NONELECTROPHORETIC GENETIC VARIABILITY
IN
MOSQUITOES: POLYMORPHISMFOR
TEMPERATURE-RESISTANTAND
TEMPERATURE-SENSITIVE PHOSPHOGLUCOMUTASE ALLELES IN
CULEX
PIPIENS
R. SCOZZARI1, G. TRIPPAI, V. PETRARCAZ AND R. CICCHETTI'
Manuscript received November 17, 1978 Revised copy received June 18, 1979
ABSTRACT
Homogenates of single individuals of two natural populations and five laboratory populations of Culez pipiens were examined by combining electro- phoresis and heat denaturation studies on phosphoglucomutase (PGM)
.
AU populations showed a high degree of polymorphism for isoelectrophoretic temperature-resistant ( t r ) and temperature-sensitive ( t s ) alleles. Formal genetic data on the heat stability differences of the PGM are given. If both electrophoretic and isoelectrophoretic alleles are taken into account, the mean increase in the degree of heterozygosity is quite remarkable, i.e., about 65%.---The data are considered in relation to the biological significance that this new type of variability of structural genes could have in natural populations.G E N E T I C variability between and within populations of the same species is
a general phenomenon of the biological world. In particular, at the level of
proteins and enzymes, electrophoretic techniques have shown the existence of a high degree (about 30% of the structural loci) of genetic polymorphism in widely different organisms (LEWONTIN 1974; HARRIS 1975; AYALA 1976). Since it has been calculated that, on the average, of all possible amino acid substitu- tions in a protein, only one-third give rise to differences that can be identified by electrophoresis, much of the total variation in structural genes is still unex- plored (LEWONTIN 1974). One of the methods recently employed to detect elec- trophoretically cryptic alleles consists in comparing the heat stabilities of allelic enzymes. WRIGHT and MACINTYRE (1965),
TRIPPA,
LOVERRE and CATAMO(1976), SINGH, LEWONTIN and
FELTON
(1976), MILKMAN (1976), COCHRANE(1976), COYNE and
FELTON
(1977) and SAMPSELL (1977) have reported in Drosophila conclusive results showing that the differences observed in heat sta- bility depend on hidden structural variations in electrophoretically homogene- ous alleles. This has been demonstrated both at loci with an electrophoretic polymorphism and at electrophoretically monomorphic loci.This paper reports the existence of electrophoretically cryptic alleles expressed as differences in heat stability at the phosphoglucomutase locus (Pgm) in popu-
Centro di Genetica Evoluzionistica del C.N.R. and Istituto di Genetica, Facolt; di Scienze, Universit; di Roma, Italia.
* Istituto di Parassitologia, Universiti di Roma, Italia.
202 R. SCOZZARI et al.
lations of Culex pipiens. This species, which is geographically very widespread, is still the subject of numerous studies because of both its parasitological impor- tance and its complicated taxonomic situation. Besides morphological differences, there are in fact clear physiological differences between different populations of CuZex pipiens, such as anautogeny and autogeny (i.e., females are able to lay their first batch of eggs with or without a blood meal, respectively)
,
stenogamy (mating takes place even in restricted spaces) and eurigamy (for mating to occur, large spaces are necessary for the nuptial flight), mammophily and orni- thophily (females preferentially bite either mammals or birds) (RIOUX 1958).Formal genetic studies of the P g m electrophoretic variants in Culex pipiens
have shown the occurrence of six co-dominant alleles of an autosomal gene (BULLINI et al. 1973). These alleles were termed Pgm", PgmBs, PgmBs, PgmB1, PgmC and PgmD, in order of increasing mobility. The eight Italian populations studied by these authors were found to be polymorphic for three to six alleles, the most frequent in all samples being PgmB1.
