THE SELECTION LIMIT DUE TO THE CONFLICT
BETWEEN TRUNCATION AND STABILIZING SELECTION
WITH MUTATION
ZHAO-BANG ZENG AND WILLIAM G. HILL
Institute of Animal Genetics, University of Edinburgh, Edinburgh EH9 3JN, Scotland
Manuscript received February 1 1, 1986 Revised copy accepted August 2 5 , 1986
ABSTRACT
Long-term selection response could slow down from a decline in genetic var- iance or in selection differential or both. A model of conflict between truncation and stabilizing selection in infinite population size is analysed in terms of the reduction in selection differential. Under the assumption of a normal phenotypic distribution, the limit to selection is found to be a function of K , the intensity of truncation selection, w2, a measure of the intensity of stabilizing selection, and U*, the phenotypic variance of the character. T h e maintenance of genetic vari- ation at this limit is also analyzed in terms of mutation-selection balance by the use of the "House-of-cards" approximation. It is found that truncation selection can substantially reduce the equilibrium genetic variance below that when only stabilizing selection is acting, and the proportional reduction in variance is great- est when the selection is very weak. When truncation selection is strong, any further increase in the strength of selection has little further influence on the variance. It appears that this mutation-selection balance is insufficient to account for the high levels of genetic variation observed in many long-term selection experiments.
N artificial selection experiments and breeding programs, there may be a
I
conflict between artificial and natural selection. Usually, the aim of artificial selection is to improve some particular traits; thus, it is generally directed toward extreme phenotypic values. It is observed that natural selection often favors intermediate expression of metric traits unless these traits are very closely associated with fitness (e.g., LINNEY, BARNES and KEARSEY 1971). Soplateaux obtained in artificial selection experiments could result from the op- posing forces of directional and natural selection, rather than from a loss of additive genetic variance, as indicated by some long-term selection experiments
(e.g., LERNER and DEMPSTER 1951; CLAYTON and ROBERTSON 1957; LATTER
1966; ROBERTS 1966; WILSON et al. 1971; Yoo, NICHOLAS and RATHIE 1980). Two extreme models have been proposed for the action of natural selection on quantitative characters. One is stabilizing (or optimum) selection, another is overdominant selection. Stabilizing selection has received considerable atten- tion over the last several decades (e.g., FISHER 1930; WRIGHT 1935; HALDANE
1954; ROBERTSON 1956; KOJIMA 1959; LATTER 1960, 1970; LEWONTIN 1964;
LANDE 1975). In this selection model it is assumed that the fitness of an
1314 Z.-B. ZENG AND W. G. HILL
individual is solely a function of its phenotypic deviation from an intermediate optimum. In overdominant selection, on the other hand, it is assumed that heterozygotes at the loci affecting the quantitative character have higher fit- ness, and this model has been thought by many people to be important as a possible explanation for the maintenance of genetic variability in both artificial and natural populations (e.g., LERNER 1950, 1954; ROBERTSON 1956; BULMER
1973; GILL~ESPIE 1984).
JAMES (1962) considered the case where the conflict is between truncation and stabilizing selection. With the assumption that heritability is not greatly altered during the course of selection, he was able to develop an approximate expression for the selection limit that is expressed in terms of K , the intensity of truncation selection, w2, a measure of the intensity of stabilizing selection, and a', the phenotypic variance of the character. It is not clear, however, whether heritability is zero or not at this selection limit. In contrast, ROBERT- SON ( 1 956), VERGHESE (1 974) and NICHOLAS and ROBERTSON (1 980) investi- gated another model, called the homeostatic model, for which the conflict is between directional and overdominant selection. T h e impact of such a model on selection limits has been explored in depth. However, in all these studies, mutation as a source of introducing fresh variability has been ignored.
T h e dynamics and maintenance of genetic variability of quantitative char- acters under stabilizing selection and mutation have been studied intensively by many authors (LATTER 1960; KIMURA 1965; BULMER 1972, 1980; LANDE 1975; FLEMING 1979; TURELLI 1984). They examined the balance between mutation and stabilizing selection to see whether this balance could account for the high levels of heritable variation usually observed for many quantitative characters in natural populations. LANDE (1 975), in particular, has forcefully argued that high heritabilities could be maintained by this mutation-selection balance even with strong stabilizing selection. Recently, TURELLI (1 984) has critically reviewed this argument and numerically given the domains of appli- cability of the various approximations produced by him and other authors. Based on a Lerch's zeta function analysis and numerical work, TURELLI argued that the approximation he used would give a better estimate of the equilibrium genetic variance when the mutation rate per locus is of the order of or less. Elsewhere we have investigated the consequences of recurrent mutation on selection response in finite populations, but in the absence of any opposing effects of natural selection (HILL 1982).
