THE IDENTIFICATION OF COMPLEX GENOTYPES
IN
BACTERIOPHAGE T4. I: METHODS
A. H. DOERMANN AND LINDE BOEHNER
Department of Genetics, University of Washington, Seattle, Washington 98105
Revision received August 17, 1970
HE
simultaneous use of more than three genetic markers in bacteriophage Tcrosses is infrequent, and where four or more have been used in a single cross, usually only one of the numerous progeny genotypes has been identified as a unique class among the complex mixture of genotypes. Foss and STAHL (1963) have made use of a 4-factor cross in which all 16 segregant genotypes could be identified. More frequently, however, selective tests have been used to score one identifiable type, as, for example, r+ in a complex progeny containing multiplerZZ markers (CHASE and
DOERMANN
1958). While that type of investigation has yielded much important information, the lack of methods for identification of a larger number of genotypes in a single progeny has severely limited the ways in which multi-factor crosses can be used.The present paper describes techniques by which that handicap can be over- come. The procedures are based on the same considerations as those underlying the replica plating procedure employed by CHASE and DOERMANN (1958) for isolating multiple rZZ strains of T4. They have the advantage, however, that the uncertainties present in the earlier method of replicating with velvet from la t o p layer of soft agar are eliminated. Moreover, any plaque can be used in countless individual tests, thus making possible the identification of many markers in the single phage particle which originated the plaque. This method has been used successfully in our laboratory for simultaneous identification of an acriflavine- resistant mutant, a host-range type, and seven rZZ markers. It has also been used to identify the ubiquitous amber mutants discovered by &STEIN (see EPSTEIN
et al. 1963) and presumably could be applied to the temperature-sensitive mu- tants of EDGAR and LIELAUSIS (1964). In general, it seems that a variation of the procedure can be developed to identify any marker for which a selective test is available and whose reversion index and/or transmission coefficient are not too high.
MATERIALS A N D METHODS
Bacterial strains: The properties of several Escherichia coli strains used are crucial for dis- tinguishing the various phage alleles. Two are used for making plaques of the phage particles to be tested. The strain S/6 is used whenever amber mutants are not included in any of the genotypes under scrutiny because r and tu mutants produce distinct plaque morphologies on that plating IThis investigation was supported by Public Health Service Research grant GM 13280. Much of this work was carried out in the Department of Molecular Biology, Vanderbilt University, Nashville, Tennessee, where it was sup- ported by Public Health Service Research grant CA 04437.
41 8 A. H. D O E R M A N N A N D L. B O E H N E R
host (DOERMANN and HILL 1953). The ambers ( a m ) , a class of mutant which will not grow on S/6 or E . coli B (EPSTEIN et al. 1963), are plated on CR-63. On CR-63, as i n all strains of the A-sensitive K type, T4rII mutants display a wild-type plaque morphology rather than the r type shown on B or S/6. Even though the r type plaque morphology of r l l phages is not observed on this host, the testing methods presented here allow identification of the r l l mutations present in the phage genome.
In making the various kinds of test plates, several streptomycin-resistant bacteria are needed. Strains B/s and K-I2(X)/s have been described by CHASE and DOERMANN (1958). When amber mutations are included in the genomes, K-l2(h)/s must be replaced by K112-12(Xh)#3/s. A streptomycin-resistant variant of E . coli strain BR-16 (originally obtained from Professor C. BRESCH) is also useful. The distinctive property of BR-I6 for the present experiments, at least, is that while it is infected and killed by either h42 or am mutants of T4, it prduces phages of neither type unless a complementing strain is also present in the same cell. Table 1 summarizes the bacterial strains used and their properties which are pertinent to the present tests.
