THE RELATION BETWEEN FITNESS COMPONENTS AND POPULATION PREDICTION I N DROSOPHILA. I:
THE ESTIMATION OF FITNESS COMPONENTS
TIMOTHY PROUT
Department of Life Sciences, University of California, Riuerside 92502
Received October 22, 1970
H E estimation of fitness components in Drosophila and other organisms is usually a n extremely laborious procedure which is commonly done under conditions remote from those of the population and is often incomplete in that some potentially important components remain unstudied.
These difficulties appear especially in connection with studying the adult phase of the life cycle, and it is precisely this phase that can generate complex modes of selection due to differential fertility and mating behavior. Thus the problem of estimating adult components is an important one in the general study of pat- terns of selection.
In order to circumvent the difficulties of the usual methods of component studies, KNIGHT and ROBERTSON (1957) devised a method of utilizing a special marker stock which provided an estimate of adult and larval components, as well as a combination of these components similar to net fitness. WRIGHT and KERR
(1 954) ? incidental to a study of small populations were able to exploit their data to yield estimates of male and female “productivity.” SPOFFORD ( 1 956) also esti- mated adult “productivity” of homozygotes for the recessive gene eyeless as com- pared to heterozygotes. This paper suggests another experimental procedure for estimating certain broad fitness components in the adult phase under competitive conditions close to those of a population. The method does not employ a marker stock as did KNIGHT and ROBERTSON, but is basically a n extension of the methods of WRIGHT and KERR and of SPOFFORD.
I n what follows the experimental system together with its theory will be set forth first, then the system will be illustrated using certain Drosophila genotypes. In an accompanying article (PROUT 1971) the fitness components estimated here are applied to certain population experiments.
THE EXPERIMENTAL SYSTEM: THEORY
Basically the procedure involves placing together contrived numerical mix- tures of genotypes as adults and then recording the proportions of the genotypes among the resulting offspring. The fitness estimates are simply the values with which the known proportions among the parents must be weighted in order to
account for their offspring.
128 T. PROUT
discrete generations and a short period of mating and egg laying. The genetic system under consideration constitutes the three genotypes produced by two alleles segregating at one autosomal locus.
Ecologically and genetically this is a very simple system. Even so, the events which may affect gene frequencies can involve considerable complexities worth studying.
It is convenient to resolve net fitness into larval and adult components for the sexes separately. The symbol L will be used for the larval components. The adult components represent very different biological activities for the two sexes. In the case of the females there will be some differential death during the short span of their adult life, but principally the female component will involve differential fecundity, hence the female adult component will be designated by F . On the other hand the adult component for males, although somewhat influenced by dif- ferential death, will principally involve differential mating activity and so will be designated V for virility.
I n addition to recognizing separate male and female larval and adult com- ponents, there is another factor which can have profound effects on gene fre- quencies-this is mating interaction. Mating interaction will be here defined as the circumstance where virility of a particular male genotype varies as a function of the particular female genotype which he encounters.
In order to facilitate the description of how these components are to be esti- mated, first it will be assumed there are no mating interactions.
For three genotypes ( A A , Aa, and aa) with a larval and adult component for each sex there will be six parameters to estimate, assuming one of the phenotypes has fitness component of unity. These six parameters are set forth below. where the index
i
denotes female genotypes as shown, and j denotes male genotypes.Females Males
Genotypes A A Aa aa A A Aa aa
I = 1 2 3 j = 1 2 3
Larval component L, 1 L, L l , 1 LQ
Adult F , 1 F , Vl 1 V ,
The larval component of fitness is the usual viability. The estimation of this component is straightforward and involves simply determining the values of L necessary to account for departures from Mendelian ratios in experimental crosses producing Fz’s or backcross progenies.
It is for the estimation of the adult components that the method of contrived mixtures is used. The male and female components are estimated in separate ex- periments. Newly emerged flies are used and a mixture of the three genotypes
of one sex is counted out and mated to a population of the other sex composed
of j u s t one genotype. Thus a mixture of female genotypes mated to one type of
ESTIMATING FITNESS COMPONENTS 129
counted can all be adjusted by the experimenter. Nevertheless, unlike the usual male- or female-choice experiment, the offspring of the mixture are raised en
masse and are classified and counted as newly emerged adults at the same stage
of development as their parents when the experiment started.
In short, the system amounts to one generation of selection where only one sex is genetically variable, and the pattern of this parental variation is determined by the experimenter.
The estimation procedure will be illustrated with a fecundity cross, in which the female population is a mixture of genotypes and the male population is com- posed of just heterozygotes. The succession of events is diagramed in Figure 1.
At the upper left the contrived proportions in the mixture are denoted by
Ql:
Q 2 : Q 1 . This initial mixture is then weighted by the fecundity parameters, F,:1 : F, to account for the differential contribution of these genotypes to the off- spring population. The gametic output is then calculated from the weighted mix- ture. It appears as though only differential death is being taken into account since the gametes are computed after the weighting is done. However, this is alge- braically equivalent to having random mating first and then differential egg laying in accordance with the weights Fl:l:F,. Thus the weights include both processes of death and egg laying. Next, random union of the gametes produces the genotypic proportions at fertilization. These proportions are then subject to weighting by the larval components of fitness, L,: 1 : L, to finally give the geno-
typic proportions among the newly emerged adults. For the purpose of this dem- onstration it is assumed that the larval components are sex independent. These proportions, which are observed by the experimenter, are denoted by Q1,
Qs,
andQs,
whose expectations are written out as equations ( 1 ) at the bottom of the figure.These equations, then. constitute the model. The observations for a n experi- ment constitute two data sets: The proportions in the contrived mixtures of par- ents, Q1:Q2:Q9, which are known precisely, and the proportions among their progeny,
Q1:
Qd:
Q,,
which are subject to sampling error.The statistical problems of both estimation and error will be pursued later in connection with the experimental data. It is sufficient to note here that in the experiment just described there are four fitness component parameters to be esti- mated: the two larval components L, and L,, and two adult components F , and F,. This number can be reduced to just the two adult components if the larval components are already accurately known as a result of independent experi- ments using backcross progenies or F,’s.
