I N T H E DEVELOPMENT OF THE MOUSE C. H. DANFORTH AND S. B. DE ABERLE Department of Anatomy, Stanford University, Cdifwnia
Received December 17, 1926
TABLE OF CONTENTS
PAGE
INTRODUCTION. ... 340
The Ay gene in mice.. ... 341
The W gene in mice. ... 344
DISCUSSION.. ... 345
CONCLUSIONS. ... 347
LITERATURE CITED.. ... 347
INTRODUCTION
While i t is obvious that somatic traits are largely determined by genes in the chromosomes, the precise way in which the genes produce their effect is as yet unknown. Two possibilities suggest themselves. Genes located in the same nucleus may mutually react with each other in such a way as to give a distinctive resultant effect which will produce charac- teristic somatic manifestations, or each gene may severally and inde- pendently exert its own peculiar influence upon the surrounding cyto- plasm.
In the first case the relation between genes and soma might be indicated
as
in which A and B represent genes, N the cytoplasm of a cell, and S,
S1, or
S2,
its possible contributions to the soma. Whether the particular contribution of the cell shall be S, S1, S2-
.
will depend on its topographi- cal position and such other factors as are studied by embryologists and experimental morphologists, but the essential character of the nuclear influence will be fixed throughout by the nature of the interaction between A and B.A " n
5 2
The second possibility may be indicated as
B-'<&
in which n and n' represent independent reactions of A and B with the cytoplasm and stand for two units in the sum total of cytoplasmic po- tentialities. I n this case A and B might both contribute to the deter-
INTER-RELATION OF GENES IN MOUSE 341
mination of S, while
B
would have no‘ part in the determination of SI, and A none with reference to S2. According to the first supposition ifB
modifies the effect of A on one trait it should also modify its effect on any other trait, even with full allowance for cytoplasmic differentiation and the action of environmental factors; while according to the second,B
might influence the effect of A on some traits but not on others.A figurative statement of the problem is suggested by an analogy em- ployed by CONKLIN (1920, p. 130) in which it is suggested that the genes “represent the differential causes of particular characters just as in the compounds H2SOa and K2S04 the hydrogen and potassium atoms represent the differential causes of the properties manifested by these two sub- stances.” Continuing the analogy essentially in this form, one might picture the nuclei of one idividual as “H2SO4” in their reaction with the cytoplasm a t successive developmental stages, while the nuclei of another individual would react as “K2S04.” This accords with the first alter- native, where the difference is not simply between “H2” and “KZ”, but between the compounds “H2S04” and “K2S04.” On the second assumption the analogy of a chemical compound breaks down. I n place of the com- pounds are the elements “H”or “K”,“S,” and“0,” each tending to formits own independent reactions with the cytoplasm. Either arrangement would seem to provide the possibility of a great many “unit characters,” but the implications of the two hypotheses are quite different.
An approach to the solution of this problem is afforded by cases in which the same primary gene produces two or more distinct effects, in connection with a t least one of which some other known gene is also operative. A y and W in the mouse and the genes which affect their
expression, present situations of this sort.
THE A U GENE IN MICE
The same gene that determines yellow coat color also results in an increased adiposity after maturity. (DANFORTH, 1926). These traits occur when the gene is in the heterozygous condition. Individuals homo- zygous for it are apparently unable to survive the early embryonic period
(KIRKHAM 1917, IBSEN and STEIGLEDER 1917), so this gene may also be regarded as a lethal. Thus three very different effects of the A gene may be recognized: early death for homozygous individuals, yellow color and adiposity for the surviving heterozygotes. But as regards color, it is well known that anumber of other genes are effective in modifying or suppressing yellow in the pelage. Among these are the genes for dominant
( W w )
and recessive (ss) spotting and that for albinism (cc).Now, since a mouse with the genetic formula CCSSAua has a yellow pelage and becomes obese, what will be the character of a mouse with some modification of this formula such as CCssAua which has only a limited amount of yellow, or c c S S A ~ a which has no yellow at all? If the genes S and A v, and c and A . interact directly with each other and in-
directly with the cytoplasm it might be expected that the spotted mouse would show only a limited tendency to adiposity and the albino none a t all; but if they do not interact directly the effect of thesegenesonadiposity could not be predicted from their effect on color.