In this paper, we examine the state of the population at the selection limit due to the conflict between truncation and stabilizing selection, i . e . , we inves- tigate the selection differential, heritability and the population mean at this limit. We assume that selection is on a single trait, gene effects are additive and the population is infinitely large. First, in the section below, we present an analysis on the phenotypic change due to selection; and then, in the follow- ing section, we investigate the genetic structure of the population at the limit.
ANALYSIS OF THE PHENOTYPIC CHANGE
Consider a metric character having phenotypic value x with the following probability density function among juveniles (before the operation of selection) in generation t
where U' is the phenotypic variance which is assumed to be independent of
the mean ut. T h e fitness of individuals under stabilizing selection with pheno- typic value x is assumed to decrease with deviation from the optimum value according to the relation
where the optimum value of x is taken to be zero, and U' is a measure of
intensity of stabilizing selection, being less intense the larger U' is in relation
to u2. Stabilizing selection may act on individuals through the whole life cycle, by differential viability or reproductivity, or both. But here, for convenience, it will be assumed that the stabilizing selection occurs before truncation selec- tion. Then it is readily shown that, among the survivors after stabilizing selec- tion, the phenotypic distribution becomes
w l ( x ) = exp(-x'/(2~')), ( 2 )
f
'(4
= f(x)wl(x)/Sf(.)~l(x)dx (3) = ( 2 ~ c a ~ ) - ' / ~ exp(-(x-
cu,)'/(2cu2)],where c = w'/(u2
+
U * ) is called the coefficient of centripetal selection by LATTER (1970). T h e distribution (3) is still normal with mean U ; = cut and variance U'' = cu2.Truncation selection induces the fitness function
(4)
where T is the truncation point in absolute value. Using (3) and (4), we can
then obtain the proportion of individuals surviving the two kinds of selection (the mean fitness of the population):
= P[u'/(u'
+
u ' ) ] ~ / ' exp(- u:/[2(u2+
U ' ) ] ) .where P = J: f ' ( x ) d x is the proportion of individuals surviving stabilizing selection that are then selected by truncation. T h e change in the population mean by truncation after stabilizing selection is
= KWU/(U'
+
W2)1/2,where K is the standardized selection differential of truncation selection, cor-
responding to
P.
Then the total selection differential is given byst = up
-
U *= ( u p
-
U:)+
(U:-
ut) (7)1316 Z.-B. ZENG AND W . G . HILL
This relation would hold for every generation if the phenotypic distribution remained normal before selection and U' were constant. When the second
assumption is violated, U' has to be replaced by U? in (7).
T h e change in the average phenotypic value in response to selection must equal the product of heritability and selection differential, i.e.,
where h f is the heritability of the character at generation t. Thus, at a selection limit, where Au = 0, either s or h' should be zero. If a limit is attained due to the attenuation of selection differential, the total predicted selection advance can be obtained by setting s = 0 in (7) and is
U m = KW(U'
+
W2)1'2/U ( 9 4o r
U m = KW(U'
+
W 2 ) l D / U-
Uo (9b)if uo # 0. This relation was first obtained by JAMES (1962) by a different derivation.
Furthermore, from
(7)
and (8), we can geth'u' h'xwu
%+I = (1
-
=)
ut+
(U'+
w')"'T h e first term on the right side of (10) shows the effect of stabilizing selection on the phenotypic change which is the same as shown in Equation 14 of LANDE
(1975). T h e second is the effect of truncation selection that could be constant in standard units, if the indicated assumptions hold; that is, the phenotypic distribution is always normal before selection, h' and U' do not change very
much during the course of experiment, and the same proportion of individuals is selected by truncation in every generation. If uo = 0, the selection response in the first generation of selection may be expressed as
u1 = h2Kuw/(u2
+
w')"', (1 1)and the ratio of the total response over the response in the first generation becomes
um/ul = (U'
+
w')/(u'h'). (12)Finally, solving (1 0) gives
{
h'u't]
[
{
h'u't)]
KU(U'+
w2)"*.
(13) ut = uo exp-
___+
1 - e x p-~
U'
+
w 2 U'+
U' UThen we can get the "half-life" of the selection process, the number of gen- erations taken to go halfway to the limit (ROBERTSON 1960). This is
to.5 = ln2(u2
+
w2)/(u'h2) generations. (14)the response curve is exponential, which is similar to that predicted by ROB-
ERTSON'S (1 960) theory for finite populations with directional selection alone.