Table 1 schematically represents a generalization of this experimental system. There are six sets of experiments shown: three sets of fecundity experiments on the left, and three sets of virility experiments on the right. These three sets differ from one another with regard to the genotype of the constant parent to which the contrived mixture is mated; the constant parent may be one of the two homo- zygous genotypes, or it may be heterozygous as it is in the case just discussed and
130 T. PROUT
G e n o t y p e AA A a aa AA A a aa
1 2 3
i - 1 2 3 j =
Q1 Q2 93
Mixture at
S t a r t
0 1 0
SELECTION:
F F F
GENOTYPIC PROPORTIONS AT FERTILIZATION
AFTER LARVAL SELECTION
FIGURE 1.-Model used for estimation of fitness Components. This case, used for illustration, assumes a heterozygous constant male parent and variable female parents. Q i genotype pro- portions in contrived mixture;
Gi
=
genotype proportions among progeny; F i=
female adult fitness component parameters or fecundity; Li E larval survival fitness component or viability.any number of different mixtures may be used ( Q l k , Q s k , Q g k where
k
= I , 2, 3 for different mixtures of femalesi
and similarly P j k for male mixtures), thus al-ESTIMATING F I T N E S S C O M P O N E N T S
TABLE 1
Summary of the various experiments which could be p e r f o r d Qik E female genotype proportions in the kth mixture; Pjlc e male genotype
proportions in the kth mixture.
131
Qll
Qlk
Qll
Q1k
Qll
'1k
VIRILITY
FECUNDITY
--
CONSTAXT dJ CONSTMT 99
PARnXS
99 MIXTURE PARENTS
~-
a xxTurs--
921
Q2k
921
Q2k
Q21 Q2k
'31
'3k
'31
'3k
Q31
'3k
'lk
Aa PI1
' l k
p11
aa
'lk
p 2 1
'2k
p21 '2k
p21
'2k
M '31
'3k
'31
'3k
'31
'3k
Aa
aa
APPLICATION O F E X P E R I M E N T A L S Y S T E M
MATERIALS A N D M E T H O D S
(1) Materials and methods: The experiments reported below were designed to test the feasi- bility of the procedure. However, frequency dependence of fitness was not investigated. The results of investigating this factor will appear in a later report.
The genetic system chosen involved the two fourth chromosome recessive mutants of D. melanogaster, eyeless (ey2) and shaven naked (sun) in repulsion linkage phase. Due to the negligible recombination in this chromosome, these mutants behave in populations as alleles. This genetic system was chosen for four reasons:
(1) The two recessives complement to produce a distinguishable wild-type heterozygote. (2) Being laboratory mutants their effects are sufficiently drastic so that most fitness com- ponents are likely t o be affected.
(3) The use of the small fourth chromosome permits the quick randomization of the residual genotype.
(4) Because of the wild-type phenotype of the heterozygote, the system will produce over- dominant population dynamics. This allows the establishment of a study population which will segregate indefinitely.
The two mutant strains were obtained from the California Institute of Technology, and the F, resulting from a cross between them was used as the foundation stock for the experiments. First, five replicated populations were established from the foundation stock. Each population was carried in discrete generations in half-pint bottles with a standard agar-sugar-yeast medium. The cultures were kept at 25°C. Further details of the population regime will be found in the accompanying article (PROUT 1971 ) .
132 T. PROUT
started. Each contrived mixture was made up in a culture bottle with a total of 60 flies of each sex, where one sex was composed of just one phenotype and the other sex composed of a known mixture of phenotypes. Two day old males and virgin females were used, they were allowed to mate, and the females laid eggs for three days. Then the flies were transferred to a new bottle and allowed to lay additional eggs for just 12 hr. These latter cultures were kept at 25°C for 13 days at which time all the flies had emerged. Then a sample of about 1009 0 and 1006 6 was classified and counted.
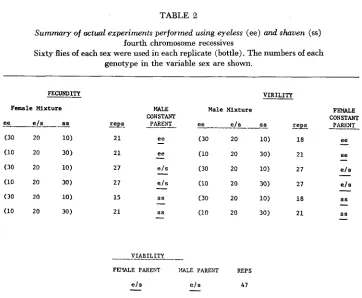
Table 2 summarizes the actual experiments performed. A unit of replication is a single culture bottle treated as just described. The symbol ee will represent the eyeless phenotype which is wild type for shaven. The symbol ss will represent the shaven phenotype which is wild type for eyeless. The symbol e/s will represent the double heterozygote which is wild type for both mutants.
I t can be seen that for each of the six types of experiments only two different contrived mixtures were used. As will be shown below, two different mixtures is not sufficient to provide any information on frequency dependence of fitness components.
RESULTS
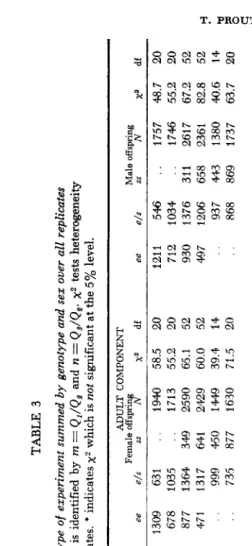
(2) Results: In Table 3 are summarized the total counts of progeny phenotypes
for each of 13 different experiments. These represent sums over the replicate cul- tures of each experiment and the sexes are recorded separately. The particular parental mixture is represented by two numbers m and n, where m is the ratio
TABLE 2
Summary of actual experiments performed using eyeless (e e ) and shaven (ss) fourth chromosome recessives
Sixty flies of each sex were used in each replicate (bottle). The numbers of each genotype in the variable sex are shown.