Phages: All phages used in the methods described here are mutants of T4D, but T4B mutants can be used in a similar manner. A number of r l l mutants from various sources have been used, principally those mapped first by EDGAR (1958) and EDGAR et al. (1962). An important factor for obtaining clear-cut tests for any particular mutant is that its reversion index should not be too high. For example, difficulties are encountered with r l l tests of mutants whose reversion index exceeds 5 x 10-7. Other mutants which can be satisfactorily identified in the manner to be described are ac41 and 941, both of which will grow in the presence of concentrations of acriflavine (or other acridine dyes) which are not tolerated by the wild-type T4. These mutants were obtained from Dr. R. S. EDGAR. The former has been described by EDGAR and EPSTEIN
(1961) and the latter by PIECHOWSKI and SUSMAN (1967). The host-range mutant, h42 (EDGAR 1958), is also well suited to the testing method because of the availability of the bacterial strain
TABLE 1
Bacterial strains used
Properties related to identification
Bacterial strain Symbol of T4 genotypes
CR-63 (orCR-63/1) CR Permissive host for all genetic
S/6
CR-63/s
B/s
K-12( h ) / s *
K112-12(hh) #3/s
BR-l6/s
types.
S/6 Gives good plaque morphology. Restrictive for am mutants. Like CR, but streptomycin re- CR/s
sistant.
B/s Streptomycin resistant. Restric- tive for am mutants.
K (1) /s Streptomycin resistant. Restric- tive for both rll and a m
mutants.
K(Xh)/s Streptomycin resistant. Restric- tive for r l l , permissive for am mutants.
BR/s Streptomycin resistant. Restric- tive for h42 and am mutants.
Reference
EPSTEIN et al. 1963
DOERMANN and HILL 1953
Isolated from CR-63
CHASE and DOERMANN 1958
CHASE and DOERMANN 1958
Isolated from K112-12(Xh) #3
(WOLLMAN 1953)
Isolated from BR-I6 which was obtained from C. BRFSCH
I D E N T I F I C A T I O N O F GENOTYPES IN P H A G E T 4 41 9 BR-16. Finally, the a m mutant series developed by EPSTEIN is a particularly important group because of its widespread distribution in the T 4 genome. From this large group, mutants with low reversion indices were selected for determining the general applicability of the testing method to this category of mutant.
General methods: Except for the differences noted below, the media and procedures outlined by CHASE and DOERMANN (1958) were followed here. One difference was that the recipe for bottom-layer agar was modified by altering the sodium citrate concentration. For S/6 platings, plaque morphology seemed optimum with 1.1 grams of sodium citrate dihydrate per liter of medium. When tu and tu+ plaques are to be distinguished, however, in platings on CR or K
bacteria, “citrate-rich” (6.25 grams of sodium citrate dihydrate per liter) plate agar may be used. Plaque mwphology was best when S/6 plates were incubated a t 34°C and CR-63 and
K plates were incubated at 37°C for 18-24 hr.
A second deviation from the techniques of CHASE and DOERMANN was that all platings from which plaques were to be tested were made with exponentially growing cultures of plating bacteria prepared according to the formula of EPSTEIN (1958). These will henceforth be called “plaque plates” to distinguish them from “test plates” onto which samples from the plaque contents are transferred. The test plates are made with overnight aerated cultures.
For making plaque plates it has been our experience that the closed brewer’s type covers made of aluminum with an absorbent liner yielded more reliable results than disposable plastic plates or plates with glass covers. The latter plates, even when incubated overnight before use, frequently showed condensed moisture around the edges. Occasionally the moisture ran over the plate, presumably mixing phage from various kinds of plaques. Any particular plaque tested would, therefore, contain an element of uncertsinty. With the aluminum covers, plates could be used as s o m as the medium had solidified, and the danger from moisture runs was largely eliminated.
Test plates are poured in glass-top plates. They, too, are used as soon as the medium has solidified, but moisture condensed on the covers is wiped out. The reason glass covers are neces- sary here is that test plates are ospen to the atmosphere for an appreciable interval while tests are being made, and they tend to dry out during that time. When aluminum covers are used for test plates, the additional loss of water through the absorbent disk during incubation is excessive and makes phage growth so sparse that tests become dubious. All the various kinds of
test plates contain 4&50 ml of plate agar per 100 mm diameter plate or 70-100 ml per 150 mm diameter plate.