In this analysis the response rate is h2a2/(a2
+
U'), a function of h2 and 02/a2. Therefore, the half-life is expected to be longer if selection is on a character with a lower heritability and/or less intense stabilizing selection (i.e., higher value of w2/a2) (14). Second, although the response rate is a function of her- itability, the selection limit predicted is independent of heritability. Third, in a large population, the total response is maximized by having the smallest possible P (9), but in a small population, P should be 0.5 to obtain the largest total response (ROBERTSON 1960).ANALYSIS OF T H E EQUILIBRIUM: DISTRIBUTION OF ALLELIC EFFECTS
In this section, we shift our analysis from the phenotypic level to the geno- typic level, and we focus attention on the distribution of allelic effects at the selection limit. T h e method of analysis is similar to that of TURELLI (1984), but with an extension to include truncation selection.
The model: Consider a randomly mating diploid population of infinite size and a quantitative character that is affected by n additive loci and an inde- pendent environmental effect. At each locus it is assumed that there is poten- tially an infinite number of allelic states, and the phenotypic effects of these alleles are continuously distributed.
Let x denote the phenotype of this quantitative character with
x = y + e (15)
and
where z: (z!) is the allelic effect of the maternally (paternally) inherited gene at the ith locus in an individual, and e is the environmental effect, assumed to be normally distributed with mean zero and variance U:.
T h e object is to find an approximation for the equilibrium distribution of zi
1318 Z.-B. ZENG AND W. G . HILL
is the density function of allelic effects after selection, where w(zJ is the selec- tion function on zi, and
g(zJ = (ZT")-"~ exp(-z?/(Zm?)] (19) is the density function of mutant effects, where m ? is the variance of mutant effects for the ith haploid locus. Interpreting (17) in words: at the beginning of generation t
+
1, an allele zi has probability 1-
pi of coming from allele ziin the previous generation, having survived selection and without mutation, and probability pi of coming from another allele in the previous generation, having survived selection and mutated to zi.
Generally speaking, it is difficult to analysis (17) directly without some sim- plification. Here we simplify (1
7)
in the following way. By expanding g(zi-
vi) in Taylor's series about (zi-
U : ) , where U{ =J
vJ:(vi)dv;, we have+
( Y 2 ) ( U ,-
U:)2g'2'(z1-
U : )+
. .
.Id U t-
- Pz[g(Z,-
U : )+
(%) v:g'2'(z,
-
U : )+
.
*.I,
where V: =
J
( U ,-
u t ) f t ( v Z ) d v , . r 2 r If g(zJ is expressed by (19),g(*)(zz
-
U : ) = (l/m,2)g(z,-
u:)[(z,-
u:)'/m;-
11. Then,-
An important observation of TURELLI (1 984) is that, under the reasonable assumption that pz 5
m,2
>>
v:,
(21)in which m,' denotes the variance of effects associated with mutation and V: denotes the equilibrium allelic variance after selection. Then the second term and the terms in higher order in the bracket in (20) can be ignored without serious error, and (1
7)
can be approximated byfi+l(ZZ) = ( 1
-
Pz)fi(z*)+
P*g(z*-
U:) (22) under the condition of (21). This is the "House-of-cards" approximation used by TURELLI (1984) and originally introduced by KINGMAN (1978). In this ap- proximation the effects of new mutants are assumed to be distributed around the population mean, and to be essentially independent of their premutation state. T h e density function of the equilibrium distribution of z, in this approx- imation is given byS
fm(%i)dZi = 1,and
(24)
Thus, if w(zJ and
g(zi
-
ui) are known,f&)
can be approximated by (23)providing pi C:
Allelic effect distribution at the limit to selection: In this section, (23) is used to find the equilibrium distribution of allelic effects zi under stabilizing and truncation selection. First, consider w(zi), the selection function on zi. Let ai = zi
-
ui be the excess of the allele zi over the mean of alleles at the ithlocus. T h e fitness of this allele, relative to the mean ui, is usually given by
wheref(x) is the density function of phenotype and w(x) is the selection func- tion on phenotype. By expandingf(x
-
a,) in a Taylor series about x, we then have that, to order a',W ( Z ~ ) = 1
+
Cui+
(?'z)Du?,(27)
where
D = s f ( 2 ) ( x ) w ( x ) d J s f ( x ) w ( ~ ) d x .
Iff(x) is the normal density function with mean U and variance u2, then
C = Au/u2
D = (Aa2
+
Au2)/u4,where Au and Aa2 are the changes in the mean and variance as a result of selection (BULMER 1980).