F E C W I T Y
Female Mixture
ee els ss
(30 20 10)
(10 20 30)
(30 20 10)
(10 20 30)
(30 20 10) (10 20 30) -____-
MALE CONSTANT
PARENT
Lens_
___
ee
ee
-
21
21
-
e/s
e l s
-
27
27
15 ss
21
-
-
ss
-
V I R I L I T Y
Male Mixture
ee els ss
(30 20 10) 18
(10 20 30) 21 (30 20 10) 27
(10 20 30) 27
(30 20 10) 18
(10 20 30) 21
____--_
VIABILITY
-_____
FEWLE PARENT NALE PARENT REPS
FEMALE CONSTANT
PARELVT
ee
ee
els
e l s
-
-
-
-
ss
-
s s
ESTIMATING FITNESS COMPONENTS 133
of ee to e / s and n is the ratio ss to e / s in the parental mixture, or in terms of Figure 1
No information is lost by this transformation since the proportions of the three phenotypes in the mixture ( Q I , Q2, Qg) can be readily calculated from m and n.
The chi-squares are calculated to detect heterogeneity among the replicate cultures. I n most all cases these chi-squares are highly significant.
This heterogeneity probably arises from two sources. The actual fitnesses probably vary somewhat from culture to culture. Also, there is a component of random drift not accounted for, since each culture was started with only 60 par- ents in the mixture.
( 3 ) Analysis: This fact of heterogeneity unfortunately would place in question
the results of a maximum likelihood analysis performed on the totals presented in Table 3 .
There is no difficulty in obtaining maximum likelihood estimates, but because of the heterogeneity, the error variances of the estimates would be too small com- pared to the true sampling variance, and therefore, any test of goodness of fit too demanding.
Consequently less efficient and perhaps less rigorous statistical procedures were adopted.
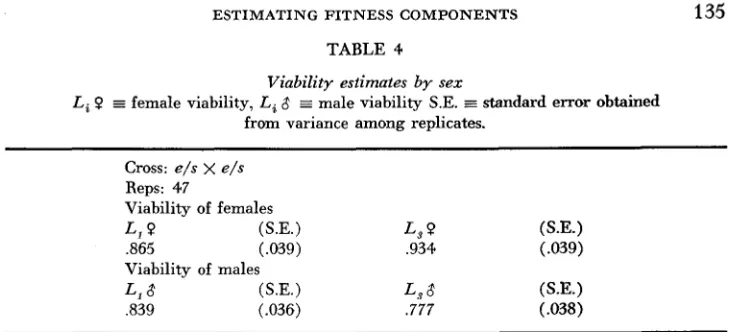
First, the estimates of the larval component of fitness (viability) are straight- forward. Assuming a normal segregation ratio in the cross e / s x e / s , then each replicate culture will yield an estimate of viability by the relations
m = Q 1 / Q 2 n =
QdQZ
LI = ( 2 Q I ) / Q 2 and L , = ( 2 Q3>/Qz
This was done for the sexes separately, and in Table
4
are shown the un- weighted means together with their empirical standard errors. These values sug- gest rather mild viability effects with a pattern of overdominance and a sugges- tion of sex dependence for shaven only.The estimation of the adult components is not so straightforward. As will be shown presently, the progeny of a single parental mixture do not contain suffi- cient information to estimate two adult fitnesses; at least two different mixtures are required. Therefore, unlike viability, it is not possible to obtain many indi- vidual estimates, one from each replicate, since each replicate necessarily results from just one mixture. In what follows is described a transformation of the data from each replicate such that the empirical variance of this transformation among replicates can ultimately be used to estimate the error variance of the component estimate. I n Table 5 , equations are set forth which are derived according to the model shown in Figure 1. The process is broken into two steps: First are shown on the right side of the table, the genotypic proportions at the time of fertilization
TABLE 3 Progenies from each type of ezperiment summed by genotype and sex over all replicates The particular mixture is identified by m = Q,/Q, and n = Qs/Q,. x2 tests heterogeneity among replicates. * indicates
x*
which is not significant at the 5% level. CONSTANT MIX PARENT m n REPS ee ee$ $ 1.5 .5 21 1309 .5 1.5 21 678 Fecundity e/s$ 8 1.5 .5 27 877 .5 1.5 27 47 1 ss$ 8 1.5 .5 15 .. .5 1.5 21 .. ADULT COMPONENT Female offspnng e/s ss N x2 df ee - 631 . . 1940 58.5 20 121 1 1035 . . 1713 55.2 20 712 1364 349 2590 65.1 52 930 1317 641 2429 60.0 52 497 999 460 1449 39.4 14 .. 735 U77 1630 71.5 20 ..4s 546 1034 1376 1206 93
7 868 Male offspring ss N xa df . . 1757 48.7 20 . . 1746 55.2 20 311 2617 67.2 52 658 2361 82.8 52 9 4+3 1380 40.6 14 cd 869 1737 63.7 20
0
I!
2
ESTIMATING FITNESS COMPONENTS
TABLE 4
Viability estimates b y sex
female viability, Li 8 E male viability S.E.
from variance among replicates.
Li 0 standard error obtained
135
Cross: e/s x e/s
Reps: 47
Viability of females
L , 0 (S.E.) L, (S.E.)
