Chimaeras and mosaics for dissecting complex
mutant phenotypes
ANDRAS NAGY
1,2and JANET ROSSANT
1,2,31Samuel Lunenfeld Research Institute, Mount Sinai Hospital, Toronto, Ontario, 2Department of Molecular and Medical Genetics, University of Toronto, Ontario and
3Department of Obstetrics and Gynecology, University of Toronto, Ontario, Canada
ABSTRACT Back at the first half of the 1980s, there was no mammalian experimental embryology in Hungary. One of us, AN, took up the challenge of establishing a small group in the field. In the absence of local information, AN and his former colleague, Andras Paldi (AP), used their tourist passport to visit several laboratories in Western Europe and collect information and advice. This is how AN and AP ended up one day sitting in Anne McLaren’s office in the MRC Mammalian Development Unit at University College, London. They never forgot her endless enthusiasm and the way she clearly explained the important points of preimplantation embryo manipulation, chimaera making and embryo transfer. As well as the extremely useful suggestions, which were crucial to starting the lab in Hungary, they also took back her deep love for embryo development. They remember her telling them, ‘never waste an embryo – there is always another unanswered question it can solve’. Many who have been lucky and experienced Anne’s spirit and advice later realized how useful it was to generate ‘new’ ideas by following the ‘not wasting’ principle. Our views on chimaeras presented below definitely contain elements which grew out from this principle.
KEY WORDS:
0214-6282/2001/$25.00
© UBC Press Printed in Spain
www.ijdb.ehu.es
*Address correspondence to: Janet Rossant. Samuel Lunenfeld Research Institute, Mount Sinai Hospital, Toronto, Ontario, M5G 1X5, Canada. FAX: ???. e-mail:
Abbreviations used in this paper:
Historical view
One of the special features of the human brain is the capacity of imagination – creating new combinations of existing elements. This capacity of the human brain is vital to many aspects of our culture: mythology, religion, technology, art and science. Imaginary crea-tures, called chimaeras, combining parts of different animals into one fierce beast, showed up independently in the ancient cradles of civilization of China, Assyria, Persia and the Roman Empire. The best known is the classic Roman ‘chimaera’, which has a serpent tail, lion head and a goat body. It is a beautiful physical representation of the chimaera from Greek mythology, who was the daughter of Typhon, the god of the hurricane, and Echidna, the snake mother. Similar imaginary creatures combining different parts of different organisms occur throughout recorded history, through the Middle Ages (see the paintings of the famous Dutch painter, Hieronymus Bosch) to our modern age of StarTrek movies. From such a rich cultural soil, it was no surprise to find that scientists were intrigued by the possibility of actually creating individuals from different embryonic origins. Mam-malian embryonic chimaeras are one such manifestation of myth made reality. Originally fierce imaginary creatures have been har-nessed and turned into a powerful and friendly tool for addressing important biological questions.
Chimaeras and mosaics
A definition of a chimaera is a single organism made up of cells of different embryonic origins. Mosaics are conceptually different from chimaeras, since in these individuals all the cells have the same embryonic origin but a random population of cells is genetically different from the rest.
use of the Cre-loxP technology to induce chromosomal alterations at will. This powerful technology is growing in use rapidly but has been reviewed extensively elsewhere (Nagy, 2000) and will not be further consid-ered here. Instead, we will focus on embryonic chimae-ras in all their different variations.
Mammalian embryonic chimaeras have been made since the 1960s, when Kristoph Tarkowski and Beatrice Mintz independently showed that two mouse embryos could be aggregated together at the 8-cell stage to generate one composite mouse that was a genetic mixture of the two parental cell lines. Those original experiments attested to the highly regulative nature of the mammalian preimplantation embryo; the embryo aggregate started at twice the normal size, but ended up as a normal-sized mouse with no abnormalities in patterning. Since that time, experimental manipulation of the early mouse embryo has been refined and
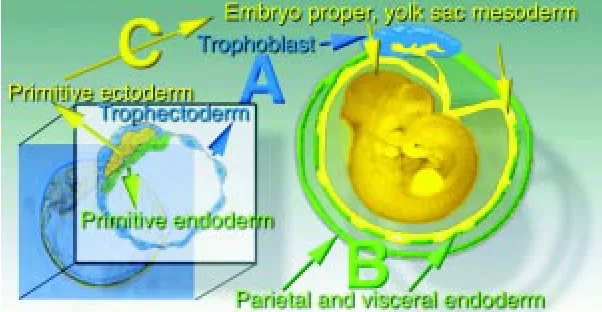
exclusively differentiate into the trophoblast layer of the placenta (Fig. 1, Lineage A), the primitive endoderm forms the parietal and visceral endoderm (Fig. 1, Lineage B) and the yolk sac mesoderm, amnion and the embryo proper are exclusively derived from the primitive ectoderm (Fig. 1, Lineage C). In the next sections, we delineate the potential of various components of chimaeras in terms of the three blastocyst cell lineages (A, B, C).