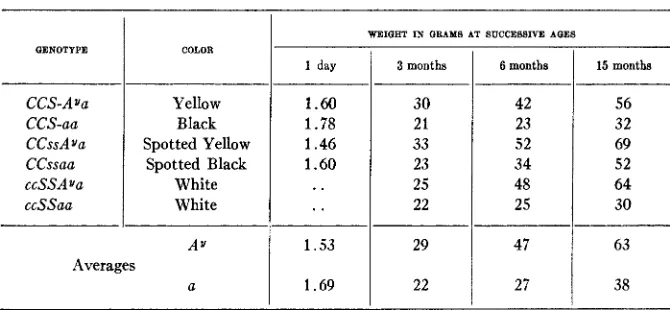
Strains with various formulae have been made up and tested for possible relations between color and adiposity. Table 1 is an abstract of a series of data on the weights of three pairs of litter-mate sisters. The two members of each pair were kept in the same cage, supplied with the same
TABLE 1
Weights of mice of dijerent genotypes with reference to the presence or absence of Ay.
I
I
TVEIQHT IN QR.418 AT SUCCEBBIVE AQE8 QENOTYPE COLORCCS-Aua
White
ccSSaa
White
ccSSAua
Spotted Black CCssaa Spotted Yellow CCssAva Black CCS-aa Yellow AV Averages a
1 day
1.60 1.78 1.46 1.60 . . 3 months 30 21 33 23 25 22 29 1.69
6 months
42 23 52 34 4s 25
15 month
56 32 69 52 64 30 63 27
food, and in every respect given as nearly identical treatment as possible. I n these three sets the Ay gene is brought into relation with a combination which permits a full expression of its effect on color, one which restricts its expression, and one which wholly suppresses it. I n each case, however, the individual carrying the A . gene becomes much heavier than the cor- responding control which lacks it. Similar results have been obtained in other controlled experiments, and have been confirmed no less con- vincingly by the repeated occurrence of typical adiposity in dominant spotted, r e d v e s p t t e d , ‘‘sooty” and albino individuals which were known to carry A., but which had not been especially bred to test the point in question. These observations make it clear that the ability of
the A y gene to produce adiposity is not related to its opportunity to affect
INTER-RELATION OF GENES IN MOUSE 343
The other well known action of the AV gene is its lethal effect when tomozygous. Since no homozygous strain of ordinary yellow, sable, hnk-eyed, or spotted yellow has ever been obtained, the combined ex- perience of many investigators may be taken as evidence that the genes which thus limit or modify the effect of A y do not prevent the lethal action
of A y A y . It remains, however to determine whether or not albinism
which causes a complete failure of the A y gene as a color factor also
suppresses it as a lethal. This point was tested by matings between indi- viduals with the formula c c A q which should produce some zygotes of the constitution c c A ~ A y . The stock was obtained from an original
mating of an albino female and a yellow male. The first generation con- sisted of some yellow and some black offspring. The latter were discarded and the yellows mated among themselves, giving in Fz some albino and some colored young. The albinos were saved and tested by mating to non-yellow colored individuals. Those that produced any yellow young were shown by that fact to be of the formula CCAU These were now mated together and their offspring, F1, raised to maturity and tested as to their genetic constitution.
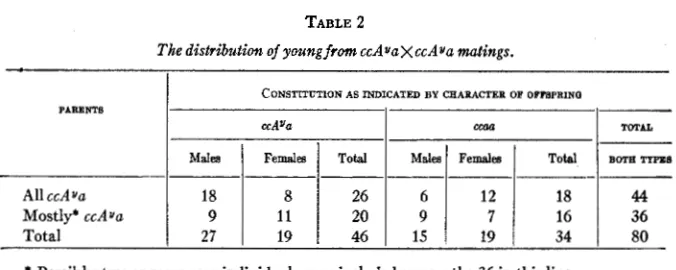
In young from the mating c c A ~ a X c c A ~ a the effect of A on color is wholly suppressed. If its lethal effect were also suppressed, one-fourth of the young should have the formula c c A ~ A u and produce only yellow young when bred to CCaa mates. The result of the tests is shown in table 2. Forty-four mice from c c A ~ a ~ c c A ~ a parents were found to include no
ccA y A individual. If ccA Y A Y were not lethal there should have been
about eleven of this formula. Of these forty-four, 26 were found to be Aua and 18 recorded as aa. Some of the latter, however, were possibly Aya since each individual was classed as not A u A y and discarded as soon as it produced a non-yellow colored young. Thus seven mice were classified as not AuAv and probably aa on the basis of litters of five or less young. This may account for the slight deviation from the ratio of 29: 15 which might have been expected on the assumption that AuAu is really lethal. If ccA AY were not lethal, the chances against this combination occurring a t least once among 44 mice of this ancestry are (3/4)44, or among 26 yellow producers (2/3)26. The odds against the failure of a ccAuAu individual to occur being a matter of chance are thus more than 30,000: 1.