Since there are two kinds of selection, Au and Ao2 are determined by two components. We have already found that in
(7)
Au = K W U / ( ~ ~
+
w ' ) ~ ' ~-
uu2/(u2+
U'). (28)From (3), we have that
Aa2 (due to stabilizing) = a2(1
-
w 2 / ( u 2+
w 2 ) ) = -U'/(.'+
w2),and it is well known that
1320 Z.-B. ZENG AND W. G . HILL
where K is the intensity of truncation selection and 2 is the standard deviate
of truncation point 7. So the total change in the variance due to selection is
Aa' = -(a4
+
K ( K-
Z)a2w2)/(o'+
U'). (29)With ( 2 8 ) and (29), ( 2 7 ) becomes
(30) KUU(U'
+
u ~ ) ~ / ~-
uu2 U'+
K ( K-
Z)w22 2 ( 2
+
0 2 )w(z,) = 1
+
ai-
a,.62(U'
+
U')In (30) Au2 is not included in the term of a:. As shown later, this does not influence the results. When a, =
z,
-
U , is small in magnitude, w(zJ can also be approxi mated byKUU(C2
+
U2)1'2-
UO'a'(u'
+
U2)a
'
+
K ( K-
Z)W'2a2(a2
+
0 2 )(31) a,
-
w(z,) = exp
= exp(- (z,
-
B)*/(2A)Je,where E is a constant,
(r'(a2 -t U')
A =
U'
+
K ( K-
2)U'and
KUCJ(U'
+
U2)1'2-
U6'c l
'
+
K ( K-
z)U2
B = U i + '
Now inserting (31) into (23) and letting g(zi
-
U,) be defined by (19), wethen have the following approximation for the density function of the distri- bution of allelic effects at the limit to selection
It can be shown that in (32) U , = B, i.e.,
U = K U ( 2
+
U2)1'2/U (33)(see APPENDIX for proof).
T h e result (33) has two implications: First, since ui in (32) could, in theory, take any value, but U = KW(C'
+
u ' ) ' / ~ / u , this shows that the mean genotype,responsible for the character. T h e increase in the number of loci is accom- panied by a decrease in the effects of individual genes. Equation (33) for the maximum response at the limit to selection is identical to (9).
Now let ai = zi
-
ui. With U = KO(CT‘+
w2)1’2/u, (32) reduces to pi exp(-a?/(Zm?)}fm(ai) = (27rmf)’” [ l
-
[ exp{-a?/(2A)}] ’ (34)which is the same as TURELLI’S equilibrium distribution of allelic effects under stabilizing selection alone. Here A = u2(u2
+
w’)/[u‘+
K ( K-
Z)w‘]. By usingLerch’s zeta function, TURELLI (1984) has been able to show that
5
=
exp{-p?7rA/m?]Vi = E(a?)
=
2pZA (35)r2 = E(a4)/[3{E(~?)}’] m?/(6pJ)
as pi + 0 for (34), where Vi is the equilibrium genetic variance due to locus
i
(haploid) and r2 is the coefficient of kurtosis for this distribution. As he pointed out, the approximations rest on the condition thatpi
<<
m?/A<<
1, (36)which will be justified numerically in the next section. In addition, it can easily be proved that this distribution is symmetric.
Considering all relevant loci, the total genetic variance can be approximated by
n
0,’ = 4piA,
i= I ( 3 7 4
if a global linkage equilibrium is assumed. In particular, if the mutation rate is equal for all loci,
4 n p u 2 ( 2
+
w2)U,’ = 4npA =
U*
+
K ( K-
z ) w 2 .A check on the approximations: T h e results of (35) were obtained by TUR-
ELLI (1 984) from (34) as approximations, as p i + 0, i . e . , p j
<<
m?/A<<
1. This condition is internally consistent with m?>>
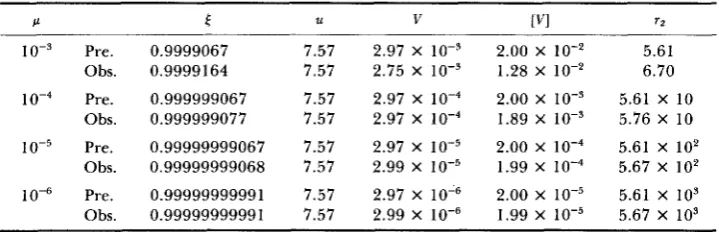
Vl (21) which leads (17) to (34) (TURELLI 1984). By a simulation of (17), TURELLI provided a numerical test of the results of (35), which clarified the conditions for the house-of-cards approximation. In this section, we provide another numerical test of (35) di- rectly from the moment calculation of (32) with truncation selection, which relies on TURELLI’S numerical calculation to support (32).1322 Z.-B. ZENG AND W. G. HILL TABLE 1
Effects of varying the mutation rate ( p ) on the analytically predicted (Pre.)
and numerically determined (Obs.) equilibrium genetic distribution for P
=
0.5,V, = 10 and tn2 = 0.05, given pi = 0.1
P € U V rvi n
IO-’ Pre. Obs. Pre. Obs. h e . Obs. Pre. Obs.
