DEVIATIONS
OF
GENOTYPIC STRUCTURESFROM HARDY-
WEINBERG PROPORTIONS UNDER RANDOM MATING AND DIFFERENTIAL SELECTION BETWEEN
THE
SEXESMARTIN ZIEHE AND HANS-ROLF GREGORIUS
Lehrstuhl fur Forstgenetik und Forstpflanzenziichtung der Universitat Gottingen, Biisgenweg 2,0-3400 'Gottingen- Weende, Federal Republic of Germany
Manuscript received August 1, 1980
Revised copy received January 5, 1981
ABSTRACT
Population genetic models, such as differential viability selection between the sexes and differential multiplicative fecundity contributions of the sexes, are considered for a single multiallelic locus. These selection models usually produce deviations of the zygotic genotype frequencies from Hardy-Weinberg proportions. The deviations are investigated (with special emphasis put on
equilibrium states) to quantify the effect of selective asymmetry in the two sexes. For many selection regimes, the present results demonstrate a strong affinity of zygotic genotype frequencies for Hardy-Weinberg proportions after two generations, at the latest. It is shown that the deviations of genotypic equilibria from the correspcnding Hardy-Weinberg proportions can be ex- pressed and estimated by means of selection components of only that sex with the lower selection intensity. This corresponds to the well-known fact that viability selection acting in only one sex yields Hardy-Weinberg equilibria.
S I N C E reliable techniques for genotype identification are available, it has be- come common to investigate mating systems active in natural populations by recording the genotypic structure at early developmental stages (as near as possible to the zygotic stage). The theoretical results on how genotypic structures in the offspring are characteristically related to mating systems of the parental populations serve as the basis for interpretation. In this context, it is common usage to present the observations as deviations from Hardy-Weinberg propor- tions.
It is well known that, in spite of random mating, Hardy-Weinberg proportions at a single autosomal locus are not necessarily realized i f , for instance, allelic frequencies in the parents differ between the sexes. This situation is typical for differential viability selection in the two sexes of dioecious populations (LI 1967; ANDRESEN 1978). Even if the two sexes show identical genotypic structures, random mating generally does not result in Hardy-Weinberg proportions for the zygotic population if the fecundities of the mating types differ (Roux 1977). Hence, it might be wrong to attribute deviations from Hardy-Weinberg pro- portions to nonrandom mating. On the other hand, BODMER (1965), CANNINGS
216 M. Z I E H E A N D H - R . GREGORIUS
viability selection at a single multiallelic locus, when restricted to one sex, re- sults in Hardy-Weinberg proportions for populations in equilibrium. For the diallelic case of this model, ZIEHE (1981) recently proved convergence for all population trajectories, which implies either fixation of a single allele or con- vergence to a nontrivial state possessing Hardy-Weinberg proportions (depend- ing on the array of fitness values). However, in all other situations, except the classical model of viability selection, nothing was known until now about such general convergence properties, and no general rules exist stating how to evaluate possible deviations of equilibria from Hardy-Weinberg proportions.
This observation raises the question of the extent to which random matirg is
capable of counterbalancing the effects of differential selection in the two sexes. I n particular, a n answer to this question would be helpful for interpreting ob- served genotype frequencies (at the zygotic stage) with respect to the popula- tion dynamic forces responsible.
THE M O D E L
For dioecious individuals reproducing in nonoverlapping generations in a randomly mating population of effectively infinite size, a single autosomal locus, with alleles A,,
.
. .
,A,, is considered. The alleles constitute ordered genotypes AiAj,i,i
= 1,. . .
,n, where the first allele is received from the female parent, the second from the male.Let the survival probabilities from the zygotic to the reproductive stage of the ordered female and male genotypes AiAj, respectively, be denoted by v f j , U$ for
i,i
= 1,. .
.
,n. Denote byfTj
and f $ the corresponding multiplicative fe- cundity contributions to zygotic offspring, i.e., letf g j
f p i be the average num- ber of zygotes producd by a mating of a female of genotype AiAj and a male of genotype AkA 1. These models of multiplicative fecundities may be applied,for example, to differential gamete production and random union of the gametes in plant populations, where the j ! j and f & are commonly referred to as fertili- ties. The genotypic fitness components, which may differ between the sexes, will be written as wyj and w;, such that w f j = u t j f $ and
WE.