MATERIALS A N D METHODS
The material examined consists of two natural populations and five laboratory populations of Cu2ex pipiens. The natural populations were collected as third-instar larvae and ninphae at Fogliano (FO), near Latina, and Sui0 (SU), near Caserta, i n epigean environments typical
of the rural form of the C. pipiens complex. Of the five laboratory populations, three were collected at Magliana (MA), near Rome, a t Latina (LA) and Rome (RO), respectively, in epigean environments characteristic of the urban form of the C . pipiens complex; the other two populations were collected at Kano (KA), in North Nigeria, and Benin (BE), i n South Nigeria, and were identified as Culex pipiens fatiguns.
All the laboratory populations were started from collections of about 2000 eggs o r larvae and ninphae. The effective founder size of the populations LA and RO was a t F, about 100 egg-laying females. These populations were subsequently maintained at about 500 individuals per generation and examined at the 80th generation. The populations MA, KA and BE fell a t
F, to about 20 egg-laying females and were then maintained for 20, 30 and ten generations at about 500, 1000 and 300 individuals per generation, respectively. The larvae and ninphae were mass reared and kept at room temperature (ranging between 15" and 20") in plastic bowls (50X25XlOcm). The food consisted of a few mg per day of fish food. The adults were kept in cages and supplied with sucrose solution for one o r two days before being electrophoresed.
The technique used for electrophoretic analysis and heat denaturation is that described by TRIPPA et al. (1978a) for single-fly homogenates of Drosophila melanogmter. After electro- phoresis of single-fly homogenates, the gel is cut into two slices. One is incubated at 37" for
15 min to develop the electrophoretic pattern and the other at 60" for 15 min for heat treat- ment and determination of the heat-sensitivity behavior of different allozymes. After incuba- tion the slices are stained following the method of SPENCER, HOPKINSON and HARRIS (1964). The temperature and time intervals for heat treatment were chosen so as to permit a non- ambiguous detection of heat-resistant bands (PGM activity is present after the 60" incubation) and heat-sensitive bands (no PGM activity o r very faint PGM activity after incubation at 60" vc.ith heat-resistant bands fully developed).
RESULTS
Five out of the six electrophoretic alleles previously described (BULLINI et al.
Culex pipiens
-
origin1 2 3 4 5 6 7 8 a i 6 5 4 3 2 1
(a1 (bl
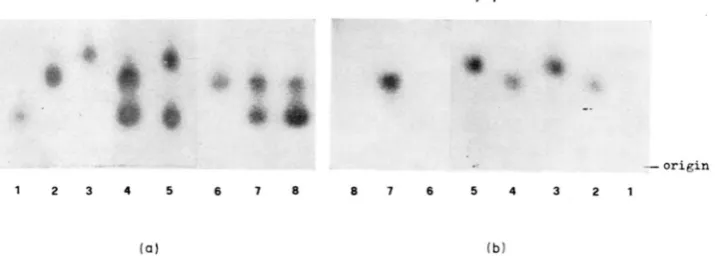
FIGURE 1 .-Electrophoretic and heat sensitivity phenotypes of homogenates of single individ- uals of Culez pipiens on the two slices of the same gel. One slice was incubated at 37" (a) and the other at 60" (b) for 15 minutes before being stained for PGM. The following phenotypes are shown: (1) PGM 0.80,ts; (2) PGM 1.00,tr; (3) PGM 1.10p; (4) PGM 0.80.t~-1.00,tr; (5) PGM O.SO.ts-1.10,tr; (6) PGM 1.00,ts; ( 7 ) PGM 0.801ts-1.00,tr and (8) PGM 0.80,ts-l.OO,ts.
and Pgm'.zo, respectively, following the criterion already adopted for the PGM system in Drosophila melanogaster (TRIPPA et al.
1977).