The data on 36 mice among which two or more ccaa individuals were inadvertently included are also given in table 2. This shows a total of 80 mice, the majority of which were suitable for critical tests, and all of which were shown by the 765 young which they produced not to be ccAyAU. It is consequently apparent that the combination cc which
suppresses the effect of A V on color does not prevent it from being a lethal when homozygous.
TABLE 2
The distribution of young from ccAuaXccAua matings.
PAHENTB
I
CONSTITUTION AS INDICATED BY CHARACTER OF OFFWRING
I
ccAual
mI
Males1
FemalesI
TotalI
MalesI
FemalesI
TotalAll ccAVa 18 8 26 12 18
Mostly' ccAua
1
2y
1
ii
1
iz
1
18
1
1
16Total 34
-
TOTAL
BDTH TYPES
44
36 80
'
Possibly two or more ccaa individuals were induded among the 36 in this line. THE W GENE I N MICESomewhat similar tests were made with mice carrying the gene ( W )
for dominant spotting. When heterozygous this gene gives rise to a certain amount of white in the pelage; when homozygous, i t produces a severe anaemia which terminates fatally within a few days of birth.
(DE ABERLE 1925.)
The gene W may be introduced into a number of combinations, some of which have been discussed by DETLEFSEN (1916) and others by
LITTLE (1917). The combinations CCPPSSAUaWw, CCPPSSaaWw,
TABLE 3
The production of anaemia by mice in which different genes were associated with W .
QENOTYPE OF THE PARENTS NORMAL YOUNQ ANAEMIC TOUNQ PERCENTAQE OF ANAEMICS'
-
C-P-ssaeWw (Black-eyed whites)
13.6
77 487
Totals
15.2 29
161 ccPPS-aWw (Albinos)
11.9 20
147 CCPPSSaaWw (Roans)
13 .S
28 179
'
The numbers in this column fall short of the expected 25 percent because some of the anaemics are born dead and others are lost between the inspections which in each case occurred at 24-hour intervals.CCPPssAuaWw, CCPPssaaWw, CCppssaaWw, and ccPPSSAuaWw give
a graded series ranging from animals that are almost completely pig- mented to those with no pigment at all. In each of the complexes except the last, W is a factor in the production of the characteristic grade. Or, considering W as the constant, its effect on color is modified in one way
INTER-RELATION OF GENES IN MOUSE 345
or another by each of these combinations. The
W
gene has been tested in each of the six combinations just mentioned (except that some of the individuals used were heterozygous for C,P
or S). But none of the tests showed any marked effect on the incidence of anaemia (table 3). The relatively low frequency of anaemia in all tests is due to the poor viability of the anaemics and the fact that the cages were inspected only at 24 hour intervals. All classes produced anaemics in about the same proportion, and each of them produced perfectly typical representatives, those from the two extremes of the series, albinos and yellows, being quite as similar as those from any other two groups. This makes it certain that the genes which influence W with respect to color do not influenceWW in any comparable way in respect to anaemia.
DISCUSSION
Since genes which modify or suppress the effect of A” on color show no corresponding effect on its lethal action or its tendency to produce adi- posity, and those which similarly influence W with respect to color do not affect the anaemia, it would seem that the second of the two suggested alternatives is the more nearly correct; that is, that genes which modify each other’s effects do so indirectly and not through direct mutual inter- action. If this conclusion is correct, and of general applicability, a com- monly expressed view as to the interaction of genes is not tenable. In this case it is not strictly true that development consists in the “orderly interaction of these substances (genes)-with each other, with the rest of the cell body,or cytoplasm; and with the oxygen,food,
.
.”