= v sf$
fori,i
=1,.
.
.
,n. Assuming no influence of the ordering of the alleles within a genotype on its survival probability and its fecundity, we have wyj=
wyi and w f j = wfi. Let Pij denote the relative frequency of the ordered genotype AiA j at the zygotic stage of the population. In the absence of sex distortion, Pij is the same among both the female and male zygotes. Let pf and pf be the allele frequenciesof
A i
within the female and the male contributions, respectively, of the preced- ing generation to the zygote population ( p ? S P i j , pp’ = Z P i i ) . The allele fre-quency for Ai in the zygote population is then
1 . 3 1 . 1
DIFFERENTIAL SELECTION 21 7 useful. Average female and male fitness contributions for those alleles present i n the population can be defined as
and
-
Yw Q ( P ) : =:-
plc
k
The total female and male average fitness contributions are w ? (P) and wd (P) with
iis?(P) =
z
Pkzw;plk, 1
and
I n the following, the functional ( P ) notation will be suppressed as long as the meaning is obvious. Indicating frequencies in the next generations by primes, the recurrence equations can then be shown to take on the following form for
P k , p l # 0 (see e.g., Roux 1977 or GREGORIUS and ZIEHE, submitted for publica-
tion)
and
I n all subsequent considerations, we shall assume that the smallest positive value for the w&’s, as well as that for the w& ’s, is equal to 1. This can be realized by dividing all w&’s by the minimum of all those w,’ ’s that are not equal to 0 and by dividing all w f z ’ s by the corresponding male value. Clearly, this does not affect the generality of the conclusions, since we are concerned ex- clusively with relative frequencies.
CONSEQUENCES F O R GENOTYPE FREQUENCY TRAJECTORIES
The phenomenon of heterozygote excess, relative to Hardy-Weinberg propor- tions (HWP)
,
as a consequence of asymmetric allelic contributions of the (ran- domly mating) sexes is formulated in BUNDGAARD and CHRISTIANSEN (1972) and ANDRESEN (1978) for the case of two alleles, and with multiallelic extensions i n PURSER (1966). Using Eqs. (1) and ( 2 ) , the deviations in the multiallelic case read1
2( q L
+
Pi,) - 2 p:p: = ( p f ’ p f ’+
pp’pf’)-
- !p,p’+ p f ’ ) (Pf’+
pt”)218 M. ZIEHE A N D H-R. GREGORIUS and for homozygotes to
(3b) Thus, after at most one generation, every homozygote frequency is less than or equal to the corresponding
H W P ,
and the total sum of heterozygote frequencies exceeds or is equal to the corresponding Hardy-Weinberg sum. This kind of homozygote decrement relative to panmixia is likewise described for models of additive fecundity selection ( GREGORIUS and ZIEHE, submitted for publication; ZIEHE 198 1 ).
It
is already clear from EQs. (3a) and (3b) that differential allelic contri- butions of the sexes to the following generation in general produce a genotype frequency deviation fromHWP.
The main aim of this paper is to estimate the maximal possible deviation of zygotic genotype frequencies due to differential viability and differential multiplicative fecundity selection between the sexes. For a genotype carrying the alleles A k and At, this deviation shall be denoted by d k z :p k - # ; = - -
1 d’- 0 ’ 2( p k
p k
),
which is always nonpositive.d k l = P k Z
+
P l k - 2pkpz fork
t? and d k k P k k-
PE.
Hence d k z for k f
I
refers to the sum of the deviations of the ordered genotypesAkAl and A Z A k .