APPLICATIONS
Method Z: Testing individually with glass rods
420 A. H. D O E R M A N N A N D L. B O E H N E R
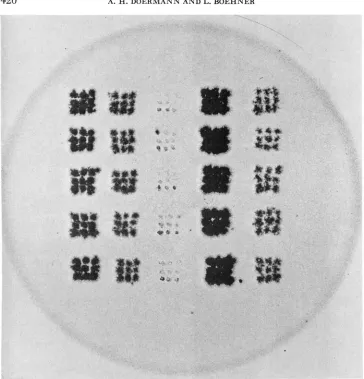
FIGURE 1.-An r70 test plate using Method I. 1.5 X. The bacterial strain K(h)/s was used and the plate incubated 24 hr. The 5 columns represent 5 genotypes of which 5 plaques each we:e tested. The genotypes tested are, from left to right, r73, r77, r70, wild type, and rX4 r77 (r73 is a mutation in the rlZB cistron, and the others are all mutations in rZZA). The test plate itself was used as a negative by inserting it into a photographic enlarger and making the print directly. The dark spots therefore indicate areas of clearings.
IDENTIFICATION O F GENOTYPES I N P H A G E T4 421
drawn to a point, broken off, and fire-polished so that the rounded tip has a diameter of 0.5 to 1.0 mm. It is advisable to leave the test plates uncovered no more than 40 min and no less than 20 min during the testing procedure. Local temperature and humidity conditions may, of course, alter these limits.
After incubation overnight a t 37"C, those spots which originated from plaques containing 7-70 phage d l be completely turbid, while those which contain r70+ phage will give clear areas (see Figure 1 ) .
A
further distinction can also be made in the rZZ test plates which is sometimes useful. A test plate seeded with a phage which contains an rZZA (BENZER 1957) marker will show a completely clear spot for any plaque tested which contains no rZZA marker. This is presumably because phage growth can begin immediately in all mixedly infected cells. If, however, the phage being tested contains an rZZA marker at a different site from the r in the test-plate seeding, a more speckled clearing will result since phage reproduction in K ( h ) / s cannot proceed until they are infected by an r+ recombinant emerg- ing from the permissive B/s cells. This difference may be seen in Figure 1.To decide whether other non-rZZ sites are wild type or mutant, similar tests are made, except that the ingredients of the test plates are different, The composition of the test plate for various other types of markers is given in Table 2. Under some conditions it is useful to incubate test plates as long as 48 hr. This is gener- ally not necessary unless closely linked markers in a single cistron are involved.
The use of copy p2ates: As the number of tests to be made from a single plaque
becomes larger, two types of problem develop: First, the plaques tend to dry out while uncovered during the prolonged period required for many tests from a single plate. Secondly, the smallest plaques in particular become so macerated from many stabs that it is not always certain that the later stabs are made in the phage-rich area of the original plaque. Both of these problems can be averted by use of copy plates. After 18-24 hr incubation of the plaques which are to be tested, they are stabbed with glass rods and copied on fresh plates seeded with either S/6 or CR-63. Glass-top plates are used for this purpose and are left un- covered for a 20-40 min interval similar to test plates. It has proved useful to make, for each plaque being copied, several stabs close together on the copy plate. This gives a continuous clearing for even the smallest plaques copied. After 18-
24 hr of incubation, tests are made from the copies. Such plates stored in the re- frigerator for as long as a week give dependable tests, even with difficult marker combinations.