,865 (.039) ,934 (.039) Viability of males
L l 3 (S.E.) L.7 8 (S.E.)
,839 (.036) .777 (.038)
constant parent are analogously derived by simply substituting a homozygous parent in the upper-right part of Figure 1.
As in Figure 1, Table 5 shows the fecundity case with parameters
F ,
and F,. For virility, the equations have exactly the same form with the substitution of parameters V , and V , and for male proportions, P,.It will be assumed that the viability estimates from the F, experiments shown in Table
4
are accurate and have the same values in the mixture experiments, i.e., frequency dependence of viability and errors in the estimates are assumed to be small. Thus in reference to the equations of Table 5, the following are known: the frequencies of the offspring genotypes,Q1,
Q2,Q,;
the ratios i n the parentalmixture, m, n ; and the viabilities. L, and L,.
There remains to be estimated the unknown adult parameters F , and F,. I n the two cases where the constant parents are homozygotes (ee and ss)
,
only two classes of offspring are produced, a homozygous class and a heterozygous class. Thus each of these homozygous constant parents can provide but one equa- tion which is not sufficient for estimating two parameters. Two or more different parental mixtures ( m , n ) are necessary to provide sufficient independent equa- tions for each constant parent. It is less obvious, but this is also true for the case with heterozygous constant parent. Although this cross produces three classes of offspring, the heterozygous classQ2
is fixed at1/2
at fertilization. Thus the tWQ fertilization homozygous classes must sum to1/2.
This means that the cross with heterozygous constant parent will provide, at most, one equation for estimation of F , and F , for each m, n mixture.I t is convenient to express this single equation from each of the threo types of constant parent by making the transformation shown in Table 6. It can be seen that the theoretical expectation of the transformation, r , has the same form for the three constant parents. Reference to Figure 1 will reveal that r is simply the gene ratio in the parental mixture after the parents have been selected according to F , and F,. Thus this gene ratio represents the maximum information the pmg- eny will yield concerning fitnesses F , and F , operating on a given parental mixture n, n, and so I will be the basic statistic of the analysis.
T. PROUT
TABLE 5
Theoretical genotype frequencies
Qi, on the left, written as functions of genotype frequencies among fertilized eggs, Qi and viability Li. O n the right the
d,
are written as functions of the mixture m, n andfecundity F,. These relationships are shown for each type of constant parent.
GENOTYPE FREQUENCIES GENOTYPE FREQUENCIES AMONG FERTILIZED E m ANONG PROGENY -
136
CONSTAXT PAREET :-e%
mF + 1 1 2 - 1
‘1 = mF1 + 1 + n F 3
-
nF., + 1 1 2 Q2 = mF1 + 1 + n F 3Q = O
3
CONSTANT PARENT: e / s
mF1 + 112 Q1 = ’ I 2 mF 1 + 1 + nFg
n F + 1 1 2 mF1 + 1 + nP3
6
= 1 1 23
CONSTANT P A R E N T 3
-
mFl + 1 1 2‘2 = mFl + 1 + nF
3
n F + 1 1 2 - 3
‘3 = mFl + 1 + n F 3
ESTIMATING FITNESS COMPONENTS
TABLE 6
137
The calculation of r, the fertilization gene ratio
r is shown for progeny from each type of constant parent and the relation of r to theory.
CONSTANT PARENT: ee
TRANSFORMATION OF DATA
CONSTANT PARENT: e f s
TRANSFORMATION OF DATA
CONSTANT PARENT: ss
TRANSFOR’IATION OF DATA
THEORY
m Fl + 1f2
n Fg + 1 f 2
THEORY
m F, + 112
n F3 + 1 f 2
THEORY
m F1 + 112
n Fg + 1f2
1
-
Qi(c)
? B e
(+)
Q1
(-
+
1)
+
43
(-L,
+
1)
=1
Qo z ______
If for the heterozygous class = 1/2, then this equation can be arranged to give
1 1
Ll
The expression on the left involving known viabilities and observed progeny will be denoted by z. Thus if z is regarded as a random variable, E ( z ) = 1.
Thus the analysis which follows will be based on transforming the genotypic proportions in each culture to I, and in the cases of a n e/s constant parent, to both r and z.
Since both I and z depend on knowledge of viability effects and since these are
different in the two sexes, the values for the two cexes were calculated separately
138 T. PROUT
r i p + r i d zi9
+
zid2
zi =
=
--
2
where
i
= the ith cultureThis procedure avoids having to deal with the mild correlation between sexes within cultures for I and also for z. Also since this correlation was not strong, using the between-sex means should help, to some extent, to produce normality of the distributions of ri and zi, an assumption of the statistical tests to be per- formed.
are the means of ri and z i over replicates. t-tests were performed for statistical agree- ment of with unity, and i n all cases agreement was found. Two assumptions are here being tested: (1) that the viabilities used are correct, and (2) that
ob
=i/z,
which in turn reflects a normal segregation ratio in the gametes of theconstant e / s parent.
Before the gene ratios,
F,
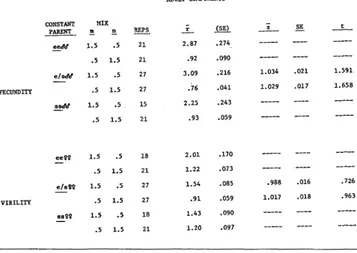
are employed for calculating the adult fitnesses, it is first possible to inquire if these adult fitnesses, whatever their values, are in-Table 7 shows the results of this calculation for each experiment;
7
andTABLE 7
Results of analysis of progenies after transforming each replicate to the gene ratio, r
The mean gene ratio,'y and its empirical standard error (S.E.) are shown.; is defined in the text. Its empirical standard error (S.E.) is shown as well as Student-t testing against unity.