Using Eq. ( I b ) for Pk,PZ f 0 and the new notation, Eqs. (3a) and (3b) can be rewritten as
The above equations also indicate that the genotype trajectories approach HWP if the frequency-dependent allelic contributions have a similar structure in both sexes. I n the special case of symmetrical selection in the two sexes, fe- male and male allelic contributions to the next generation are identical, and a Hardy-Weinberg structure is reached after the first generation, This observation demonstrates that the deviations from H W P depend basically on the asymmetry relationships for each single allele. Thus, it could happen that HWP are realized for only a few of the alleles under consideration. On the other hand, since the allelic contributions are frequency dependent, the deviations from HWP cannot be expressed solely as a function of the asymmetry i n the selection coefficients. This requires a measure that separates the influence of frequency dependence from that of asymmetric selection between the sexes.
DIFFERENTIAL SELECTION 219 primary interest lies in finding appropriate inequalities that involve a meaning- ful measure of asymmetry in the selection coefficients. APPENDIX A includes the derivation of such a n inequality that gives an upper limit for the difference be- tween p i and p f . The inequality contains two parameters, Ak and L , ( P ) , which need to be defined. L w ( P ) is the product of two factors, of which one is the sum of all those genotype frequencies with female genotypic fitness contributions not equal to 0 and the other refers analogously to the male genotypes. Thus,
Clearly, ii no male and no female fitness contribution is 0, as will be assumed in much of the following sections, both factors add up to 1, and Lw(P) = 1 is independent of P. The other parameter, A,, is the maximum of the absolute asymmetry coefficients \ A 2
I
in which the kth allele is involved:Ak=max lA’$!,whereAk,:=w$wfi-w$wTi.
6 2 , Z
In the case of only two alleles, A, and A2 are identical and can easily be com- Now, following APPENDIX A, after one generation
puted as the maximum of {
I
Ai:I,
I
A;:I, I
A;:I
}.
and after two generations
Remember that n is the number of alleles considered. In combination with Eqs. (3a) and (3b), two useful inequalities for the maximum deviation of genotype frequencies from HWP after one and two generations, respectively, follow:
-
and
In APPENDIX B, certain properties of the A i s are discussed. Among these, the most interesting is that, provided Ak = 0 for one
k,
it follows immediately thatall A, = 0. Thus, it is very unlikely that a selection regime that implies HVVP for one allele (irrespective of the genotype frequencies) does not do so for all of
the other alleles. In particular, denoting by A the maximum of all Ak (A = max A h ) , it is shown in APPENDIX B that A = 0 if and only if selection acts symmetri- cally in both sexes. Given this fact and the observation that the Ak/is vary mono-
220 M. Z I E H E A N D H-R. GREGORIUS
tonically with the selection coefficients, A is an appropriate measure of asyni- metry. Eq. (8) shows that
B = - ( 1 - 1 4(n-l)
)'
A'/Lw'(~')32 (9)
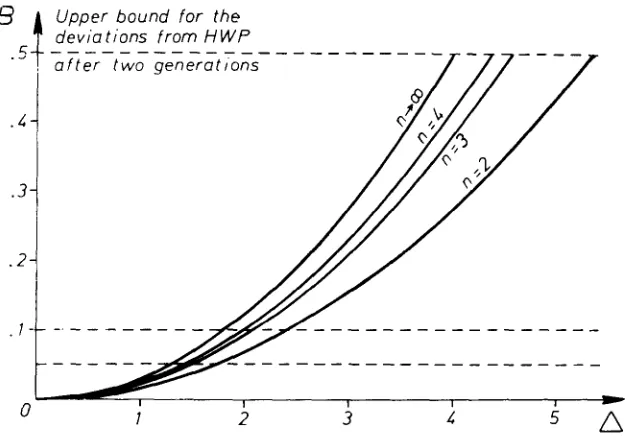
can be considered as a n upper bound f o r the deviations of the frequencies of the single genotypes from the corresponding HW frequencies after two generations at the latest. In the following section, we shall therefore demonstrate the behavior of this upper bound as a joint function of the measure of asymmetry, 4, and the number of alleles, n, and shall apply this to particular models of selection.