Diploid preimplantation stage embryos (D)
The developmental potential of any normal, diploid preimplantation embryo is complete. Their developmental potential is ABC (Fig. 1).
Tetraploid preimplantation stage embryos (T)
The experimental production of tetraploid embryos is very simple and efficient. Blastomeres of two cell stage embryos can be fused into one tetraploid cell by a simple electric pulse. These embryos go through preimplantation normally and implant, but rarely form proper embryonic structures beyond gastrulation. If they are aggregated with diploid embryos, they can contribute to extraembryonic com-partments but they are efficiently selected against in the embryo proper (Tarkowski et al., 1977). More specifically, they can contribute to the trophoblast and primitive endoderm lineage, but not very well to the primitive ectoderm derivatives (Nagy et al., 1990). Their developmental potential is AB (Fig. 1).
Inner cell mass (ICM)
The ICM of the late stage blastocyst consists of two layers of cells: primitive endoderm and primitive ectoderm. Its isolation is relatively easy with immunosurgery, developed by Davor Solter (Solter and Knowles, 1975). Since these cells are no longer able to differentiate into trophoblast, the developmental potential of ICM is BC (Fig. 1).
Trophoblastic vesicles (TV)
With skilled micromanipulation, it is possible to isolate pure trophectoderm vesicles from blastocyst stage embryos (Gardner, 1972). Just like ICM this is also a primary component of a blastocyst, therefore its developmental potential is the corresponding A (Fig. 1).
Embryonic stem (ES) cells
ES cells are permanent cell lines derived from the blastocyst with properties that suggest they are closest to the embryonic ectoderm
Fig. 1. Fate of the first embryonic lineages established in the blastocyst. (A)
Trophectoderm lineage; (B) primitive endoderm lineage; (C) primitive ectoderm lineage.
modified in many different ways; cells can be removed, reoriented, added back at different stages; cells from sources other than the embryo can be added; embryos can be reconstituted from their constituent cell lineages; DNA, RNA and protein can be added; and still the embryo manages to survive and compensate for the indignities to which it has been exposed. Anne McLaren was a pioneer in the application of chimaera techniques to understanding the allocation of cell lineages in development and her monograph in 1976 entitled “Mammalian chimaeras” laid out the tenets of the field and formed the intellectual basis for much succeeding work on chimaeric analysis.
The utility of the original aggregation chimaeras for following the fate of genetically marked cells was somewhat restricted. First, aggregation at the 8-cell stage produced conceptuses in which all cell lineages, both embryonic and extraembryonic, were geneti-cally mixed and, second, the markers used to follow lineage were not ubiquitous or detectable in situ at a single cell level. The first problem was alleviated in the late ‘60s when Richard Gardner in the UK and Ralph Brinster in the US developed techniques for intro-ducing different cell types into the blastocyst stage of development. The second problem was intractable for a much longer time but now has been solved by the development of easily detectable in situ enzymatic markers and by the viable cell marker, Green Fluorescent Protein. In this review we will discuss the various kinds of embryonic chimaeras that can be generated and what different kinds of information can be obtained from them, using examples mostly from our own work.
The basic building blocks of chimaera production
lineage of the blastocyst. Chimaera studies showed that their devel-opmental potential is C, with no capacity to contribute to the primitive endoderm and trophectoderm (Nagy et al., 1990) (Fig. 1).
Trophoblast stem (TS) cells
The recently established TS cells are derived from the trophecto-derm lineage and just like ES cells they retain many features of their origin, including developmental potential (Tanaka et al., 1998), which is now in this case A (Fig. 1).