0.9999067 0.9999164 0.999999067 0.999999077 0.99999999067 0.99999999068 0.99999999991 0.9999999999 1
7.57 2.97 x 10-3 2.00 x 1 0 - 2 5.61
7.57 2.75 X lo-’ 1.28 X lo-* 6.70 7.57 2.97 X 2.00 X lo-’ 5.61 X 10 7.57 2.97 X 1.89 X lo-’ 5.76 X 10 7.57 2.97 x 10-5 2.00 x 1 0 - ~ 5.61 x 1 0 2
7.57 2.99 x 10-5 1.99 x 5.67 x 102 7.57 2.97 x 10-6 2.00 x 5.61 x 103
7.57 2.99 x 10-6 1.99 x 10-5 5.67 x 109 [ is a parameter of (32); U = K U ( ~
+
U‘)”/. is the mean genotype; V is the variance of thedistribution; [VI is the variance without truncation selection, i.e., when P = 1; and r2 = E(r,
-
u,)~/{ 3 [ E ( z , - u , ) ~ ] ~ ) is the coefficient of kurtosis of the distribution.
of these variables toward the solutions were calculated. This calculation was iterated until all increments were under the tolerance error, and the final values of the variables were regarded as the solutions. In this study, the initial guess was supplied by (33), (35) and r l = 0 with U * = 0.1, and the tolerance error was IF4.
In the computations, for convenience, all measurements except p were scaled so that U = 1. T h e results of the computations are shown in Tables 1-4, which
illustrate the effects on the analytically predicted and numerically observed equilibrium genetic distribution of varying p , m2, V, and P separately around
p = m2 = 0.05, V, = 10 and P = 0.5, with ui = 0.1 where V , = u2
+
U‘.These values of parameters are chosen to be consistent with TURELLI’S analysis, so VJu2 = 10 is equivalent to V5/u: = 20 in TURELLI (1984). T h e equilibrium genetic variance [VI without truncation selection (P = 1) is also presented in the tables for comparison. Since the distribution is symmetric, r l was found always to be zero and thus was excluded from the tables.
TABLE 2
Effects of varying the variance of the effect for the mutants (m’) for 1.1 = lo4, P = 0.5 and
V, = 10, given 1.1~ = 0.1
~
m 2 E U V [VI r2
0.001 Pre. 0.99995333 7.57 2.97 X 2.00 X IO-’ 1.12
Obs. 0.99997010 7.57 2.08 x 5.83 x 2.14
0.005 Pre. 0.99999067 7.57 2.97 x 10-4 2.00 x 10-3 5.61 Obs. 0.99999164 7.57 2.73 X 1.28 X lo-’ 6.65
Obs. 0.99999559 7.57 2.85 X 1.55 X lo-’ 1.23 X 10
Obs. 0.99999908 7.57 2.97 X 1.89 X IO-’ 5.76 X 10 0.01 Pre. 0.99999533 7.57 2.97 x 1 0 - ~ 2.00 x 10-3 1.12 x 10
0.05 Pre. 0.99999907 7.57 2.97 x 2.00 x 10-3 5.61 x I O
0.1 Pre. 0.999999533 7.57 2.97 x 10-4 2.00 x 1 0 - ~ 1.12 x 1 0 2 Obs. 0.999999536 7.57 3.01 x 1 0 - ~ 1.94 x
io4
1.15 x 1 0 2 0.5 Pre. 0.9999999067 7.57 2.97 X IO-* 2.00 x IO-’ 5.61 x IO2Obs. 0.9999999068 7.57 3.24 X 2.01 X 6.07 X lo2
TABLE 3
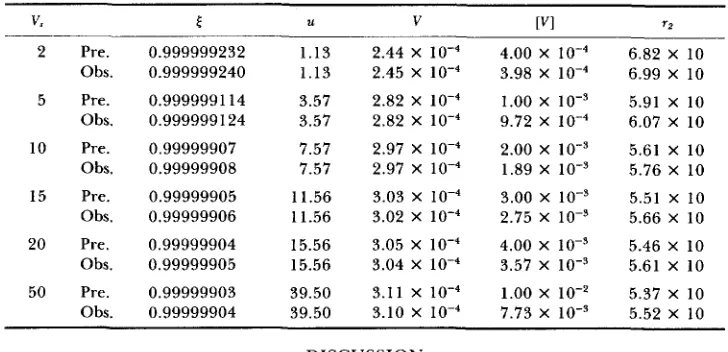
Effects of varying the intensity of stabilizing selection (V,) for 1.1 = P = 0.5 and m 2 = 0.05, given 1.18 = 0.1
2 Pre. Obs. 5 Pre.
Obs. 10 Pre.