ADULT COMPONENTS
CONSTANT
PAREEFT e& -
e / d b
-
FECUNDITY
S s d d
-
V I R I L I T Y
ssso
-
nix
E 9
1.5 .5 .5 1.5 1.5 .5 .5 1.5 1.5 - 5 .5 1.5
1.5 .5 .5 1.5 1.5 .5 .5 1.5 1.5 .5 .5 1.5
-
r
IcEps
-
21 2.87
21 .92
27 3.09
27 .76
15 2.25
21 .93
18 2.01
21 1.22
27 1.54
27 .91
18 1.43
E S T I M A T I N G F I T N E S S C O M P O N E N T S 139
TABLE 8
Analysis of variance of gene ratio, r, among diflerent constant parents for each of the two
mixtures, m, n in the fecundity expen'ments
Mixture: m = 1.5, n = .5
Analysis of variance
ss df MS
Within constant parents 76.60 60 1.277
Total 83.59 62
Among constant parents 6.99 2 3.495
F,,,, = 2.74 Not significant
Mixture. m = . 5 , n = 1.5
ss df MS
Within constant parents 6.00 66 .091
Among constant parents .43 2 .215
Total 6.43 68
F,,,, = 2.36 Not significant
fluenced by the constant parent to which the mixture is mated. If the adult fit- nesses operating on a given mixture are the same over the three constant parents, then that given mixture should give the same gene ratio over the three constant parents.
In Tables 8 and 9 are set forth four analyses of variance for constant-parent effect on gene ratios, one test for each parental mixture and type of experiment (fecundity or virility). Although there may be some question concerning lack of normality underlying these tests, the pattern and size differences of F-ratios strongly suggest that the adult female fitnesses are independent of the genotype of the constant male to which they are mated, while the adult male fitnesses are affected by the constant female to which they are mated. This latter case, there- fore, indicates mating interactions.
Thus the gene ratios; for fecundity experiments were combined over the three constant parents to give:
-
Mix r Standard error
m n
1.5 . 3 2.83 .146
.5 1.5 .86 .037
+
The virility gene ratios remain those shown in Table 7.
It is now necessary to calculate the adult fitnesses using the gene ratios
F
from The equation forr
shown in Table 6 can be expanded into two equations for mixture pairs.two different mixtures as follows:
- mlFl f
$4
rl =
140 T. PROUT
TABLE 9
Analysis of uariance of gene ratio, r, among diflerent constant parents for each of the
two mixtures m, n in the virility experiments
Mixture: m = 1.5, n = .5
ss df MS
Within constant parents 17.30 60 288
Among constant parents 3.61 2 1.805
Total 20.91 62
F,,,, = 6.27
ss df MS
Significant at 1 % level
Mixture: m = .5, n = 1.5
Within constant parents 8.65 66 .I31
Among constant parents 1.54 2 .770
Total 10.19 68
F2,,, = 5.88 Significant at 1 % level
-
where;,, m l , n, represent one mixture, and r, ms n, represent the other. case) giving
These can then be solved together for F , and F , ( V , and V , for the virility
-
-m, - m8
+
mzrl - m,re2 (r,nzml - rlnlm,)
F ,
___
-For the present case where m, = 1.5, nl= .5, m , =
.5,
and n, = 1.5, equations (3) become;,
- 3r,+
2F , = ----
9r, - r,
The adult component estimates together with their asymptotic standard er- rors* are shown in Table 10.
Taken at face value, these component estimates suggest for the female com- ponent a recessive deleterious effect of shaven which is independent of male part- ner. The male component on the other hand shows an overall powerful over- dominant pattern, with shaven less fit than eyeless and details depending on fe- male partners. And these adult patterns, it will be recalled, are coupled with a larval component showing a mild overdominance with a suggestion of sex de- pendence.
ESTIMATING FITNESS COMPONENTS 141
TABLE 10
Estimates of adult fitness components using gene ratio, ., and equations ( 4 )
The two estimates f o r fecundity, Fi, result after pooling over constant parents. Estimates of virility, V i i , are shown for each constant parent. The S.E. asymptotic standard errors
1. Fecundity
F , (S.E.) F 3 (S.E.)
1.037 (.122) .458 (.068)
2. Virility
Constant parent VI (S.E.) v 3 (S.E.)
ee ,363 (.074) ,039 (.033)
e / s ,243 (.04.2) ,122 (.037)
ss .135 (.036) -.018 (.030)
DISCUSSION
1. Shortcomings of the method: It should be made clear that the biology of the
eyeless and shaven genotypes are of no particular interest in themselves. They were chosen, by criteria given earlier, for the purpose of illustrating the appli- cation of the experimental system, and it is this system to which attention should be directed.
First. there are some statistical problems. The estimates obtained using just two contrived mixtures and the estimators, equations ( 3 ) are inherently unre- liable. This is because sampling errors could result in the denominators of ( 3 )
taking on the value zero. The likelihood of this increases with increasing similar- ity of m, to m, and n, to n2. This problem could be overcome by increasing the number of different mixtures together with the use of some sort of fitting pro- cedure for estimation. Increasing the number of mixtures would also allow the investigation of different subsets of mixtures, for frequency-dependence effects.
The problem of heterogeneity among replicates is, in part, an experimental one and might be alleviated by improved techniques. There is also a statistical aspect arising from the fact that two sampling errors are committed, one at the start of a n experiment due to finite numbers of parents in the mixture, and one at the end of the experiment due to the finite sample of offspring. The x2 heterogeneity test allows only for the latter. A statistical study of this problem might show that all of the heterogeneity could be accounted for and thus open the way to a maxi- mum likelihood analysis.