APPLICATIONS A N D ILLUSTRATIONS
( a ) The upper bound for the deviations of the genotype frequencies from
H W P :
Assuming that all selection coefficients are positive (the smallest in eachsex being equal to l ) , L w ( P ) = 1 and the upper deviation bound B is a parabola in A with a coefficient depending on the number of alleles n. Figure 1 illustrates this deviation bound for several values of n. Since a deviation from
HWP
cannot exceed 0.5, it is seen from (9) that the upper bound B contains relevant informa- tion only for those A smaller than 16/3. This value is reduced as the number ofalleles increases. Moreover, the practical importance of the upper bound in- creases rapidly as A decreases.
( b ) Diallelic differential viability and multiplicatiue fecundity selection between the sexes: For the diallelic case n = 2, the following property of the deviations from H W P is valid for every genotypic structure:
1 2
0 5
-
d12 = -dll = -dzz.
The computation of the asymmetry coefficient A reduces f o r n = 2 to
Assuming again that all selection coefficients are positive, the inequalities ( 7 ) and (8) that delimit the deviations from H W P after one and after two genera- tions, respectively, take on the following form:
A = A , = ~ , = m a x {lw~,w$ -wfIwlpZ), I w ~ p f , - ~ ~ ~ ~ w ~ ~ I , ~ w , 9 1 w 2 $ 2 - w ~ p ~ 2 ~ } ~
1 4
DIFFERENTIAL SELECTION 221 Upper bound for the
' k
deviations f r o m H W P. L -
.3-
. 2 -
I I I t
FIGURE 1.--Illustration of the upper bound B f o r the deviations from HWP as a function of
the number of alleles, n, and the asymmetry measure, A .
Thus, for example, if the selective advantage of the female A,A, relative to the female A, A, and of the male A,A, relative to the inale A,A,, respectively, is less than 32%, the deviation after more than one generation never exceeds 1%.
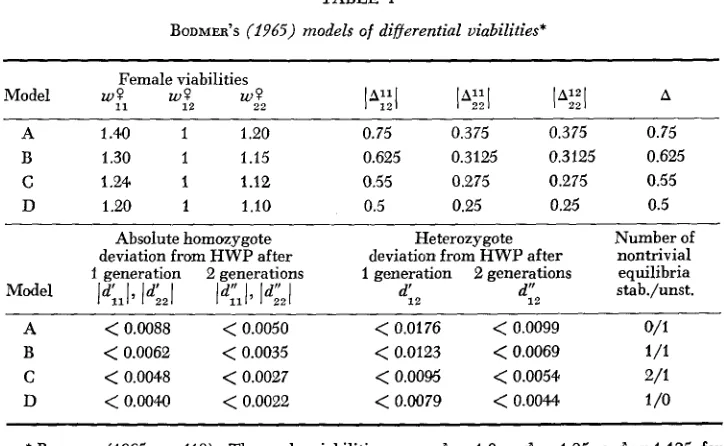
Furlher examples: Models A-D are taken from BODMER (1965). They are characterized as having the same ratio of homozygote-heterozygote viability differences in both sexes [i.e., (w,O
-
W & ) / ( W ? ~-
w&)
= (w:',-
w $ ) / ( w $ ~ -w$)
1.
The differential viabilities are listed in Table 1. All models have one non- trivial equilibrium exhibiting HWP, which is stable only in modelD.
Models B and C present further stable polymorphic equilibria with positive distances to HWP. The upper deviation bounds after one and after two generations, respec- tively, calculated from Eqs. (10) and (1 1 ),
are given in Table 1. It can be seen that the trajectories' deviations from H W P after two generations never exceed( c ) Viability and fecundity selection acting only in one sex: 'She case of via- bility and fecundity selection acting only in the male sex is considered (w,P, = l for all k , l ) . Ak and A can be computed as
1%.