Figure 1 shows thetwo mirror slices of the same starch gel with some of the PGM electrophoretic phenotypes as visible after treatment at 37" (a) and a t 60" (b). It can be seen that, in some individuals with one (#1 and 6) PGM electrophoretic band, this band is inactivated after treatment a t 60". Moreover, the individuals 2 and 6
and 4 and 8 show that bands with identical electrophoretic mobility can display different heat-sensitivity behaviors. From this figure it can also be seen that the heterozygotes 4,5 and 7, with two electrophoretic bands at
37",
display only one band inactivated after heat treatment. No gross differences in the enzymatic activities of the two allele products of the heterozygotes with one of the two bands inactivated at 60" resulted from the visual scoring of the electrophoreticTABLE 1
Testing the Mendelian segregation of the electrophoretic and heal sensitiuity phenotypes of Pgm0*8o,fs, p-l.oo.fr, pgm1.oo.t~ and p g " . i o . i r alleles
Type of cross F, Phenotype S O . ' F2 I'heno!ype No.
_ _ _ _ ~
PGM 1.00,tr 38
(1) P p m l.OO,/r X Pgm l.ZO,tr PGNI l.OO./r-l.lO,/r Si PGM [email protected],fr 79 PGM 1.10,tr 36 52 PGM 1.00,ts
(2) P p m 1.00./s X P g m Z.lO,/r PGM l.OO,ts-l.lO,fr 84 PGAM l.OO,/s-1.10,tr 108 PGM 1.10,tr 61
34 PGM 0.80.ts
(3) Ppm O.SO,ts X Pgm l.OO,/r PGM 0.80,rs-l.O0,tr 68 PGM 0.8O.ts-1.00,tr 77
PGM 1 .OO,tr 30
43 PGM 0.80,is
(4) P p m O.SO,/s X Pgm l.lO,/r PGM 0.80,/s-l.OO,tr 72 PGM 0.80,ts-l.lO,tr 103
40
PGiM 1.10,tr
204 R. SCOZZARI et al.
zymograms obtained at 37". Since some individuals heterozygous for PGM elec- trophoretic alleles have two bands displaying clearly different thermosensitivi- ties, it can be argued that the genetic basis of the difference in heat stability must lie in the structural Pgm locus. Formal genetic data on the heat-stability differ- ences in PGM were obtained from a series of crosses between individuals from
four lines characterized with regards to PGM by both electrophoresis and heat- denaturation behavior (see Table 1). The data in Table 1 show that the heat- sensitivity phenotype of each PGM band goes along with the electrophoretic phenotype of this same band and is limited to it. Therefore, we conclude that the property of sensitivity depends on the Pgm structural gene (unless we consider the remote possibility that it depends on an allele-specific linked modifier).
TABLE 2
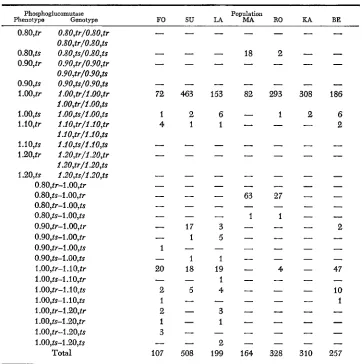
Distribution of PGM electrophoretic and heat sensitivity phenotypes in seven populations of Culex pipiens
Phosphoglucomutase Population
FO SU LA MA RO RA BE
Phenotype Genotype 0.80,tr O.XO,tr/O.XO,tr
O.XO,tr/O.XO,ts
0.80,fs O.XO,ts/O.XO,ts 0.90,tr 0.90,tr/0.90,tr 0.90,tr/0.90,ts 0.90,fS 0.90,fs/0.90,ts l.OO,tr 1.00,tr/1.007tr
1.OO7tr/1.0O,ts
1 .oo,ts 1.oo,ts/1.oo,ts l.iO,tr Z.107tr/1.10,tr
1.1O,tr/l.1O7ts 1.1 0,ts l.1o,fs/1.1o,ts 1.20,tr 1.20,tr/1.20,tr 1.207tr/1.20,ts 1.20,ts 1.20,ts/1.20,ts