Somatic stem cells
This is an emerging and very exciting area of exploration and discovery, which is challenging the dogma of renewal and regenera-tion. Many adult tissues contain stem cells, which have been consid-ered to have self-renewal capacity but restricted developmental potential. However, recent studies have shown that adult restricted stem cells can have much broader differentiation capacity than originally thought. For example, haematopoietic stem cells can contribute to muscle lineages, neuronal stem cells can apparently contribute to multiple cell types, and muscle stem cells can make haematopoietic lineages (Clarke et al., 2000; Gussoni et al., 1999; Jackson et al., 1999). Identifying and isolating these stem cells is not easy due to their small number and more are expected to be discovered. At this stage not many have been tested for their capacity to contribute to embryonic lineages in chimaeras. However, this is a critical test for multipotentiality and will be an important area of research for the future.
Legitimate combinations and component allocations
If the possible components of chimaera production are put to-gether into a single matrix, where the rows are the three primary lineages of development (A, B and C, as above) and the columns are the components (Fig. 2), it is immediately obvious that not all the possible combinations of two components could cover all the three lineages, which is a minimal requirement for an embryo to develop. Those combinations where the two components together cover all the lineages and, therefore, allow embryonic development are
shown in Fig. 3. Untested but potentially legitimate combinations are shown with a question mark.
The differing developmental potential of the different chimaera components means that it is possible to generate chimaeras with chimaerism restricted to different embryonic lineages. This capacity has been extremely useful in both cell lineage analysis and dissec-tion of complex mutant phenotypes, as described below.
Applications
Diploid embryoóDiploid embryo chimaeras
This type of chimaera is one of the easiest to produce by aggregating 8-cell stage embryos. It is very useful for examining mutant vs. wild-type cell behaviour in a competitive situation and addressing the question of cell autonomous phenotypes (Rossant and Spence, 1998). Since chimaeras are made by aggregating the embryos from a heterozygous cross with wild-type embryos, it is necessary to be able to distinguish the three possible classes of chimaeras. This can be achieved by using two different mutant alleles to generate homozygous embryos, or by use of linked markers. In the case of imprinted genes, chimaera analysis is made easy by the fact that heterozygous embryos will have the mutant phenotype if they inherit the mutant allele on the non-imprinted chromosome. We took advantage of this in chimaeric analysis of Mash2 mutants. Earlier we had shown that a targeted null mutation in the Mash2 gene resulted in a midgestational embryonic lethality (Guillemot et al., 1994). The dying embryos had a severe placenta phenotype characterized by the lack of spongiotrophoblast and defective labyrinth layers. In order to address the cell autonomous nature of the phenotype we performed a chimaeric study (Tanaka et al., 1997). One component of the chimaeras was wild-type and the other was Mash2 deficient, and tagged by ubiquitous lacZ expres-sion of the Rosa26 transgene. In the chimaeras, the Mash2 deficient cells were completely excluded from the spongiotrophoblast lineage, strongly supporting the cell autonomous nature of the Mash2 defi-ciency in this lineage. On the other hand, Mash2 deficient cells could contribute at very high levels to all other layers of the placentas, including the labyrinthine layer, without showing any signs of abnor-mality. Therefore, the labyrinthine phenotype observed in the Mash2
Fig. 2. Developmental potential (green = full, red = restricted) of the possible chimaera components in the three primary embryonic line-ages shown in Fig. 1. D, diploid embryo; T tetraploid embryo; ICM, inner cell mass; ES, embryonic stem cell; TV, trophoblastic vesicle; TS, trophob-last stem cell.
deficient animals was presumably secondary to the spongiotrophoblast defect. This kind of study is important in focussing the search for downstream effectors of a gene’s action on a particular cell lineage.
Diploid embryoóTetraploid embryo chimaeras
The skewed distribution to the different embryonic lineages achieved in diploid embryoótetraploid embryo chimaera, can be used to advantage to distinguish primary placental or embryonic phenotypes. We first used this technique with the Mash2 mutants, where we suspected that the embryonic lethal phenotype was due solely to a placental function for Mash2. We aggregated wild-type tetraploid embryos with diploid embryos from a cross of Mash2 +/-heterozygotes. With the tetraploid embryos providing cells in the placenta, Mash2 homozygous embryos were capable of surviving pre- and perinatal development. Adult Mash2 homozygotes were normal and fertile, clearly demonstrating that Mash2 gene function is dispensable in the embryo proper and for postnatal life and it is solely required for placental development (Guillemot et al., 1994).