A, = max iwfj
-
wfiI
and A = max Ak = max w f l-
min w $.
r , i , Z s,j k,i
Under the assumption that no male selection coefficient is 0, due to the standardi- zation of the coefficients, we have min w$
=
1, and A is interpretable as the se- lective advantage of the male genotype with the highest selection coefficient rela- tive to that with the lowest. For the &allelic case, inequalities (10) and (1 1) are again applicable. The inequalities demonstrate that if, in the male sex, the222 M. Z I E H E A N D H - R . GREGORIUS
TABLE 1
BODMER'S (1965) models of differential viabilities+
Female viabilities
Model 9 1
we
w,Pl"3
l"3
l A 3 AA 1.40 1 1.20 0.75 0.375 0.375 0.75
B 1.30 1 1.15 0.625 0.3125 0.3125 0.625
C 1.24 1 1.12 0.55 0.275 0.275 0.55
D 1.20 1 1.10 0.5 0.25 0.25 0.5
Absolute homozygote Heterozygote Number of deviation from HWP after deviation from HWP after nontrivial 1 generation 2 generations 1 generation 2 generations equilibria stab./unst.
ld;ll, IdLI Id:lL ld;,l d:, d:2
A
<
0.0088<
0.0050<
0.0176<
0.0099 0/1B
<
0.0062<
0.0035<
0.0123<
0.0069 1/1C
<
0.004.8<
0.0027<
0.0095<
0.005+ 2/1D
<
0.0044l<
0.0022<
0.0079<
0.0044 1 /o* BODMER (1965, p. 418). The male viabilities are wd = 1.0, w$ = 1.25, w d = 1.125 for
11 22
models A-D.
selective advantage relative to that genotype with the lowest selection coefficient is less than 100% for each of the other genotypes (i.e., A
<
I), the absolute homozygote deviation fromHWP
never exceeds 2% after the first generation and 1% after the second generation. For models of viability selection acting only in one sex, ZIEHE (1981) proved that every trajectory that has arrived a tHWP
has either reached an equilibrium or will leave the set of Hardy-Weinberg structures in the following generation. The described affinity of the trajectories f o rHWP,
thus, is the more remarkable.Examples: Two numerical examples
E
and F may conclude the considerations on selection acting only in one sex. They are taken from GREGORIUS and ZIEHE(submitted for publication.) The male selection coefficients and the results for
TABLE 2
TWQ examples for selection acting only in the male sex'
Absolute homozygote Heterozygote deviation from HWP after
1 generation 2 generations
deviation from HWP after 1 generation 2 generations
Model w f 1 w$ w,", A P;lL K 2 I I d 3 I d 2 $12 d L
E 1 1.158 1.105 0.158
<
0.00040<
0.00022<
0.00079<
0.00044F 1.222 1 1.167 0.222
<
0.00078<
0.00044<
0.00155<
0.00087D I F F E R E N T I A L SELECTION 223 the deviation bounds are presented in Table 2. An extreme trajectory attractivity of HWP is apparent.
( d ) Additive fecundity selectio'n: As pointed out in ZIEHE (1981), models of additive fecundity selection lead tu the same structure of the recurrence equa- tions for unordered genotype frequencies as models of viability selection acting only in one sex, provided the selection coefficients of the sex undergoing selec- tion are interpreted in a suitable manner. These coefficients must be conceived as the sums of the additive fecundity contributions of those pairs of mates with the same female and male genotype [for a more detailed explanation, see GREGORIUS and ZIEHE (submitted for publication) and ZIEHE (1981)l. Then, the statements of (c) are valid and applicable in the same way, taking, for exam- ple, fur
a
the difference between the maximum and the minimum of the stan- dardized sums of genotypically identical female and male additive fecundity contributions.( e ) Diflerential mating selection acting only in one sex: Models with random contact of individuals and differential probabilities for either fertilization or perfurmance of mating determined only by the genotypes of one sex are equiva- lent to models of viability selection acting only in one sex. The methods of (c) are feasible after standardization of the probabilities.
HARDY-WEINBERG DEVIATIONS O F EQUILIBRIA
A great effort has been made to elaborate conditions for the existence of one, two or three polymorphic equilibria for the diallelic model and to determine their stability properties (BODMER
1965;
M ~ R A T
1969), as well as to classify se- lection regimes admitting of one or more stable or unstable equilibria(KIDWELL
et a2. 1977). However, to date, little has been known about the location of these equilibria. Referring to our particular objective, our interest is focused on the position of equilibria relative to the corresponding Hardy-Weinberg proportions. A necessary condition for a genotypic structureP*
to be at equilibrium, is, of course, thatp l f
= p l for allk.