Errors when rare multiple recombinations are required during incubation of
the test plate: The method, even in the first trials yielded completely unambigu-
ous results when used in testing single-r plaques against single-r test plates. It
422 A. H . D O E R M A N N A N D L. B O E H N E R
TABLE 2
Conditions used in making tests for specific mutants of T4D'
Titer of phage Quality
Mutant+ suspension Bacterial mixture$ of tests
amB23 amB16 amNl32 amEl8 a d 7 6 amE389 amS71 amB6 amN85 amS6 amN54 a d 4 5 3 amB25 amN58 amA455 a d 2 6 5 amN52 amN82
amB256 amN135 amE609 amN53 amB17 a d 5 2 amS29 amN122 amB22
amC42 amE1 7
amN93 amNI28 amN69
amB251 amN5O amN98 amE1236 amB258
amE355 amE51 amS2 amN81 amN130
amB2O amE509 amE9OO
5 x 108
e x 108
5 x 1 0 7
5 x 108
A A
B B A A
10 B/s : 1 CR/s
10B/s : 1 CR/s
20B/s : 1 CR/s
10 B/s : 1 CR/S
20B/s : 1 CR/s
50 B/s : 1 CR/s
20 B/s : 1 CR/s
A A A
I D E N T I F I C A T I O N O F GENOTYPES IN P H A G E T 4 423
am8-82 amH26 amB255 a d 6 6 amN131 amS105 amS92 a d 6 0 amB252 amH17 rll rll rb41 rb41 ac41 h42 (ZYS) (ZYS)
( 1 0 )
(16) (26) ( 2 6 ) (27) ( 2 7 ) (35)
( 5 2 ) (absence of
a m mutants) (presence of
a m mutants) (absence of
a m mutants) (presence of
a m mutants)
Titer of phage Quality
sumension Bacterial mixturet of tests
5 x 107
5 x 107
2 x 108 5 x 107 I x 105
1 x 108 5 x 108
5 x 108
3 x 108
1 x 10s
50 B/s : 1 CR/s B/s only
20 B/s : 1 CR/s
50 B/s : 1 CR/s
3 B/s : 1 CR/s 5 B/s : 1 CR/s 5 B/s : 1 CR/s
3 B/s : 1 CR/s
B B B B 10B/s : 1 CR/s
50B/s : 1 C R / s
1.5 x 109 50 K ( h ) / s : 1 B/s A,C
1.5 x 109 12 K ( h h ) / s : 1 CR/s B,C
B,C
1.0 x 109 25 K ( X ) / s : 1 B/s
5.0 x 10s 8 K ( A h ) / s : 1 CR/s B,C
2.0pg acriflavine per ml bottom layer agar with CR in top layer
(useful only in absence of a m mutants) BR/s bacteria used
* All Method-I tests are made on a 2 ml top layer of soft agar which is seeded with 0.1 ml cf phage suspension (where appropriate) and 0.15 m l of bacterial mixture. Except for ac41, 0.5 mg of streptomycin sulfate is added to the top layer of each test plate. For Method I1 (de- scribed later) all components of the top layer are increased by a factor of 2.5.
+
Most of the a m mutants were obtained from R. S. EDGAR and R. H. EPSTEIN. The numbers in parentheses are the gene numbers assigned by EDGAR, DENHARDT and EPSTEIN (1964). All mutants with the letter S preceding the mutation number were isolated in the present program from phage grown in the presence of bromodeoxyuridine.2 Volumetric ratio of overnight aerated cultures
$ All tests listed are satisfactory when no other mutant in the same cistron is to be distinguished. Quality symbols give the following additional information:
A = Good tests even with other mutants in the same cistron
B = Tests difficult although possible to distinguish when another mutant is included
C = Improved and generally satisfactory with 36-48 hour incubation of test plates. in the same cistron
crossovers is anticipated when the phage being tested carries r77+ surrounded by the closely linked markers r70 on one side and rb42 on the other, and when the test-plate is seeded with 7-77 (for map, see DOERMANN and BOEHNER 1963). Of these markers, rb42 is located in the rZZB cistron while the other 2 are located in
the rZZA cistron. It is possible, therefore, that functionally complementary com-
424 A. H. D O E R M A N N A N D L. B O E H N E R
the latter). An example of such tests is shown in Figure 1. All tests were con- clusive in that easily recognizable clearings were found for each plaque tested. Closer combinations of markers would, of course, necessitate new controls.
Similar tests with a m mutants using condkions given in Table 2 yielded satis- factory results. It seems, however, to be more difficult to resolve three amber mutations in a single cistron than it does to distinguish three or more rZZA or
rZZB mutations.
