Electrically Evoked Dopamine Release in Virgin and Postpartum Rats:
Lack of Modulation by Nasal Oxytocin
Kyla Mace
The University of North Carolina at Chapel Hill
Spring 2016
A thesis presented to the faculty of The University of North Carolina at Chapel Hill in partial
fulfillment of the requirements for the Bachelor of Arts degree with Honors in Psychology.
Donita Robinson, PhD (Advisor)
Josephine Johns, PhD
Acknowledgments
I would like to thank my advisor Dr. Donita Robinson and co-advisor Dr. Josephine
Johns for their continuous support, encouragement, and advice. I would also like to thank Dr.
Tatiana Shnitko for her expert training in voltammetry and surgery and her patience in answering
my countless questions and concerns. Our lab manager Kaitlin Sullivan was of great help to me,
coming in many early mornings to help with rat breeding and cocaine injections. All of my
labmates deserve credit for providing moral support throughout the year. Finally, I am always
grateful to my parents for their unending support and love. This research was supported in part
Abstract
The oxytocin (OT) and dopamine (DA) systems are highly intertwined and have been implicated
in a number of similar behaviors, particularly social, sexual, and maternal behaviors. However,
the direct pharmacological effect of OT administration on the DA system is yet to be determined.
Additionally, although nasal OT is the primary route of administration in human research, it is
rarely used in animal studies, with researchers often relying on other routes for systemic OT
administration (e.g., intraperitoneal injection). The present study aims to fill this gap in the
literature by examining the effect of nasal OT on the DA system using fast-scan cyclic
voltammetry. Specifically, stimulated DA release in the nucleus accumbens was measured before
and after nasal OT administration in female rats. Because the peripartum period is rife with
neuroendocrine changes, both virgin and postpartum animals were used to examine how
pregnancy affects OT-DA interactions. However, no effect of OT was found, which is
inconsistent with past research and suggests an indirect mechanism by which OT affects DA and
DA-driven behaviors. Parity status was found to have a significant effect on baseline levels of
evoked DA release, with postpartum animals showing larger concentrations of DA compared to
virgin females. This corroborates past studies finding alterations in the DAergic system
Electrically Evoked Dopamine Release in Virgin and Postpartum Rats:
Lack of Modulation by Nasal Oxytocin
Our drive to form social bonds between friends, family, and romantic partners relies on a
complex neural circuitry, partially involving the oxytocin (OT) and dopamine (DA) systems.
These two neurochemicals are highly involved in motivational behavior, and their interaction has
been implicated in social-cognitive processing. However, the details of this interaction are still
under investigation, and OT’s effect on motivational behavior (and the DAergic system by
extension) seems to vary depending on the context. Although DA appears to be involved in all
types of motivational cues, OT seems to enhance the salience of social cues in particular. It has
been hypothesized that the dysregulation of these two systems can lead to maladaptive behaviors,
such as drug abuse and child neglect, where naturally motivational social cues are devalued and
drug cues are overvalued. In this paper, I will first provide a brief overview of the OT and DA
systems, their circuitry, and the behaviors in which they are involved, followed by a description
of how the two systems are connected. Next, I will relate animal findings to human research and
provide a brief introduction to fast-scan cyclic voltammetry, the electrochemical technique I will
use to measure DA release. Finally, I will introduce my general plan for investigating the
interaction of the OT and DA systems.
Oxytocin and Dopamine: Circuitry and Interaction
OT exerts a wide range of effects, both behavioral and biological, likely due to its ability
to act as both a neurotransmitter in the nervous system and a hormone throughout the body. On a
physiological level, it plays a fundamental role in lactation, erection, ejaculation, and triggering
labor (Gimpl, Fahrenholz, & Gene, 2001). In the central nervous system, OT has been implicated
caregiving (for review, see Lee, Macbeth, Pagani, & Scott Young, 2009). This range of effects
may be due to OT’s widespread presence throughout the brain. OT is primarily synthesized in
the paraventricular nucleus (PVN) and supraoptic nucleus (SON) of the hypothalamus (Gimpl et
al., 2001). From there, the SON primarily projects to the pituitary to release OT into the
bloodstream, but the PVN has a wide range of projections, including the olfactory bulb, medial
preoptic area (MPOA), nucleus accumbens (NAc), amygdala, hippocampus, and ventral
tegmental area (VTA) (Rutherford, Williams, Moy, Mayes, & Johns, 2011).
OT has been linked with a number of neurochemical systems, including serotonin
(Marazziti et al., 2012), nitric oxide (da Silva, Cedraz-Mercez, & Varanda, 2014), and GABA
(Lonstein, Maguire, Meinlschmidt, & Neumann, 2014). Additionally, OT and DA have been
found to share some circuitry and are involved in similar behaviors. DA plays a fundamental role
in motivational behaviors, including reward learning (Day, Roitman, Wightman, & Carelli,
2007), drug administration (Di Chiara et al., 2004), social and sexual behavior (Brom, Both,
Laan, Everaerd, & Spinhoven, 2014; Insel, 2003), and maternal caregiving (Love, 2014). A
major source of DA is the VTA, which projects to a number of forebrain structures, including the
olfactory tubercle, prefrontal cortex, and NAc, the last of which being the main focus of the
present study. The NAc has been strongly implicated in motivational behavior, and the amount
of effort an animal will exert to achieve a reward is dependent on extracellular DA in the NAc
(for review, see Salamone, Correa, Farrar, & Mingote, 2007). In addition, NAc DA transients, or
burst firing of DA neurons, are thought to be important in cue-reward associations. Through the
course of Pavlovian conditioning, where an unconditioned stimulus (e.g., a light) is presented
before an unconditioned reward (e.g., a food pellet), DA transients shift from occurring after the
predictive value of the cue (Day et al., 2007). NAc DA also plays a fundamental role in maternal
behavior, especially appetitive behaviors (goal-directed behaviors), such as pup retrieval, which
is impaired after administration of a DA antagonist to the NAc (Keer & Stern, 1999). DA release
in the NAc has also been found during pup licking/grooming (Champagne et al., 2004) and in
response to pup cues (cries and odors) in hormonally primed virgin females (Afonso, Shams, Jin,
& Fleming, 2013).