From (I), this condition is equivalent to
z"f*
q*
-
ZP*+=--
-
2w d * for all k with p ; # 0, where an asterisk
denotes the functional dependence of the fitness contributions on
P*.
W ?
*
Thus at equilibrium,
(
wdi
-
5)
may be written as(
l Z o *-
w k - ? *)
,
( E g * - z " d * ) or 2 x,
d
*
Z?*
2 X
224 M. Z I E H E A N D H - R . GREGORIUS
(E?*
-
Z ? * ) ( Z ? *-
E ? * )-"*
- -"*)(E"* - - E *( W k w U ) i
1
-____
= 2P:P: ( Z $ * ) *
for k f l and
dtk =
-PE"
(
Z,,"*--9* Z ?*
1
=-p;z ( z g * - G 8 * Z d *)
'
.
(12b)
An immediate consequence of Eqs. (12a) and (12b) is that equilibria are of Hardy-Weinberg type under selection acting only in one sex. Equations (12a) and (12b) suggest that, if selection coefficients in one sex differ only slightly, then there should be a strong equilibrium affinity to HWP. I n other words, one expects that deviations from H W P are strongly related to selection conditions of that sex showing the smaller differences in selection coefficients. The rest of this paper will be devoted to an investigation of this conjecture.
In
APPENDIX c, the following inequality for p5 # 0 is proved:where A ? =maxlw? - w ?
I
and L? ( P ) = 1-
2 P,, = Z Puv.k 1,1,1 k l 1; VV U p
w ? = O U, W E v # O
An analogous formulation f o r the male sex with the corresponding definitions of A$ and L$ ( P ) can be derived i n the same way.
Therefore, upper limits for the deviations of the genotypic equilibrium fre- quencies from H W P can be stated for pl,p: # 0:
for k # l and
[
*,pZ
Of2- - ( 1 - 1 4(n-l)
)'
Xmin c Lp,(P)2,
L $ ( P ) 2 ) i d ; k S O . (15) 16Denoting the maximum of all A i ,
k =
1,. . .
,n, by A ? and the respective male maximum by Ad, these coefficients measure the variation of the selection Coefficients separately in the sexes and can be written asA ? = max wzl
-
min~ 7 %
and A d = max w $-
minW E
,k, 3 1 , s k J 1 . 2
D I F F E R E N T I A L SELECTION 225 value relative to the female genotype with the lowest; Ad has an analogous
interpretation for the male genotypes. By applying Eqs. (14) and (15), the expression
)2 X min (A?’, Ad2}
1
Be
=8’
(
4(n-1)is an upper bound f o r the equilibrium deviations from HWP. Since Be depends only on the variation of the selection coefficients, neglecting the possibility of an affinity to HWP produced by symmetrical selection, this b u n d makes sense only if the maximum of the selection coefficients in one sex is sufficiently small. Examples: Two diallelelic models G and
H
of differential viability selection in the sexes are considered. Except for the heterozygote female viability w : ~ , they possess the same viability structures:w$
=
1, wf2= 3,w,$,
,= 0,w e
= 1,w : ~ = 1.25. However, the models demonstrate different trajectory behavior, espe- cially relative to HWP.
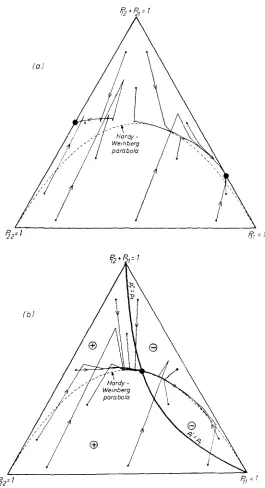
wP,
= 0 in model G, a few trajectories of which are traced in Figure 2a. The figure shows that, under certain differential viability selection models, trajectories starting near HWP may leave the immediate neigh- borhood of the Hardy-Weinberg parabola and converge to the margin of the triangle. It is notable that, for model G, there exist two stable equilibria at the margin, one of which has a heterozygote equilibrium deviation of more than10% from HWP (d:z = 0.125).
