, or even all, progeny kernels in an active state. Since the plants that exhibit such phase reversals com- menced development with the zygotic element in the same phase as that in the endosperm, it follows that the genetic events that determined the subsequent phase reversal preceded the change in phase.
Moreover, there are marked and persistent genetic differences among the progeny of an individual in
I O 0
80
$
60 0>
0
0
C
.-
+ 40
-
$ 2 00
L
3 6inactive
B , ,
3 6
inactive
7
1
9 u
inactive
I , ,
9
d
active
FIGURE 8,”Differential transmission of Spm element activity through female and male gametes. The percentages of kernels with an inactive Spm transmitted to progeny ears through the egg parent (0) and the pollen parent (8) are compared pairwise (the two values obtained for each plant are connected by a line). All values are for the first ear produced on the main stalk (0) and for tester ears that received pollen from the main stalk tassel (8). Comparisons are shown for all plants in the selected families for which the difference between egg and pollen transmission was greater than 5 % . Panel A, Families 2223 and 2224 (Table 8); both families were grown from kernels with an inactive Spm element that had received the element through a tiller ear (family 2223) or pollen collected from a tiller tassel (family 2224) of plant 2003A-10. Panel B, Family 2226 (Table 8); the plants were grown from kernels with an inactive
Spm that received it through pollen collected from a tiller tassel of plant 2004A-2. Panel C, Family 2222 (Table 6); the plants were grown from kernels with an inactive Spm that had received it through the first ear on the main stalk of plant 2003A-8. Panel D, Family 2240 (data not shown elsewhere); the plants were grown from kernels with an active Spm element that received it through the first ear on the main stalk of plant 2003A-8 (the plants in families 2222 and 2240 are therefore siblings).
which a phase change has occurred. Thus among the progeny derived from the first two plants showing significant inactivation of the Sprn element in the a-