We have gone on to use the same approach to rescue the placental phenotypes of a number of mouse mutations, including ERR_ and Brca1 (Hakem et al., 1996; Luo et al., 1997). The approach is generally useful for revealing later phenotypes obscured by early placental defects, e.g. PPARγ (Barak et al., 1999). So far Mash2 is unique in showing complete phenotypic rescue by this approach; most genes involved in placental function appear to have some later embryonic or postnatal function. This type of chimaera is similar to the ES cellótetraploid embryo chimaera in outcome. Its main advan-tage is that viable mutant offspring may be born if the extraembryonic phenotype can be fully rescued, whereas such an outcome is rare when ES cells are used. Its main disadvantage is that the skewing of distribution between embryonic and extraembryonic lineages is not as complete as for ES cellótetraploid embryo chimaeras.
Diploid embryoóICM chimaeras
ICMs can be aggregated with 8-cell embryos or injected into blastocysts, and will contribute to all lineages except the trophoblast. Chimaeras of this sort were fundamental to the delineation of the fate of early cell lineages in the mouse embryo and to the establishment of the time at which different lineages became committed (Rossant, 1987). In addition, it was the observation that a single ICM cell could contribute to all somatic cell lineages including the germ line (Gardner, 1985), that led to the drive to develop permanent cell lines from the ICM as a vehicle for introducing genetic changes into the germ line.
Diploid embryoóES cell chimaeras
The development of ES cell lines did indeed allow the power of chimaeras to be applied to the generation of many kinds of geneti-cally altered mice. ES cells can be manipulated genetigeneti-cally in vitro, mostly notably by various homologous and site-specific recombina-tion approaches (Rossant and Nagy, 1995), and then used to generate chimaeras for transmission of the altered genotype into the germ line.
However, embryo-ES cell chimaeras are more than just a vehicle for germ line transmission of mutations. ES cells, homozygous for a mutation of interest, are extremely useful for analyzing the lineage-specificity of mutant defects. Although mutations generated in ES cells are generally only at one allele, the cells can be made ho-mozygous by a variety of means. These include loss of
heterozygos-ity induced by increased drug selection, gene targeting at the second allele, or generation of new homozygous lines from homozygous embryos (Rossant and Nagy, 1995). In all cases, it is also possible to introduce markers to help follow mutant cells in chimaera
Having homozygous ES cells makes the task of generating large numbers of mutant ó wild-type chimaeras much easier, and this approach is now widely used for chimaeric analysis of mutants. We have used it in a number of studies, including study of the complex gastrulation phenotype of Fgfr1 mutants (Ciruna et al., 1997). Chimaeric analysis revealed that the phenotypic effects appeared to be separable into an effect on cell adhesion, manifested by the failure of mutant cells to migrate through the streak, and an effect on mesoderm cell-type specification. Rescue of the early gastrulation phenotype in the chimaeras also revealed later tissues where Fgfr1 mutant cells were excluded, which thus were likely to be targets of Fgfr1 action. These included the progress zone of the limb buds (Saxton et al., 2000), the branchial arch mesenchyme and the gut. Because ES cells do not contribute to the trophectoderm and primitive endoderm lineage, diploid embryoóES cell chimaeras can help delineate the primary site of action of mutations causing early patterning defects. If chimaeras made by mixing wild-type ES cells with mutant embryos always show the mutant phenotype whatever the extent of ES cell contribution, then it is likely that the phenotypic effect resides in the trophoblast or primitive endoderm. In the reverse experiment, rescue of the phenotype by introduction of mutant ES cells into wild-type embryos also implies an extraembryonic effect. Experiments of this sort have been critical in establishing the role of the visceral endoderm in anterior-posterior patterning, via the action of the nodal-Smad2 signaling pathway (Varlet et al., 1997; Waldrip et al., 1998).