The heterozygote female viability for model
H
is chosen as w,pZ = 1.04. This implies that A ?=
0.25, so that Eq. (16) leads to an upper equilibrium deviation bound B , = 0.0044. The exact heterozygote deviation of the single polymorphic equilibrium is greater than 0.0013 and less than 0.0014. Figure 2b illustrates the trajectory development.DISCUSSION
The classical selection model of symmetrical viabilities in the sexes leads to Hardy-Weinberg proportions (HWP) after one generation. By analyzing the development of genotypic frequencies in zygotes, the problem can be reduced to the study of allelic recurrence equations. However, in natural populations, it might be more realistic to separate the functions of the sexes in connection with sexual reproduction, which could imply asymmetric allelic contributions of the sexes. But, in general, this asymmetry generates deviations from HWP of the zygotic genotypic frequencies and complicates the consideration of genotypic structure development.
226 M. Z I E H E A N D H-R. GREGORIUS
FIGURE 2.-(a) DeFinetti diagram that illustrates the trajectory development under selection indicate regions of model G. (b) Trajectory development under selection model H. @ and
DIFFERENTIAL SELECTION 227
Relative to the corresponding HWP, there already occurs a decrement in frequency for each homozygote and an excess of the sum of all heterozygote frequencies (Eqs. 4a74b). However, the amount of the deviation from
HWP
strongly depends on frequency-independent asymmetry coefficients (see AP- PENDIX B) of selection. These coefficients generally allow specification of a useful upper bound for the absolute deviations from corresponding
HWP
after one and two generations at the latest (Eq. 9). As demonstrated for some examples taken from the current literature, the deviations of the genotypic trajectories fromHWP
become very small after several generations unless the selective differences of the genotypes show extreme asymmetry between the sexes.The second part of this paper is concerned with deviations of genotypic equi- libria from HWP.
It
is known that selection acting in only one sex leads to Hardy-Weinberg equilibria, if they exist, and that the introduction of slight se- lective differences f o r the nonselected sex can immediately change the number and type of nontrivial equilibria. Nevertheless, the equilibrium deviations from HWP remain small and are controlled by that sex with the lower selection in- tensity. An upper bound for the deviation is presented (Eqs. 14,15). These find- ings do not depend on whether or not the equilibria are stable.The described
HWP
affinity of zygotic genotypic frequencies appears to be a characteristic property of many representations of the described models of dif- ferential selection between the sexes. The effect of random combination of the alleles following selection in the sexes considerably reduces the generation of zygotic Hardy-Weinberg deviations produced by differential allelic contribu- tions of the sexes. Otherwise, if the selectional forces are able to maintain relevant deviations fromHWP,
either selection must act extremely differently between the sexes (compare the example illustrated in Figure 2a),
or the allelic contribu- tions of the selected sexes are combined in a nonrandom manner as a consequence of a special mating system.These investigations were supported by a Heisenberg Fellowship to HRG and a grant from the Deutsche Forschungsgemeinschaft to MZ. The authors are indebted to E. GILLET for the drawings.
LITERATUKE CITED
ANDRESEN, E., 1978 A note on deviation from Hardy-Weinberg proportions due to differences i n gene frequencies between parental males and females. h i m . Blood Grps. Biochem. Genet 9: 55-58.
BODMER, W. F., 1965 Differential fertility in population genetics models. Genetics 51: 411- 424.
BUNDGAARD, J. and F. B. CHRISTIANSEN, 1972 Dynamics of polymorphisms: I. Selection com- ponents in a n experimental population of Drosophila melanogmter. Genetics 71 : 439-460.
CANNINGS, C., 1969 The study of multiallelic genetic systems by matrix methods. Genet. Res., Camb. 14: 167-183.
KIDWELL, J. F., M. T. CLEGG, F. M. STEWART and T. PROUT, 1977 Regions of stable equilibria
for models of differential selection in the two sexes under random mating. Genetics 8 5 :
171-183.