0.80,tr-1 .OO,tr 0.80,ts-l.OO,tr 0.80,tr-1 .OO,ts 0.80,ts-1 .oo,ts 0.90,fr-1.00,tr 0.90,ts-1 .OO,tr 0.90,tr-I .OO,ts 0.90,fS-1 .oo,ts 1.00,tr-1.10,tr l.OO,ts-l.IO,tr 1 .OO,tr-l .lo$ 1.oo,fs-1.1o,ts 1 .OO,tr-l.2O,tr l.OO,ts-l.20,tr 1.00,tr-1.207ts 1.00,ts--1.20,ts
I-rEm ALLELES pipiens
In the following, we report the findings of studies on the
PGM
of natural and laboratory populations of Culex pipiens with respect to both electrophoretic and thermosensitivity phenotypes (Table 2 ) . As far as the P g m electrophoretic alleles are concerned, the estimates of gene frequencies were obtained by direct count for every co-dominant allele. In all samples, the Hardy-Weinberg equi- librium was verified. On the contrary, direct counts of tr and ts alleles were not possible because in individuals with a singlePGM
electrophoretic band, tr and ts behave as dominant and recessive alleles, respectively. Therefore the estimateof the cumulative ts gene frequency has been obtained in each population by assuming Hardy-Weinberg equilibrium for the tr and ts alleles. Depending on whether the population under study was electrophoretically monomorphic or polymorphic, this was calculated as either the square-root of the frequency of individuals with a temperature-sensitive phenotype or one minus the sum of the frequencies of all tr alleles. The frequency of each of the tr alleles was in turn estimated by the general formula
pAi = 1
-
Anon-i/Anon--i,
where A4 is the ith electrophoretic tr allele and Anon-c/Anon-i the frequency of individuals with no
Ai
allele. For example, in the Rome population the estimateof the Pgml.OO.tr frequency was obtained as follows:
1
-
d
( P g m 0.80,ts)f
( P g m 1.00,ts)+
( P g m O.8O,ts-~.OO,ts) ==
1-0.11
=0.89
-
1/
2+lfl
328
and
Pgml.lojtr = 1
-
.\/
( P g m 0 . 8 0 ~ ~ )4-
( P g m 1 .OO,tr) 4- ( P g m O.80,ts-I .OO,tr)+= 0.006.
2+293+1+27+1
328 ( P g m 080,ts-2.00,ts) = 1
-
d
Thus,
in this population, the estimate of the frequency of all P g m thermo- resistant allelesf ( P g m t T ) was f (Pgmz*ooJT) -I- f (Pgml.’o*t‘) =
0.896
and f ( P g m t s )=
1
-
[f(Pgml.OO*tr) f(Pgml.lo*tr)]1
-
(0.890)
4-
0.006)
= 0.104.The frequencies of the various Pgmts alleles, grouped according to their electro- phoretic behavior, were obtained by subtracting the frequency of the corres- ponding tr allele from the cumulative frequency of that same electrophoretic allele (for example, if f(Pgm1*O0) was the frequency of the Pgml-OO allele and f(Pgml.OO~tr) that of the Pgml.OO,tT allele, the frequency of the Pgml.oo.ts allele was estimated as f(Pgml.OOJs)
=
f(Pgmleoo)-
f(Pgml.oo*tr) ).
The frequencies of the rare alleles were calculated by directly counting the heterozygotes.Culex pipiens
DISCUSSION
The present findings reveal a hidden variation at the Pgm locus of Culex pipiens in the form of heat-sensitive alleles. The phenomenon observed, inactiva- tion of one band and not of another, seems to be due to true heat-stability differ- ences. If this were not so, we should expect to find an asymmetrical pattern at 37" in those heterozygotes with only one of the two bands inactivated at 60".