Obs. 15 Pre.
Obs. 20 Pre.
Obs. 50 Pre.
Obs. 0.999999232 0.999999240 0.9999991 14 0,999999 124 0.99999907 0.99999908 0.99999905 0.99999906 0.99999904 0.99999905 0.99999903 0.99999904 1.13 1.13 3.57 3.57 7.57 7.57 11.56 11.56 15.56 15.56 39.50 39.50
2.44 X 4.00 X 6.82 X 10 2.45 X 3.98 X 6.99 X 10 2.82 x 10-4 1.00 x 10-3 5.91 x I O
2.97 x 10-4 2.00 x I O + 5.61 x I O
3.03 x 3.00 x 1 0 - ~ 5.51 x I O 2.82 X 9.72 X 6.07 x 10
2.97 X 1.89 X IO-’ 5.76 X 10
3.02 X 2.75 X lo-’ 5.66 X 10 3.05 X 4.00 X IO-’ 5.46 X 10 3.04 X loT4 3.57 X 5.61 X 10 3.11 x 1 0 - 4 LOO x 10-2 5.37 x 10 3.10 x IO-‘ 7.73 x IO-’ 5.52 x I O
DISCUSSION
Apart from the decline in heritability that causes plateaux in some experi- ments (e.g., BROWN and BELL 1961; ROBERTS 1966), a reduction in selection differential seems to be responsible for some other selection limits, particularly in long-term selection experiments (see FALCONER 1981, chapter 12, for re- view). It has been observed that a decline in mean was rapid after relaxation of selection, and that fertility and reproductivity were much poorer in many lines-even to the extent of causing extinction of selection lines-in many long-term selection experiments (e.g., LERNER and DEMPSTER 195 1 ; CLAYTON
and ROBERTSON 1957; LATTER 1966; ROBERTS 1966; WILSON et al. 1971; YOO,
1324 Z.-B. ZENG AND W . C . HILL TABLE 4
Effects of varying the proportion of truncation selection (P) for p = V, = 10 and m 2 = 0.05, given p i = 0.1
P t U V 72
1
.o
Pre.Obs.
0.9 Pre. Obs. 0.7 Pre.
Obs.
0.5 Pre.
Obs.
0.3 Pre. Obs. 0.1 Pre.
Obs. 0.9999937 0.9999942 0.99999825 0.99999829 0.99999887 0,99999889 0.99999907 0,99999908 0.999999 176 0.999999184 0.999999258 0.999999265 0.00 0.00 1.85 1.85 4.71 4.71 7.57 7.57 1 1
.oo
11.00 16.65 16.652.00 x
1.89 x l o + 5.57 x 10-4
5.50 x 10-4
3.59 x 1 0 - ~ 3.58 x 10-4 2.97 x 1 0 - ~ 2.97 x 10-4 2.62 x 1 0 - ~ 2.63 x 10-4 2.36 x 1 0 - 4 2.37 x 1 0 - 4
8.33 9.39 2.99 X 10 3.11 X 10
4.64 X 10 4.78 X 10 5.61 X 10 5.76 X 10 6.35 X 10 6.52 X 10 7.06 X 10 7.24 X 10
iance was not exhausted in these populations at apparent limits and also sug- gested that natural selection opposing artificial selection might cause the re- duction in the effective selection differential.
In this paper, a conflict between natural and artificial selection is examined. It is shown that stabilizing and truncation selection can take a large population to a selection limit where the population mean is determined by the intensities of truncation and stabilizing selection and the phenotypic variance of the char- acter (9). There is still genetic variance at this limit, and its amount can be approximated by (37) in terms of the mutation-selection balance. However, it is necessary to examine whether this balance can account for high levels of genetic variation observed in some long-term selection experiments.
T h e balance between mutation and stabilizing selection has been examined in detail by many authors. Both LANDE'S (1975) and TURELLI'S (1984) math- ematical calculations and parameter estimations from relevant data suggested that high levels of variation could be maintained by mutation in the face of stabilizing selection. However, when additional truncation selection is taken into account in the model, this balance may or may not be sufficient to explain the maintenance of genetic variability. Figure 1 shows the ratio of genetic variances maintained by the balance with and without truncation selection for different values of u2/a2 and P. It can be seen that truncation selection is a crucial factor in quantifying the equilibrium genetic variance. Even a small amount of selection can severely reduce the variance. But when selection is strong, any further increase in the strength of selection has little further influ- ence on the variance. Relevant data for estimating n p and w'/u' have been reviewed by TURELLI (1984), who considered that n p =: 0.01 and w2/o' =: 5-
10 might be typical estimates for many quantitative characters. With those values, the house-of-cards approximation predicts heritabilities (h2 = 4np( a2
+
1.