142 T. PROUT
2. Interpretation of estimated parameters: The virility and fecundity param- eters which have been estimated are operationally defined simply as those weights which must be attached to the parental mixture in order to account for the offspring produced. Now will be described a slightly more fundamental model from which the virility and fecundity parameters can be derived, thus providing an interpretation of these components.
If a female of genotype i encounters a male of genotype j , the number of off- spring, Rij ultimately resulting from the encounter can be constructed as follows: ( 5 ) where aij
R . . = a . . A..
c 3 1.3 2 3
probability that a mating takes place place.
k..
t 3 = - conditional expectation of fertilized eggs, given that the mating takesIt is convenient to consider the mating process measured by the aii parameter and the egg-laying process measured by kii in two separate steps.
When a population of Pi males encounters a population of virgin females, Qi, the mating process which is initiated can be be roughly regarded as involving repeated collisions resulting in the virgin female population being gradually converted into a population of mated females. It will be assumed that each fe- male mates just once to one of the j male genotypes, but that the males mate repeatedly. Let
where t
Qit
+
XZ
qijt = 1 (6)i j
= an arbitrary time after the start, Qit
qiit
the fraction of all females still virgin at time t,
the fraction of all females of type i which have mated to males of type j at time t.
If it is assumed that the general level of sexual activity of the mixture is low, but once a mating occurs it lasts only a short time, then the following equations should approximate the process:
-- dQit
-
-C Qit Z Pi aijdt i
and
where C
These equations integrate to
fraction of Qit which encounter (are courted by) males in unit time.
- Q i t = Qie-CWit and
(7)
(9)
where
-
Wi =
X
Pj aijj
ESTIMATING FITNESS COMPONENTS 1 43
attained when t
+
00 giving,Q i ( t - + a )
= O
The egg-laying step is now considered by multiplying each mated female ( 1 1) by the expectation of offspring, kij,
(12)
QiPjai jki j
Wi
Expression (12) then represents the mean number of eggs laid by a female of type i mated to a male of type
i
from a population initiated with a mixture ofvirgin females Qi and a mixture of males Pi, assuming all females mate just once. The virility and fecundity component experiments, then, simply represent special cases of (12) where the populations are initiated with one parent invari- ant, i.e., where Qi = 1 or where Pj = 1.
__
For the virility case with constant female
i,
(12) becomes P.a. . k . .3 1 1 '13
(13)
Qikij (14)
-
Wi
and for the fecundity case with constant male
i,
(12) becomesIn this latter case the factor a i j / W i , providing for differential mating, equals unity and so drops out, because only one type of male is available and all females mate with it.
It is now possible to relate expressions (13) and (14) to the model used for
estimation shown in Figure 1. Since only relative rather than absolute contribu- tions to the off spring are under consideration, (1 3) and ( 14) must be normalized as follows:
P.a.
.k..
(15)
,Pjaiik;j/Wi - - 3 23 23 - - PjAijK(z)jPjaij k i j / W i ?Pjaij kij ZPj Aij K (() j
3 3 j
for virility; and
f o r fecundity.
Aii, K ( i ) j , and K i ( j ) are parameter values normalized to the heterozygote thus:
144 T. PROUT
parameters Vij and Fii may be equated with the theoretical parameters as follows:
V i i = A . . K 2 1 (i)? . (17)
F . . - K . 2 3 - %(!) . (18)
In
the particular experiments here reported, the fact that the fecundity com- ponents showed no interactions with the different male constant parents allows further simplification. This lack of interaction means that,Klc1) = K l ( , ) = K1(,?) = F,, say and
K p ( l ) = K a ( * ) = K.9(.9) = F,, say.
I t can readily be shown that this empirical result means that in the virility case the egg-laying factor of (1 7) cannot interact with the different female constant parents, i.e., it must be that,
K ( l ) l = K,P)l = K ( 3 ) 2 = M I , say and
K ( 1 ) 3 = Krzjs= K ( 3 ) 3 = M,, say.
I n other words there may be differential effects among the male types. inhibiting or enhancing the egg laying of the females to which they are mated, but this is independent of female type. The effect is a property only of male genotype and is measured by M I and M,.
Thus the operational components can be interpreted more simply:
(19)
Fi = F i (20)
V . . = A . . M .
2.3 2 3 3
It can be seen that the mating factor Aii and the male’s effect on egg laying M i are confounded. However, what can be concluded is that the interactions
observed in Vij must have arisen solely from the mating factor Aij.
An even simpler interpretation is possible if it be assumed that the males have no differential effect on egg laying. Then (19) becomes
V . . = A . .
2 1 2 3
It should be emphasized that this final simplification still allows for a variety of biological effects on fitness. Aij may include differential death of males; also it can encompass any sort of male sterility such that, in the collision process described by equations (7) and (8)
,
a virgin female upon encountering such a sterile male is returned to the pool of virgin females to try again. Both this kind of male sterility as well as death would not give rise to interactions in Aij; rather interactions would be expected to appear due to differential acceptability by females during courtship.The factor M i would be brought into play by male sterility effects which in some way preempted the female from further encounters, thus lowering her contribution to the population of fertilized eggs. There is also the formal possi- bility that M i could arise from differential enhancement of female egg laying, but this seems less likely.
3. General considerations: The general question raised by the foregoing con-
ESTIMATING FITNESS COMPONENTS 145 Recognizing that fitness component studies can be done for a variety of pur- poses, the following will be directed toward studies done for the purpose of pre- dicting genotype frequencies in some population.