In the present screening, no such pattern was detected (cf., Figure 1). This finding is in contrast with that reported for the a-glycerophosphate dehydro- genase locus (a-Gpdh) in Drosophila melanogaster by
BEWLEY
(1978). Hereports that spectrophotometric analysis of all 11 putative thermolability vari- ants demonstrated that these were not true heat-stability variants, but rather variants in total enzyme activity.
Tables 2 and 3 show that in Culex pipiens all Pgm electrophoretic alleles detected-with the exception of the PgmO.so allele, which was regularly found to be heat sensitive-can be split into at least two alleles differentiated on the basis of their heat stability. More important, there is an increase in the number of polymorphic alleles because most of the common electromorphs consist of two alleles, these in turn polymorphic. The estimate of genetic heterogeneity f o r this locus considerably increases if the heterozygosity values for each population are compared by using either the electrophoretic alleles alone or the electro- phoretic and heat-sensitivity alleles together (Table 3). In particular, the Kano population, which is monomorphic from an electrophoretic point of view, is polymorphic according to the thermosensitivity criterion. To date, this is the second case in mosquitoes of an electrophoretically monomorphic gene that, if
criteria other than electrophoresis are adopted, has been shown to be effectively polymorphic (the first was found in Culiseta titorea; SCOZZARI, CICCHETTI and
TRIPPA 1977).
Unlike previous observations concerning natural populations of Culex pipiens
(BULLINI et at. 1973) the data in Table 3 show a certain degree of heterogeneity in the distribution of gene frequencies in the pcpulations examined, as deter- mined both by electrophoretic analysis alone and by combining the two methods. In fact, apart from the Pgml.oo~tr allele, which is the most frequent allele in all the populations examined, different alleles are polymorphic in different popula- tions. Of course, founder effect and drift during culture could have played an important role in determining the pattern of distribution of allele frequencies in the laboratory populations. In particular, this is probably true for the popula- tions MA, KA and
BE,
which started from a small number of founder females.The pattern of distribution of gene frequencies both at the intra- and the inter- specific level can be used to test the validity of different theoretical models on the biological importance of protein polymorphisms in natural populations. The resolution of putative homogeneous alleles into distinct classes of alleles at a polymorphic frequency can have important implications with respect to these theories (KING 1974). For example, BERNSTEIN, THROCKMORTON and HUBBY
208 R. SCOZZARI et al.
electrophoretic alleles are not. They suggested that the latter are maintained by balancing selection, while the former are selectively neutral and drift in local populations. COCHRANE (1976), on the other hand, reported that the relative constancy of the gene frequencies for Esterase-6 (Est-6) in natural populations of Drosophila melanogaster, obtained on the basis of electrophoretic mobility alone, is not altered when the criterion of heat stability is applied. However, it was not possible to draw any conclusions as to the evolutionary significance of the variation at the Est-6 locus. From our data, it appears that in Culex pipiens
phosphoglucomutase alleles are geographically differentiated with regard to both electrophoretic and thermostability behavior.
Another approach to the problem of the biological importance of protein polymorphisms is to identify those differences in protein properties that are likely to reflect in themselves functional differences in vivo. A correlation could then be sought between those differences and habitat factors differing in ways expected to affect the functioning of the protein. One variation, which could in itself be of physiological importance, is that relative to heat stability as described
in this paper. For example, it may be that a greater heat stability in vitro at a temperature higher than the physiological one experienced for a short time corresponds to a greater heat stability in vivo at physiological temperatures experienced for a longer time. This could be important from a selective point
of view. Data obtained in six species of Aedes and Culiseta (Culicidae, Diptera) indicate the existence of a strong correlation between the frequency of phospho- glucomutase heat-sensitive alleles and the temperature of the environment where larvae develop, suggesting that different selective values are associated with the
PGM thermoresistant and thermosensitive genotypes in the different habitats
(TRIPPA
et al. 1978b).populations and A. MICHELI for excellent technical assistance.