v)
al
;
0.g
0.ld
c
.r(
CI
0 0 0.
p:
.r(
+-'
(d
FIGURE 1.-The ratio of genetic variances maintained by the balance with and without trun- cation selection is plotted against the proportion of truncation selection (P%) for different values of w'/u'. The ratio is equal to [ l
+
K ( K - Z)w2/aZ]-'.suggest that, in the presence of opposing truncation selection, the heritability can only be maintained at a low level with np = 0.01. In this case the change in the intensity of stabilizing selection does not make a significant difference to the variance. However, if n p is 0.02, rather than 0.01, the heritability at the selection limit would be about 0.303-0.088 for
P
= 0.95-0.01 and u2/a2 = 10. Clearly, the argument relies on the estimation of the relevant parameters, particularly the total mutation rate of the loci controlling the character concerned.In view of some experimental evidence that indicates that mutations in the broad sense occurring during the experiments from whatever sources may have contributed to the long-term responses (FRANKHAM 1980) and that plateaux have not been achieved in some long term selection experiments, for example the Illinois corn experiments (DUDLEY 1977), the present analysis is not a complete description of the process. In due course it will be necessary to synthesize theories in predicting responses from mutations in the absence of natural selection (HILL 1982), the present theory incorporating stabilizing se- lection at the phenotypic level, and theories of natural selection acting at the genetic level through heterozygote superiority. Nevertheless, it is hoped that the calculation presented here will be of value in interpreting some long-term selection experiments which reached a selection limit.
We thank M. TURELLI, R. LANDE and M. LYNCH for their helpful comments on an earlier draft, and R. THOMPSON for his reading of the APPENDIX.
LITERATURE CITED
BROWN, W . P. and A. E. BELL, 1961 Genetic analysis of a "plateaued" population of Drosophila
The genetic variability of polygenic characters under optimizing selection,
melanogaster. Genetics 46: 407-425.
mutation and drift. Genet. Res. 19: 17-25.
1326
BULMER, M. G., 1973
Z.-B. ZENC AND W . G. HILL
T h e maintenance of the genetic variability of polygenic characters by heterozygous advantage. Genet. Res. 22: 9-1 2.
BULMER, M. G., 1980
CLAYTON, G. A. and A. ROBERTSON, 1957
The Mathematical Theory of Quantitative Genetics. Clarendon Press, Oxford. An experimental check on quantitative genetical theory. 11. T h e long-term effects of selection. J. Genet. 55: 152-170.
DUDLEY, J. W., 1977 76 generations of selection for oil and protein percentage in maize. pp. 459-473. In: Proceedings of the International Conference on Quantitative Genetics, Edited by E. POLLAK, 0. KEMPTHORNE and T. B. BAILEY, JR. Iowa State University Press, Ames.
FALCONER, D. S., 198 1
FISHER, R. A., 1930
FLEMING, W. H., 1979
Introduction to Quantitative Genetics, Ed. 2. Longman, London.
The Genetical Theory of Natural Selection. Clarendon Press, Oxford.
Equilibrium distributions of continuous polygenic traits. S I A M J. Appl. Math. 36: 148-168.
FRANKHAM, R., 1980 Origin of genetic variation in selection lines. pp. 56-68. In: Selection Ex- periments in Laboratory and Domestic Animals, Edited by A. ROBERTSON. Commonwealth Ag- ricultural Bureaux, Slough.
GILL, P. E., W. MURRAY and M. H. WRIGHT, 1981 Practical Optimization. Academic Press, Lon-
Pleiotropic overdominance and the maintenance of genetic variation in
T h e measurement of natural selection. Proc. 9th Int. Congr. Genet. (Part don.
GILLESPIE, J. H., 1984
HAIDANE, J. B. S., 1954
polygenic characters. Genetics 107: 321-330.
1, Cargologia) 6 (Suppl.): 480-487.
40: 255-278.
HILL, W. G., 1982 Predictions of response to artificial selection from new mutations. Genet. Res.
JAMES, J. W., 1962 KIMURA, M., 1965
Conflict between directional and centripetal selection. Heredity 17: 487-499.
A stochastic model concerning the maintenance of genetic variability in quan- titative characters. Proc. Natl. Acad. Sci. USA 54: 731-736.
KINGMAN, J. F. C., 1978 A simple model for the balance between selection and mutation. J.
Appl. Probab. 15: 1-12.
993.
KOJIMA, K., 1959 Stable equilibria for the optimum model. Proc. Natl. Acad. Sci. USA 45: 989-
LANDE, R., 1975 T h e maintenance of genetic variability by mutation in a polygenic character with linked loci. Genet. Res. 2 6 221-235.