Errors due to false positive tests resulting from reverse mutations during plaque growth: Genetic reversions might give rise to a second source of error. If, for example, reversions occur frequently during growth of an
rZZ
plaque, the revert- ants would make a clear spot, implying an r+ genotype at the site being tested. The use of copy plates, with an additional cycle of growth, would probably tend to accumulate revertants and perhaps exaggerate this source of error.Six rZZ and 15 a m mutants were checked for the occurrence of such false posi-
tives when tests are made with Method I. Stocks of each were plated against CR- 63, and a set of 234 plaques of each copied on CR-63. Tests of the copied plaques were made by the standard procedure, seeding the test plates with the homologous mutant. Of the rZZ’s, three (r61, r64, and r73) have no detectable reversion fre- quency, and, as expected, no errors were found in the 702 plaques tested. Stocks of the other three (r70, r77, and rB32) always contain revertants in low fre- quency. However, only one test (among the rb42 set) was found to be misleading out of the 702 tests made. It seems reasonable to ignore any error from this source when these particular rZZ mutants are used.
Each of 15 a m mutants investigated similarly showed reversions in stock suspensions. Seven (amB16, amN52, amN54, amN58, amN82, amN85, and
amS71) yielded no test error, and four (amB6, amS6, amS52, and amSlO5) gave
one doubtful test each. Three others (amN131, amS29, and amS60) produced one positive test each. The remaining mutant (amS92) yielded three false posi- tives in the 234 plaques tested. The frequency of misclassification of these ambers due to revertants seems negligible, with the possible exception of a d 9 2 tests.
I t should be called to attention here that any phage which is heterozygous a t a given site segregates the wild type during the growth of its plaque. Heterozygous sites are therefore misclassified as wild type both i n Method I and in Method 11. For measurement of large recombination values this error can be neglected, but when closely linked markers are being studied the proportionate error becomes large. A more detailed and quantitative discussion of this complication is reserved for a subsequent report.
Method
ZI:
Testing large numbers with a replicating techniqueReplicating test procedure: Method I is practically feasible as long as the num- ber of markers per cross is not too large. I t has been extensively used in 7-factor crosses by DOERMANN and BOEHNER (1963) and in a 9-factor cross by WOMACK
I D E N T I F I C A T I O N O F G E N O T Y P E S IN P H A G E T 4 425
principle of Method I1 is the same as for Method I, the modifications require that it be described and tested separately.
The velvet replicators used by CHASE and DOERMANN (1958) are not depend- able enough for $he present type of experiment because they tend to pick up large areas of the soft-agar #top-layer, making repeated replications from one master plate impossible. Metal replicators were therefore constructed using No. 18 gauge spring-temper stainless steel wire mounted in a babbict metal base, leaving a uni- form length of about 1.27 cm protruding. The wires were originally spaced 4.33 to the cm in rows 0.196 cm apart. This high density of inoculating wires pre- vented complete burning away of the alcohol used to sterilize the replicators, so that it was necessary to remove every even-numbered wire in alternate rows.
Copy plates are essential intermediates when using replicators of this type, especially when crosses include numerous amber mutants which bring about considerable plaque-size variation. In order to make the number of tests per repli- cation as large as practical, Petri dishes of 150 mm diameter are used for copy and test plates. Both are prepared as in Method I, except that the soft-agar layer is increased to 5 in1 and the other components of the test (bacteria, indicator phage, streptomycin, etc.) are increased by a factor of 2.5.
Replicators are sterilized by standing them for
5
sec or longer in 3-5 mm of95% ethyl alcohol which is then burned off. A little time must be allowed for cooling and it becomes expedient to have a set of 6-12 replicators if many tests are to be made. After a number of cycles of replication and sterilization it is use- ful to brush the replicators with a wire brush to remove adhering bits of soft agar which tend to accumulate.