Diploid embryoóTS cell chimaeras
TS cells, which derive from the trophectoderm of the blastocyst, produce chimaeras in which only the trophoblast lineages are genetically mixed (Tanaka et al., 1998). This combination of lineage contributions is not available from any other chimaeric situation and is potentially very powerful for trophoblast-specific rescue of mu-tants. However, to date, the extent of chimaerism achieved by injection of TS cells into blastocysts is not consistently high, making mutant rescue difficult. We have attempted to rescue Mash2 mutants by injection of wild-type TS cells without success so far (unpublished results). However, improved contribution of TS cells to chimaeras should be possible by introduction of TS cells into earlier embryos. Clearly more technical work is needed before these chimaeras can be used to their full potential.
Tetraploid embryoóICM chimaeras
This particular combination is rarely used but can generate mice entirely derived from the ICM genotype very efficiently. We used this as a control when trying to generate ES-cell derived offspring from tetraploid embryoóES cell aggregates (Nagy et al., 1990). ICM-derived offspring from tetraploid embryo aggregates survive at a much higher rate than ES-derived offspring, showing that the neonatal problems of ES-derived mice stem from their ES cell origin not the mode of chimaera production.
Tetraploid embryoóES cell chimaeras
and trophectoderm lineages (from the tetraploid cells) (Nagy et al., 1990). We used this to show that it was possible to produce viable entirely ES-derived mice, at least from early passage ES cells (Nagy et al., 1993). This demonstrated unequivocally the pluripotential nature of ES cells and is a critical underpinning of the current interest in the derivation of human ES cells for possible use in tissue repair of all cell types.
However, the tetraploid embryoóES cell chimaera has much wider uses. It is the cleanest way to assess the relative roles of a particular gene in embryonic versus extraembryonic lineages. For example, we have used tetraploid embryo-ES cell aggregates to show that early gastrulation effects of the HNF3β mutation reside in the primitive endoderm, but that later defects in the node and notochord are due to defects in the primitive ectoderm lineage (Dufort et al., 1998). Wild-type tetraploid embryoómutant ES chimaeras showed normal gastrulation but absence of node and notochord, while mutant tetraploid embryoówild-type ES cell chimaeras pro-duced the mutant gastrulation defects.
If the effect of a particular mutation is confined to the primitive ectoderm lineage, tetraploid embryoómutant ES cell chimaeras can be a very useful way of generating large numbers of mutant embryos for analysis. For example, we use this routinely to generate Fgfr1 mutant embryos for analysis, since this mutation shows no trophob-last or primitive endoderm defects.
In the previous example, the homozygous mutant phenotype was known from standard breeding of heterozygotes first and then duplicated in tetraploid embryo aggregates. However, it is also possible to analyze the primary mutant phenotype in tetraploid embryo chimaeras and thus avoid germline transmission. We exam-ined both the heterozygous and homozygous phenotype of deletion of the Vegf-A gene by this approach (Carmeliet et al., 1996). Embryos heterozygous for loss of Vegf-A showed embryonic lethality due to reduced vascularity. Thus, no live heterozygotes could even be produced to assess the homozygous phenotype. We generated homozygous mutant ES cell lines and used them to generate homozygous embryos by tetraploid embryo aggregation. These embryos showed an even more severe vascular defect similar to the mutant defect in the knock-out of the VEGF receptor, Flk1 (Shalaby et al., 1995). In this particular instance, analysis of the homozygous phenotype by tetraploid embryo aggregation was the only possible route. However, because it speeds up considerably the time from identification of targeted cell line to analysis of mutant phenotype, it can also be used as a means of gaining a quick insight into mutant phenotype. This is useful for early embryonic lethal phenotypes but should not be relied on for late gestation defects onwards. Even wild-type ES cellótetraploid embryo chimaeras tend to show reduced viability close to term and so it becomes difficult to separate mutant effects from generalized ES cell defects.
ICMóTrophectoderm vesicle chimaeras
This masterpiece of micromanipulation involves injection of ICMs into microsurgically prepared trophectoderm vesicles. This gener-ates a reconstituted blastocyst with ICM and trophectoderm of separate genotypes. This was the original tool used to determine the later lineage derivatives of ICM and trophectoderm (Gardner et al., 1973). It was also used to rescue trophoblast defects before the development of the tetraploid approach. For example, we showed that embryos from the divergent mouse species, Mus caroli, fail to develop in the M. musculus uterus. However, M. caroli ICMs in M.
musculus trophectoderm vesicles can implant and develop normally (Rossant et al., 1983). This implies that species-specific trophoblast-uterine interactions may be a critical component of speciation. Rescue of endangered species by interspecific transfer may be enhanced by provided trophoblast of host genotype by either this microsurgical approach or the easier tetraploid embryo aggregation method. Although tetraploid embryo aggregation has tended to supercede the reconstituted blastocyst approach, because of its simplicity, tetraploid embryo aggregation does not lead to the imme-diate separation of the lineage components. Thus, the reconstituted blastocyst approach may still be necessary to dissect apart very early lineage defects (Gardner et al., 1990).