( JENNINGS, 1924). The “with each other” should be deleted. If the seeming implica- tions of the data are incorrect, it must be that we have been unfortunate in selecting examples where traits produced by the interaction of A-Nand AB-N are indistinguishable while A-NI and
AB-N1
are widelydifferent. ( N and N I , representing the cytoplasm a t two different stages of development). However improbable, this is not entirely impossible.
A point of interest in this connection is the behavior of multiple allelo- morphs. In Drosophila according to MORGAN (1926, p. 307) changes in the same locus affect all the traits which were affected by the original gene. This would seem to be confirming evidence from an entirely different angle. Instead of keeping one gene constant and varying its modifiers, the general complex is kept constant and the particular gene is varied. Never- theless, the data presented here do not indicate that the relation mentioned by
MORGAN
is a necessary one and indeed evidence, which is not as yet adequate, suggests that it may not hold with the A , A * , Aw, a series of multiple allelomorphs in the mouse.The uncertainty in the situation arises from the fact that it is not entirely clear whether or not each mutant gene actually contributes to all of the reactions of the original or if some of the observed effects are due to the failure of the mutant to act at all in certain capacities. It is not surprising, however, that allelomorphic genes are frequently observed to affect the same series of traits, for if in the development of a normal individual a certain gene plays a necessary rBle in several vital reactions any mutant gene which replaces it must be able to somehow function in each of them or else behave as a lethal. Failure of mutant genes to meet this requirement may account for most lethals, and the likelihood of a viable mutation is probably quite as dependent on the’number of “secondary” reactions to which the gene must contribute as upon the importance of the “primary” trait. It would seem that, other things being equal, viable mutations should most frequently be due to changes in genes which participate in a minimum number of reactions during de- velopment, and that they should most often affect “superficial” characters. But as yet there seems to be no real information as to just how a mutant gene fits into the place of the original.
Two possible objections to the procedure in this paper will possibly suggest themselves. I t may seem that the idea of modifying genes is employed in an unjustifiable sense, that spotting, for example, does not affect yellow but color distribution and yellow merely happens to be the color in question. But wholly apart from the question of whether or not there are modifying genes in any other sense, it may be recalled that in the presence of W , yellow shows a very different degree of extension than does black or brown (LITTLE, 1917). Moreover, the question a t issue really is this: Given a certain genetic constitution, does a new gene introduced into the stock by breeding produce its effect by reacting directly with the other genes introduced into the zygote a t t h e same time or by reacting indirectly through the cytoplasm later on? For throwing light on this question the data under consideration would seem to be pertinent.
INTER-RELATION OF GENES I N MOUSE 347
attribute to two genes, A . and W , are in reality due to five different genes, the data and discussion in this paper would have very little point.
CONCLUSIONS
Several genes which influence the effect of A @ on color do not influence its effect on adiposity, or its lethal action when homozygous.
Similarly, combinations which influence the effect of W on color have no affect on anaemia.
These facts indicate that there is no constant interaction between the genes themselves, but that their somatic effects are produced by inde- pendent reactions with some other substance.
LITERATURE CITED
DE ABERLE, S. B., 1925 Hereditary anaemia and its relation to dominant spotting in mice. CONKLIN, EDWIN G., 1920 Heredity and environment in the development of men. xv+361 pp.
DANFORTE, C. H: 1925 Adiposity and doubling as constitutional traits in the mouse. Anat. Rec.
1926 The interaction of genes in development. Proc. Soc. Exp. Biol. and Med. 26: 69-71. DETLEFSEN, J. A., 1916 Pink-eyed white mice carrying the color factor. Amer. Nat. 50: 4649. IBSEN, HERMAN I., and STEIGLEDER, EMIL, 1917 Death in utero of the homozygous yellow
JENMNGS, H. S., 1924 Heredity and environment. Scientific Monthly 19: 225-238.
KIRKHAY, W. B., 1917 Embryology of the yellow mouse. Anat. Rec. 11: 480481.
LITTLE, C. C., 1917 The relation of yellow coat color and black-eyed white spotting in mice.
MORGAN, T. H., 1926 The theory of the gene. xvi+343 pp. New Haven: Yale University Amer. Nat. 29: 327-335.
Princeton: Princeton University Press.
29: 354,
mouse. Amer. Nat. 51: 740-752.
Genetics 2: 433-445.
Press.