Optimum tests appear to be obtained when test plates are not stored in the refrigerator but used directly after the soft agar has hardened. The lids are re- moved 5-10 min ahead of the replicating process to allow evaporation of excess moisture. It is advisable to finish replication as soon as possible after pouring the soft agar on the test plates, but even when delayed as long as three hours, satis- factory tests are generally experienced. If it becomes necessary to refrigerate the test plates, they should be removed from the refrigerator and the lids removed for 10-20 min ahead of the replicating process. After replicating they are left open 10-20 min longer, depending on atmospheric conditions.
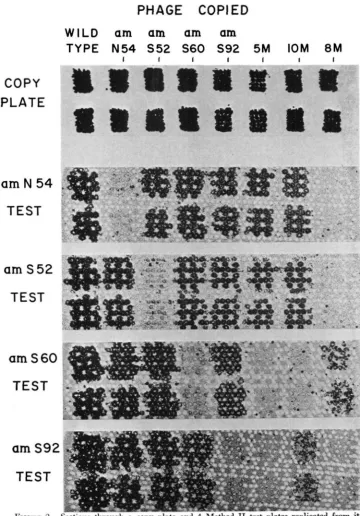
Figure 2 shows a section of a copy plate and
4
test plates made by Method 11. Controls against misidentifications: The replicating procedure was checked for the same two sources of error as were investigated with Method I. Plaques of the double mutants r70 rb42 and r64 r77 were copied and replicated to test plates containing r77 and r70, respectively. In the total of 700 tests none failed to show a definite clear spot, indicating that under proper test conditions the double cross- over is frequent enough to permit recognition of the centrally located r+ with a very high degree of reliability.PHAGE COPIED
W I L D
am am
am am
TYPE N 5 4 S 5 2 S 6 0
S 9 2
5M
IOM
8 M
I I I I I I I
COPY
PLATE
am
N 54
TEST
am
S52
TEST
a m S 6 0
TEST
am
S 9 i
TEST
FIGURE 2.-Sections through n copy plate and 4 Method I1 t e s t plates replicated from i t
1.2 x. Two plaques of 8 different genotypes were copied and tested. The designation 5M indi- cates phage carrying the 5 mutant genes amN85, amS71, amS60, amS92, and amSlO5. Phage
I D E N T I F I C A T I O N O F G E N O T Y P E S I N P H A G E T4
TABLE 3
427
Frequency of false positive tests in mutant identification b y Method I 1
Three hundred forty-fiue plaques from phages carrying a single mutant were copied on CR-63 and replicated to test plates seeded with the homologous phage
~~
List of mutants tested
~
Errors' in 345 tests
amNl35 a d 1 6 amB23 amN53 amB17 amE355 amS52 amNI31 a d 1 0 5
amB256 a d 9 2
amB251
amS6O amB6 amN85 amS6 amN54 amN58 amN52 r73 rb45
a d 7 1 rb42
amS29
rb41 rED220 r70 r71 r64 r61
rED219
amNI30
amN81 amN82
ac41
0
1
2
*Whenever doubt existed about how a test should be scored, it was classified as a n error. Additional evidence exists which indicates that when tests are made on plaques from multi- mutant phages, false positives are even less frequent than indicated by tests of single mutants.
to cause concern, particularly in view of the fact that tests from multiple-mutant stocks (which are generally involved in crosses which employ replication) show a lower frequency of false positives than do the single-mutant tests on which the table is based.
Multi-amber phages often make very small plaques which may give rather sparsely cleared areas on the copy plate. To obtain dependable tests it is some- time? necessary to recopy the original copy in order to produce a more or less completely cleared area. This raised the question whether repeated copying ac- cumulates revertants which might give misleading tests. As a check against this possibility plaques from a phage stock marked with one rZZA and 21 a m muta- tions were copied and tested for each marker. (The markers were r219, amB256, amN135, amB251, amBl6, amB23, amB17, amS52, amS105, amS29, amS92, amS71,amB6, amN85, amN54, amB25, amA455, amN52, amS2, amN81, amN82,
and amNl30.) After the first test, the copy spots were recopied on a fresh plate
and again tested for each marker. Among 210 plaques handled in that way, none showed a nonmutant test for any of the markers in the test from the original copy. The tests of the second copy plate showed one nonmutant test (for amB251). When 140 of the second copies were again recopied, four reversions (all amB251) were found. Thus, for the markers tested, a single recopying of small plaques does not appear to be hazardous.