Addendum
It is clear that the chimaera is more than a mythological curiosity, but a living, breathing manifestation of our curiosity about the origins of development from egg to embryo to adult. Manipulation of the preimplantation mouse embryo brings together the power of both experimental embryology and genetics to further our understanding of our own origins. Anne McLaren has throughout her career recog-nized the importance of combining embryology and genetics and has led the way in promoting their convergence. Her early work on defining the parameters necessary for successful embryo transfer was critical for all succeeding work in the field. Her recognition of the power of chimaeras was critical for her own studies on sex determi-nation, and her lucid exposition of the questions and problems in the field in many different publications was also the catalyst for many of the rest of us to pursue the questions she raised. Anne has been a wise and supportive presence in the field of mammalian develop-mental biology and her influence continues to be felt worldwide. It is always a pleasure to meet with Anne, discuss science and respond to her questions, which always get to the critical issues of the day. When giving a talk, she is scrupulous in her acknowledgement of other people’s contributions, and so it is a pleasure to contribute to a whole issue where we in turn acknowledge and applaud her major contributions to our research programs.
References
BARAK Y., NELSON, M.C., ONG, E.S., JONES, Y.Z., RUIZ-LOZANO, P., CHIEN, K.R., KODER, A., and EVANS, R.M. (1999). PPAR gamma is required for placental, cardiac, and adipose tissue development. Mol. Cell. 4: 585-95.
CARMELIET, P., FERREIRA, V., BREIER, G., POLLEFEYT, S., KIECKENS, L., GERTSENSTEIN, M., FAHRIG, M., VANDENHOECK, A., HARPAL, K., EBERHARDT, C., DECLERCQ, C., PAWLING, J., MOONS, L., COLLEN, D., RISAU, W., and NAGY, A. (1996). Abnormal blood vessel development and lethality in embryos lacking a single VEGF allele. Nature 380: 435-9.
CIRUNA, B.G., SCHWARTZ, L., HARPAL, K., YAMAGUCHI, T.P., and ROSSANT, J. (1997). Chimeric analysis of fibroblast growth factor receptor-1 (Fgfr1) function: a role for FGFR1 in morphogenetic movement through the primitive streak. Develop-ment 124: 2829-41.
CLARKE, D.L., JOHANSSON, C. B., WILBERTZ, J., VERESS, B., NILSSON, E., KARLSTROM, H., LENDAHL, U., and FRISEN, J. (2000). Generalized potential of adult neural stem cells. Science 288: 1660-3.
DUFORT, D., SCHWARTZ, L., HARPAL, K., and ROSSANT, J. (1998). The transcrip-tion factor HNF3beta is required in visceral endoderm for normal primitive streak morphogenesis. Development 125: 3015-25.
GARDNER, R.L. (1985). Clonal analysis of early mammalian development. Philos Trans R Soc Lond. B. Biol. Sci. 312: 163-78.
GARDNER, R.L., BARTON, S. C., and SURANI, M.A.H. (1990). Use of triple tissue blastocyst reconstitution to study the development of diploid parthenogenetic primitive ectoderm in combination with fertilization-derived trophectoderm and primitive endoderm. Genet.Res. 56: 207-220.
GARDNER, R.L., PAPAIOANNOU, V.E., and BARTON, S.C. (1973). Origin of the ectoplacental cone and secondary giant cells in mouse blastocysts reconstituted from isolated trophectoderm and inner cell mass. J.Embryol.Exp.Morph. 30: 561-572.
GUILLEMOT, F., NAGY, A., AUERBACH, A., ROSSANT, J., and JOYNER, A.L. (1994). Essential role of Mash-2 in extraembryonic development. Nature 371: 333-6.