First, it seems quite clear that the study must include in one way or another the entire life cycle. I t is well known that a given genotype can affect many components of fitness, and with our present state of knowledge, there is no way to predict one component of fitness from knowledge of another. Thus an incom- plete study could lead to erroneous population predictions. The data at hand illustrate this point well. If viability alone were studied, one might conclude a mild overdominance of fitness and predict a weakly held equilibrium around
50%. If one studied fecundity alone, the conclusion would be a rapid elimination of shaven by eyeless. However, neither of these conclusions is correct, for when virility is taken into account, the system can be roughly described as one showing a powerful sex-dependent overdominance slightly favoring eyeless.
That it is necessary to include the entire life cycle in a fitness component study has been emphasized by many writers. Indeed, a great many aspects of Drosophila life history have been studied with respect to one genotype or another. However, there are not many studies like those of WALLACE (1948), LEVINE
(1952), SPIES (1955), and Moos (1955), which attempt to cover all aspects for just one set of genotypes.
A second, more important and interesting aspect of the problem arises from the fact that although the entire life cycle must be included, it is not necessary to study every detail of it; rather. it is sufficient for population prediction, to lump segments of the life cycle into a small number of net fitness components. Therefore, the problem becomes one of identifying the minimum number of such net components, i.e., it is a problem of defining LEVINS’ (1966) “sufficient parameters. ”
Both the number and kind of such net components which must be measured depend on the particular pattern of selection actually operating. Since we approach this problem from the point of view of a n investigator who is about to study an unknown system, it is safest to assume a model of selection of maximal complexity, limited only by biological plausibility, for the population regime under study.
In Table 11 the various components discussed in the previous section are em- ployed to define a series of models arranged from 1 to 5 in decreasing order of complexity. All possibilities are not shown, but rather a choice was made based on b;ologically reasonable expectations. The parameters are normalized as de-
fined earlier with the exception of K,, in Model l , which, in this caze, is normal- ized to the A a x Aa pair. It is demonstrated in the accompanying article that for any given model in Table 11 it is necessary to evaluate each component explicitly for the purpose of population prediction, assuming the type of mating bchavior discussed in the previous section.
146 T. PROUT
TABLE 1 1
Different models of fitness component participation arranged in decreasing order of complexity
Model Definition
1. 2. 3. 4.
5 .
Interactions in mating and egg laying No interactions in egg laying
No differential effect of males on egg laying No interactions 3 simple sex dependence
Classical model, A i = F i when i = i
A . . K . .
2.3 2,
A i i F , M i
A i i F ,
A i F ,
F i F ,
Symbols defined in text.
have as do eyeless-shaven, by not showing interactions in K t 3 , so that one could be safe in assuming he is dealing with Model 2. It is even possible that normally males do not differentially affect female egg laying ( M , = l ) , and that Model 3 will do; however, whether or not this is a safe generalization must await further studies of male sterility from this point of view.
It seems safe to assert that one is not justified in assuming that an unknown system is any simpler than Model 3 where interactions are still operating in the
A , , factor. I have emphasized elsewhere (PROUT 1965, 1969) that one must
assume that an unknown system may have fitness effects on the adult phase and that if it does, the effects will almost certainly be sex dependent (Model 4 ) .
Furthermore, many studies of Drosophila visible mutants suggest that mating behavior effects are common. and because of the behavioral complexities of such phenomena, interactions are to be expected (Model 3 ) . I t is interesting to note that analytic population genetic theory becomes available only at the unreason- ably simple levels of Model 4 and Model 5. Model 4 is the case of simple sex dependence which has been investigated analytically by BODMER (1965), and Model 5 is the classical textbook case.
Finally, there is the complexity introduced by frequency dependence of fitness parameters. There is mounting evidence that this may be of general importance
(KOJIMA and TOBARI 1969; EHRMAN 1966; ALLARD and ADAMS 1969).
Thus it is strongly suggested that the objective of a fitness component study be that of measuring the frequency-dependent behavior of the parameters of Model 2:
F , M , A , ,
=,,
= , IThe viability effects L , are listed to be determined separately because the biology of Drosophila allows their easy determination, and also they must be known explicitly if one is attempting to predict the population behavior where genotype frequencies are measured on adult stages each generation after viability selection has operated. Otherwise L,, may be confounded with F,, and L,, with A , , for the prediction of egg-to-egg genotype frequencies, a situation which can sometimes be arranged.
ESTIMATING FITNESS COMPONENTS 147 are confounded in V,i. The former defect is not a defect of the method in general but simply of the way it was applied in this case. The latter is a defect of the experimental system.
How-ever, it is possible that this difficulty could be overcome and
M i
could be measured explicitly if the present experimental system were supplemented by a study of mixtures of premated females similar to the technique of KOJIMA andTOBARI (1969). In this way the effect of the father on the apparent fecundity could be evaluated. Also egg laying can be studied directly on individual females mated to different male genotypes. This has been done by BUNDGAARD (1970) for certain other fourth chromosome mutants and by WOOL (1970) for certain Tribolium mutants. BUNDGAARD found no male effect on fecundity, while WOOL discovered there was such an effect.
The evaluation of the frequency dependence of the five components listed above would allow population prediction in populations with discrete generations. It is even conceivable that knowledge of these five components would go a long way toward understanding continuous, age-structured populations as well. But even if this list of components must be modified or extended, the writer wishes to emphasize the general approach of describing and measuring the entire life cycle by means of a small number of net fitness components which allow for the detection of a variety of selection patterns.