LATTER, B. D. H., 1960 Natural selection for an intermediate optimum. Aust. J. Biol. Sci. 13:
30-35.
LATTER, B. D. H., 1966 Selection for a threshold character in Drosophila. 11. Homeostatic be- havior on relaxation of selection. Genet. Res. 8: 205-218.
LATTER, B. D. H., 1970 Selection in finite populations with multiple alleles. 11. Centripetal selection, mutation, and isoallelic variation. Genetics 66: 165-1 86.
LERNER, I. M., 1950
LERNER, 1. M., 1954
LERNER, I . M. and E. R. DEMPSTER, 1951 selection in poultry. Heredity 5: 75-94.
Population Genetics and Animal Improvement. University Press, Cambridge.
Genetic Homeostasis. Oliver & Boyd, Edinburgh.
Attenuation of genetic progress under continued
T h e interaction of selection and linkage. 11. Optimum models. Genetics LEWONTIN, R. C., 1964
50: 757-782.
LINNEY, R., B. W. BARNES and M. J. KEARSEY, 1971 Variation for metrical characters in Dro-
NICHOLAS, F. W. and A. ROBERTSON, 1980 The conflict between natural and artificial selection in finite population. Theor. Appl. Genet. 5 6 57-64.
genetic nature of the limits. Genet. Res. 8: 361-375.
homozygosis. J. Genet. 5 4 236-248.
234-249.
TURELLI, M., 1984
ROBERTS, R. C., 1966 The limits to artificial selection for body weight in the mouse. 2. The
The effect of selection against extreme deviants based on deviation or on
A theory of limits in artificial selection. Proc. Roy. Soc. Lond. (Biol.) 153:
Heritable genetic variation via mutation-selection balance: Lerch's zeta meets ROBERTSON, A., 1956
ROBERTSON, A., 1960
the abdominal bristles. Theor. Pop. Biol. 25: 138-193.
selection. Genetics 7 6 163-168.
VERCHESE, M. W., 1974 Interaction between natural selection for heterozygotes and directional
WILSON, S. P., H. D. GOODALE, W. H. KYLE and E. F. GODFREY, 1971 Long-term selection for
WRIGHT, S., 1935 Evolution in populations in approximate equilibrium. J. Genet. 3 0 257-267.
YOO, B. H., F. W. NICHOLAS and K. A. RATHIE, 1980 Long-term selection for a quantitative character in large replicate populations of Drosophila melanoguster. 4. Relaxed and reverse selection. Theor. Appl. Genet. 57: 1 13-1 17.
Communicating editor: W. J. EWENS body weight in mice. J. Hered. 62: 228-234.
APPENDIX
From (32), (24) and ( 2 5 ) , we have
where [ is a constant, such that
and
In this APPENDIX, we prove U ; = B in (Al).
First, we need to clarify the range of the constant C;. Since ( A l ) is a density function and
p; is a small value, this implies that
0 < 1
-
C; exp(- (z; - B)'/(2A)}always, which suggests that 0 < [ < 1. T h e n it follows that
[
1-
t exp(-VI]'
= exp(- n(zi 2A - B)*1.
?l=O
Therefore, (AI) can be written as
1328 Z.-B. ZENG AND W . G . HI1.L
after some calculation. Note that the last two terms in the right-hand side of (A4) define a normal density function. By using this property and putting (A4) into (A2) and (A3), we then obtain
and
and
Let
and
A
+
nm?B/u,A
+
nmg 'b, = a,
(A5) and (A6) are then equivalent to
m m
1
"=O n=o A
a , = b, = -.
Sufficiency: It is easy to show that
cr=o
a , is a monotonically decreasing series with positive terms. Then, if U , > R , it follows that a , > b, for n = 1 , 2,.
. . , except n = 0, where a,, =bo. Consequently,
c:=,)
a, >cr=p=o
b, which contradicts the condition of (A7). Similarly, ifU , < B ,
c,"=~
a, <c,"=p=o
b,, which also contradicts the condition of (A7). While, when U , = B,a , = b, holds for every n = 0, 1 , 2, . . .; therefore, (A7) is satisfied.
Necessity: (A5) and (AS) can be rewritten as
and
J
n ( u , - B)' A+
nm:B/u,A I/'
= rd,,
io
'
[
X
I
e x p \ - 2 ( A 4- n m f ) j A+
nm: "=nwhich are two power series. According to the identity theorem for power series, if the two power series
m m
2
yc. and rd,have the sanie sun1 in an interval 0 < [ < 1 (in this case) in which both of them converge, then the t w o series are entirely identical. That is to say, for every n = 0, I , 2, . . ., c, = d,,
since (AS) and (A9) do converge with 0 < f < 1 . I t then gives U , = B always. Thus, the
proof is completed.