We thank A. SCIROCCHI (Centro Antimalarico, Latina) for providing samples from natural
LITERATURE CITED
AYALA, F. J., 1976
BERNSTEIN, S. C., L. H. THROCKMORTON and J. L. HUBBY, 1973
BEWLEY, G. C., 1978
BULLINI, L., M. COLUZZI, A. P. BIANCHI BULLINI and L. RENNA, 1973
(Editor) Molccular Evolution. Sinauer Associates, Sunderland, Massachu- setts.
Still more genetic variability in natural populations. Proc. Natl. Acad. Sci. U.S. 70: 3928-3931.
Heat stability studies at the a-glycerophosphate dehydrogenase locus in populations of Drosophila melanogaster. Biochem. Genet. 16: 769-775.
Stability of frequencies of phosphoglucomutase alleles i n Culex pipiens breeding in ecologically different environ- ments. Accad. Naz. Lincei 53: 154-157.
Heat stability variants of esterase-6 in Drosophila melanogaster. Nature
Genic heterogeneity at two alcohol dehydrogenase loci
The Principles of H u m a n Biochemical Genetics, North-Holland, Amsterdam. COCHRANE, B. J., 1976
263: 131-132.
COYNE, J. A. and A. A. FELTON, 1977
i n Drosophila pseudoobscura and Drosophila persimilis. Genetics 87: 285-304.
HARRIS, H., 1975
HEAT SENSITIVE ALLELES IN
Culex
pipiensLEWONTIN, R. C., 1974 The Genetic Basis of Evolutionary Changes, Columbia University Press, New York.
MILKMAN, R., 1976 Further evidence of thermostability variation within electrophoretic mobility classes of enzymes. Biochem. Genet. 14: 383-387.
RIOUX, J. A., 1958 Les Culicides du ”Midi“ Mediterraneen. Encyclopedie Entomologique.
35: 244-260.
SAMPSELL, B., 1977 Isolation and genetic characterization of alcohol dehydrogenase thermo- stability variants occurring in natural populations of Drosophila melanogaster. Biochem. Genet. 15: 971-988.
SCOZZARI, R., R. CICCHETTI and G. TRIPPA, 1977 Hidden phosphoglucomutase (PGM) genetic polymorphism in a natural population of Culiseta litorea (Diptera, Culicidae). Parassitologia
19: 103-107.
SINGH, R. S., R. C. LEWONTIN and A. A. FELTON, 1976 Genetic heterogeneity within electro- phoretic “alleles” of xanthine dehydrogenase in Drosophila pseudoobscura. Genetics 84.:
SPENCER, N., D. A. HOPKINSON and H. HARRIS, 1964 Phosphoglucomutase polymorphism in
man. Nature 204: 742-743.
TRIPPA, G., A. CATAMO, A. LOMBARDOZZI and R. CICCHETTI, 1978a A simple approach for dis- covering common nonelectrophoretic enzyme variability: A heat denaturation study in
Drosophila melanogaster. Biochem. Genet. 16: 299-305.
TRIPPA, G., G. A. DANIELI, R. COSTA and R. SCOZZARI, 1977 A new allele at the Pgm locus i n Drosophila melanogaster. Drosophila Inform. Serv. 52: 74.
TRIPPA, G., A. LOVERRE and A. CATAMO, 1976 Thermostability studies for investigating non. electrophoretic polymorphic alleles in Drosophila melanogaster. Nature 260 : 4244.
TRIPPA, G., R. SCOZZARI, R. CICCHETTI, V. SEPE and R. CAPPELLATO, 1978b Variabilig genetica isoelettroforetica in alcune specie di Culicini (Culicidae, Diptera). Atti Ass. Genet. Ital. 23:
243-246.
Heat-stable and heat-labile esterase-6F enzymes in Drosophila melanogaster produced by different Est-6F alleles. J. Elisha Mitchell Sci
Soc. 81: 17.
Corresponding editor: D. L. HARTL
609-629.