With the notable exceptions of KNIGHT and ROBERTSON (1957), WRIGHT and
KERR
i
1954), and SPOFFORD (1956), almost all of the studies of the adult com- ponents of fitness involve the collection of excess detail on one aspect of the life cycle while leaving out some other aspect. Thus for females it is not necessary to know absolute fecundity and age-specific survival; rather it is sufficient for populations with discrete generations to know relative fecundity confounded with net survival. as probably measured adequately by the fecundity parameter F , of this study.On the male side, it is sufficient to know Aii and M i ; the former includes survival. mating behavior. and certain types of sterility as discussed, while the latter includes other types of sterility. It is not necessary to understand the com- plex biology of all of these factors and it is not sufficient to understand just one of them. For instance, SPIESS and LANGER (1964), PRAKASH (1967), and others have measured mating speed, and KAUL and PARSONS (1965) measured duration of copulation. Even if it were known how differences in these measures influence A , , , it could be that for a given case the sterility or survival components of A,i are much more important.
148 T. PROUT
study of mating behavior; the next most important component is fecundity, sug- gesting a study of true fecundity; while the viability effects, by comparison, are trivial.
That this component analysis of eyeless and shaven does reasonably well satisfy the objective of population prediction is the subject of the accompanying article
(PROUT 1971).
The writer wishes to acknowledge the invaluable assistance of Dr. JEROME WILSON, Mrs. JOAN MCFARLANE, and Miss MARCIA K. WYNN. Also special thanks are due to Mrs. VIBEKA SIMONSEN for her numerical study of maximum likelihood estimators. This work was supported by a grant from the National Institutes of Health GM-06174.
SUMMARY
An experimental system is described for estimating the components of fitness operating in the adult phase of the life cycle of Drosophila. The system entails the mating together of contrived genotypic mixtures. The progeny from such crosses allow for the estimation of adult components of fitness for the sexes sepa- rately.- The application of the experimental system is illustrated by experi- ments on Drosophila melanogaster fourth chromosome recessive mutants, eyeless
( e ) and shaven ( s ) . These were studied in repulsion linkage phase in order to
have a genetic system mimicking two alleles with a distinguishable heterozygote
( e / s wild type) .- The female adult components indicate that ee and e / s have
the same fitness which is considerably greater than ss. The male adult compon- ents show a strong superiority of e/s over both homozygotes, and the depressed fitness values of the latter two vary with the female genotype to which the males are mated, indicating mating interactions. The larval component (viability) is also estimated for each genotype, and it is found that these viability effects are small compared to the adult components.- The adult fitness components esti- mated are interpreted in terms of a collision model of mating behavior involving a conditional probability of mating, aij, upon each male-female encounter. and a conditional expectation of eggs, kij, given that a mating occurs.- Stress is placed on the general approach of defining a small number of net fitness components which encompass the entire life cycle and which are accessible for experimental evaluation.
LITERATURE CITED
ALIARD, R. W. and J. ADAMS, 1969 Population studies in predominantly self-pollinating species. XIII: Intergenotypic competition and population structure in barley and wheat. Am. Naturalist 103: 621-64.5.
Differential fertility in population genetics models. Genetics 51 : 41 1-424.
A simultaneous study of several modes of selection in Drosophila melano-
Mating success and genotype frequencies in Drosophila. Animal Behavior
The genotypic control of mating speed and durations of
BODMER, W. F., 1965
BUNDGAARD, J., 1970
EHRMAN, L., 1966
KAUL, D. and P. A. PARSONS, 1965
gaster. Genetics Institute GIIC No. 21 University of Aarhus, Aarhus, Denmark.
14: 332-339.
ESTIMATING FITNESS COMPONENTS 149 KNIGHT, G. R. and A. ROBERTSON, 1957 Fitness as a measurable character in Drosophila.
Genetics 42 : 524-530.
The pattern of viability changes associated with genotypic frequency at the alcohol dehydrogenase locus in a population of Drosophila melanogaster. Genetics 61 : 201-209.
LEVINE, R. P., 1952 Adaptive responses of some third chromosome types of Drosophila pseudo- obscura. Evolution 6: 21&233.
LEVINS, R., 1966 The strategy of model building in population biology. Am. Scientist 54: 421 -43 1.
Moos, J. R , 1955 Comparative physiology of some chromosomal types in Drosophila pseudo-
PRAKASH, S , 1967 Association between mating speed and fertility in Drosophila robusta. Genetics 5 7 : 655-663.
PROUT, T., 1965 The estimation of fitnesses from genotypic frequencies. Evolution 19: 5 6 5 5 1 .
-, 1969 The estimation of fitnesses from population data. Genetics 63: 94.9-967. -,
1971 The relation between fitness components and population prediction in Drosophila. 11: Population prediction. Genetics 68: 151-167
SPIESS, E. B., 1955 Chromosomal adaptive polymorphisms in Drosophila persimilis. 11: Effects of population cage conditions on life-cycle components. Evolution 12 : 234-245.
SPIESS, E. B. and B. LANGER, 1964 Mating speed control by gene arrangements in Drosophila
persimilis. Proc. Natl. Acad. Sci. U.S. 51: 1015-1019.
SPOFFORD, J. B., 1956 The relation between expressivity and selection against eyeless in Dro- sophila melanogaster. Genetics 41 : 938-959.
WALLACE, B., 1948 Studies on “Sex-ratio” in Drosophila pseudoobscura. Evolution 2 : 189-21 7. WOOL, D., 1970 Deviations of zygotic frequencies from expectations in eggs of Tribolium
WRIGHT, S. and W. E. KERR, 1954 Experimental studies of the distribution of gene frequencies KOJIMA, K. and Y. N. TOBARI, 1969
obscura. Evolution 9: 141-151.
.
castaneum. Genetics 66: 115-132.