428 -4. H. D O E R M A N N A N D L. B O E H N E R
S U M M A R Y
Two methods have been developed for determining the genotypes of individual phages derived from complex crosses. Method 11, which makes use of replicators especially designed for use with soft-agar overlays, has two main advantages: (1) when many markers are being tested in one phage, the process becomes much more efficient when replication is employed; (2) the orientation of the plaques is the same on all test plates, thus eliminating accidental transpositions and other sorts of technical errors. On the other hand, Method I is preferred when the num- ber of tests per phage is only 2 or 3 because it is then unnecessary to interpolate a copy plate between the original plaque and the final test. Both methods seem to yield equally reliable results when mutants with low reversion indices are selected and if the number of markers in one cistron is not too great.
L I T E R A T U R E CITED
BENZER, S., 1957
CHASE, M. C. and A. H. DOERMANN, 1958
DOERMANN, A. H. and L. BOEHNER, 1963
DOERMANN, A. H. and M. B. HILL, 1953
EDGAR, R. S., 1958
The elementary units of heredity. pp. 70-93. In: The Chemical Basis of Heredity. Edited by W. D. MCELROY and B. GLASS. Johns Hopkins Press, Baltimore.
High negative interference over short segments of the genetic structure of bacteriophage T4. Genetics 43 : 33S-353.
An experimental analysis of bacteriophage T 4 hetero- zygotes. I. Mottled plaques from crosses involving six rll loci. Virology 21 : 551-567.
Genetic structure of bacteriophage T4 as described by recombination studies of factors influencing plaque morphology. Genetics 38: 79L90.
Mapping experiments with r l l and h mutants of bacteriophage T4D. Virology 6: 215-225.
EDGAR, R. S., G. H. DENHARDT and R. H. EPSTEIN, 1964 A comparative genetic study of con- ditional lethal mutations of bacteriophage T4D. Genetics 49 : 635-648.
EDGAR, R. S. and R. H. EPSTEIN, 1961 Inactivation by ultraviolet light of an acridine-sensitive gene function in phage T4D. Science 134: 327-328.
EDGAR, R. S., R. P. FEYNMAN, S. KLEIN, I. LIELAUSIS and C. M. STEINBERG, 1962 Mapping experiments with r mutants of bacteriophage T4D. Genetics 47: 179-186.
EDGAR. R. S. and I. LIELAUSIS, 1964 Temperature-sensitive mutants of bacteriophage T4D: Their isolation and genetic characterization. Genetics 49 : 619-662.
EPSTEIN, R. H., 1958 A study of multiplicity-reactivation in the bacteriophage T4. I. Genetic and functional analysis of T4D-K12(X) complexes. Virology 6: 382-404.
EPSTEIN, R. H.. A. BOLLE, C. M. STEINBERG, E. KELLENBERGER, E. BOY DE LA TOUR, R. CHEVALLY,
R. S. EDGAR, M. SUSMAN, G. H. DENHARDT and I. LIELAUSIS, 1963 Physiological studies of conditional lethal mutations of bacteriophage T4D. Cold Spring Harbor Symp. Quant. Biol. 28: 375-392.
Foss, H. M. and F. W. STAHL, 1963 Circularity of the genetic map of bacteriophage T4. Genetics 48: 1659-1672.
Acridine-resistance in phage T4D. Genetics 56:
Sur le determinisme gGn6tique de la lysogenie. Ann. Inst. Pasteur 84:
An analysis of single-burst progeny of bacteria singly infected with a PIECHOWSKI, M. M. and M. SUSMAN, 1967
133-148.
WOLLMAN, E. L., 1953 281-293.
WOMACK, F. C., 1963