228 M. ZIEHE A N D H - R . GREGORIUS
M ~ R A T , P., 1969 SBlection diffBrente dans les deux sexes. Discussion g&n&ale des possibilitPs
PURSER, A. F., 1966 Increase i n heterozygote frequency with dif€erential fertility. Heredity
Roux, C. Z . , 1977 Fecundity differences between mating pairs for a single autosomal locus, sex
ZIEHE, M., 1981 Population trajectories for single locus additive fecundity selection and related
Corresponding editor: J. F. KIDWELL dCquilibre pour un locus autosomal & deux allhles. Ann. GBnBt. SB1. Anim. 1 : 49-65.
21: 322-327.
differences in viability and nonoverlapping generations. Theoret. Pop. Biol. 12 : 1-9.
selection models. J. of Math. Biology 11 : 33-43.
APPENDIX A
Since At: = 0, the second summation can exclude the case
(j,Z)
= ( i , k ) . Thus,W d
= o
W O 11U = o uv
DIFFERENTIAL SELECTION 229 After at most one generation, the homozygotic decrement occurs:
Pik
<_ p;; and therefore (for n>
I ) , ,= p; -Pik
2 p; (I-p’ ).
k
i #k
1 1
4 Finally, one obtains ip;”
-
pf”1 S - [I I-A P P E N D I X B
First it will be shown that Ak = 0 implies proportionality between female and male selection coefficients for the kth allele, i.e.,
w;j
= cWE.
for all j and a posi- tive constant c. Letwi.
= w : ~ ,wP.
= ?l wkOl,
and let analogous sums bedefined for the male sex. Without loss of generality, we assume that wP . .
>
0 ,vvd
>
0. (Otherwise, extinction of the population occurs after one generation. jThe assumption A, = 0 leads to wfiwfi.= w,$wyi for all j,&,
i
#k.
Summa- tion over l,i,i
#k
leads to w & ( W O ^ . .-
w f ) := U?$ ( W O . . ’- wf. ), and summingup this expression over j yields wkq wd . . := wP . . wf,. Hence, (w.4 ’- w $ ) and
( w ?
. . w,PJ are both positive or both equal to 0. In the case where they are positive, proportionality is already shown and c can be verified as W O . ./w”
. ..
Thus, assume (wd . . 8-
w,$
) and(w?.
I-w $
) to be equals to 0. This implies that= w $ ,= 0 if 1 #
k
andi
#k.
Consequently, summation of w ? ~ and w f i over1 and
i, i
#k
yieldswi.
’-w& and
w f ,-
w $,
respectively; therefore,. .
0.K WO - WP = WO
-
WP = w99-.
. k. k . kk.
. w,9,’
0:= W d
. .
8- w6 .= wf.-
W f , = wf.- WSc.
h-.
From this, it follows that wpj = wfj = 0 for all. j , j #
k
and w ? = W &>
0, wd . . = w t k > 0. Hence, proportionality is again realized with a constantc = W O . .
/w”
. .,
and the proof is complete.0 = wfiwfi
-
wfiwTi = w $(
7
w f i-
w i i ).
Since there should be at leastone j with wgi# 0, wTi =
-
w f i follows for all l,i, i #k and, thus,
Ai = 0 for Using this result, A, .= 0 implies that for all j,l,i, z #k
WO
WP
230 M. Z I E H E A N D H - R . GREGORIUS
all
i, i
#k.
Therefore, if one Ak is 0, then all Ai,i
= 1,..
.
,n must be 0, and selection acts symmetrically in the sexes. For the reverse direction, it is easily verified that symmetrical selectioii in the sexes, i.e., proportionality of the selec- tion coefficients between the sexes: implies that Ai = 0 for all i.A P P E N D I X C
In
an equilibrium pointP*,
for p i # 0,P f
I p ; { W p * ) - E ? ( P * ) } l = p , * ( l - p y L T S p * )
-
I__Ep
(P*>I
i#k,;,+o 1-p:
where q*(i,i,l) (referring to P * ) is as defined in APPENDIX A and A ? : = m a x
k k r , 3 , 1
I
w&
-
wFiI.
Then following from APPENDIX A, we havewhere L:, ( P ) : = ( 1 - 1
W Q
= o
u,u
I I v
A formula with analogously defined sex :
and L:. ( P ) can be derived for the male
G f ( P * ) - W $ ( P * ) 1