GUSSONI, E., SONEOKA, Y., STRICKLAND, C. D., BUZNEY, E. A., KHAN, M. K., FLINT, A.F., KUNKEL, L.M., and MULLIGAN, R.C. (1999). Dystrophin expression in the mdx mouse restored by stem cell transplantation. Nature 401: 390-4.
HAKEM, R., DE LA POMPA, J. L., SIRARD, C., MO, R., WOO, M., HAKEM, A., WAKEHAM, A., POTTER, J., REITMAIR, A., BILLIA, F., FIRPO, E., HUI, C.C., ROBERTS, J., ROSSANT, J., and MAK, T. W. (1996). The tumor suppressor gene Brca1 is required for embryonic cellular proliferation in the mouse. Cell 85: 1009-23.
JACKSON, K. A., MI, T., and GOODELL, M.A. (1999). Hematopoietic potential of stem cells isolated from murine skeletal muscle. Proc. Natl. Acad. Sci. USA 96: 14482-6.
LUO, J., SLADEK, R., BADER, J.A., MATTHYSSEN, A., ROSSANT, J., and GIGUERE, V. (1997). Placental abnormalities in mouse embryos lacking the orphan nuclear receptor ERR-beta. Nature 388: 778-82.
MOHRENWEISER, H., and ZINGG, B. (1995). Mosaicism: the embryo as a target for induction of mutations leading to cancer and genetic disease. Environ. Mol. Mutagen. 25: 21-9.
NAGY, A. (2000). Cre recombinase: the universal reagent for genome tailoring. Genesis 26: 99-109.
NAGY, A., GOCZA, E., DIAZ, E.M., PRIDEAUX, V.R., IVANYI, E., MARKKULA, M., and ROSSANT, J. (1990). Embryonic stem cells alone are able to support fetal development in the mouse. Development 110: 815-821.
NAGY, A., ROSSANT, J., NAGY, R., ABRAMOW-NEWERLY, W., and RODER, J.C. (1993). Derivation of completely cell culture-derived mice from early-passage embryonic stem cells. Proc. Natl. Acad. Sci. USA 90: 8424-8.
ROSSANT, J. (1987). Cell lineage analysis in mammalian embryogenesis. Curr.Topics Dev.Biol. 23: 115-146.
ROSSANT, J., CROY, B.A., CLARK, D.A., and CHAPMAN, V.M. (1983). Interspecific hybrids and chimeras in mice. J. Exp. Zool. 228: 223-33.
ROSSANT, J., and NAGY, A. (1995). Genome engineering: the new mouse genetics. Nat. Med. 1: 592-4.
ROSSANT, J., and SPENCE, A. (1998). Chimeras and mosaics in mouse mutant analysis. Trends Genet. 14. 358-63.
SAXTON, T. M., CIRUNA, B.G., HOLMYARD, D., KULKARNI, S., HARPAL, K., ROSSANT, J., and PAWSON, T. (2000). The SH2 tyrosine phosphatase shp2 is required for mammalian limb development. Nat. Genet. 24: 420-3.
SHALABY, F., ROSSANT, J., YAMAGUCHI, T.P., GERTSENSTEIN, M., WU, X.F., BREITMAN, M.L., and SCHUH, A.C. (1995). Failure of blood-island formation and vasculogenesis in Flk-1-deficient mice. Nature 376: 62-6.
SOLTER, D., and KNOWLES, B.B. (1975). Immunosurgery of mouse blastocyst. Proc. Natl. Acad. Sci. USA 72: 5099-5102.
TANAKA, M., GERTSENSTEIN, M., ROSSANT, J., and NAGY, A. (1997). Mash2 acts cell autonomously in mouse spongiotrophoblast development. Dev. Biol. 190: 55-65.
TANAKA, S., KUNATH, T., HADJANTONAKIS, A-K., NAGY, A., and ROSSANT, J. (1998). Promotion of trophoblast stem cell proliferation by FGF4. Science 282: 2072-5.
TARKOWSKI, A.K., WITKOWSKA, A., and OPAS, J. (1977). Development of cytochalasin B-induced tetraploid and dipoid/tetraploid mosaic mouse embryos. J. Embryol. Exp. Morph. 41: 47-64.
VARLET, I., COLLIGNON, J., and ROBERTSON, E.J. (1997). nodal expression in the primitive endoderm is required for specification of the anterior axis during mouse gastrulation. Development 124: 1033-44.