The DAergic and OTergic systems are interrelated in a number of ways. The PVN
projects to both the VTA and the NAc, and the MPOA sends strong projections to the VTA (for
review, see Numan & Stolzenberg, 2009). Both the VTA and NAc express OT receptors, and
many OT cells in the MPOA, PVN, and SON express DA receptors in return, indicating a
reciprocal relationship between the two systems (Baskerville, Allard, Wayman, & Douglas,
2009; Vaccari, Lolait, & Ostrowski, 1998). OT receptors are not found on the DAergic cells in
the VTA, however, indicating that their relationship may be indirect (Yoshida et al., 2009). Even
so, their neurobiological interaction is plausible, given that both neurochemicals have been
implicated in similar behaviors, including social, sexual, and maternal behavior.
OT’s influence on the DA systems can be facilitatory or inhibitory, depending on the
context. For example, infusing DA into the subthalamic nucleus of the hypothalamus enhanced
conditioned place preference, but adding OT abolished this effect (Baracz & Cornish, 2013).
This suggests that OT in this area inhibits the motivating effect of DA in the conditioned place
preference task. Additionally, OT has been found to suppress some effects of drugs of abuse (for
review, see McGregor & Bowen, 2012). Peripheral OT administration has been shown to reduce
the locomotor effects of methamphetamine and cocaine, both of which increase DA release and
These effects may result from OT’s inhibitory effect on the DA system, given that OT reduced
methamphetamine-induced activity in the NAc (Carson et al., 2010) and inhibited the increased
DA utilization found after cocaine administration (Kovàcs et al., 1990).
However, in other contexts, OT appears to exert an excitatory effect on the DA system
and its related behaviors. Although adding OT diminishes a DA-induced conditioned place
preference, the same effect is not found in a conditioned partner preference task (Triana-Del Rio
et al., 2015). In this experiment, male rats were co-housed with a same sex partner and received
saline, a DA receptor agonist, or a combination of a DA receptor agonist and OT. When later
presented with either their same-sex partner or a sexually receptive female, both the DA and
DA/OT animals preferred their male partner, whereas saline animals preferred the female,
suggesting an alteration in either social or sexual behavior. In this case, adding OT did not
eliminate the partner preference like it did for conditioned place preference. In terms of maternal
behavior, administering OT to virgin female rats increases pup-directed behaviors, such as
licking, nursing, pup retrieval, and nest-building, all behaviors that rely on DA (Fahrbach, 1984).
Infusing an OT antagonist into the VTA of postpartum dams reduced DA release in the NAc
during pup interaction, suggesting that pup-induced DA release is dependent on OT (Shahrokh,
Zhang, Diorio, Gratton, & Meaney, 2010).
It seems as though OT’s influence on DA depends on the type of reward involved,
possibly due to parallel brain systems used for processing non-social and social rewards. Either
reward system can be strengthened by exciting the DAergic system, either by administering a
DA agonist or drugs of abuse that act on the DA system, like methamphetamine and cocaine.
However, the OTergic system appears to devalue non-social rewards and increase the salience of
drugs of abuse, OT appears to have an inhibitory effect on the DA system and the behaviors
involved. However, for social and/or sexual rewards, such as a partner preference and maternal
behavior, OT appears to exert an excitatory effect on the DA system. Although there is a wealth
of research investigating OT-DA interactions in awake, behaving animals, much less is known
about the basic pharmacological effect of exogenous OT on the DA system.
Oxytocin Administration in Animal and Human Studies
Animal studies tend to use infusion or injection to administer OT, but these techniques
are too invasive for human studies. Instead, intranasal OT has been used widely in the human
literature to modulate social functioning. Intranasal OT may increase the salience of socially
relevant cues, including positively and negatively charged faces (Groppe et al., 2013), infant
laughter (Riem et al., 2012), and both infant and sexual images (Gregory, Cheng, Rupp,
Sengelaub, & Heiman, 2015). Additionally, intranasal OT has been found to improve symptoms
of conditions that impair social functioning, such as schizophrenia (Feifel et al., 2010) and
autism (Domes et al., 2013; Guastella et al., 2010). Finally, as mentioned earlier, intranasal OT
has been proposed as a possible treatment for drug addiction, due to its ability to reduce the
salience of drug cues (for review, see McGregor & Bowen, 2012).
Because intranasal OT is so widely used in human studies, its use in animal studies would
allow for a more accurate translation of animal findings to human research. Additionally,
intranasal administration has some practical benefits that other administration routes lack.
Intranasal administration allows OT to enter the central nervous system through the olfactory
epithelium, largely bypassing the blood-brain barrier, which can prevent as much as 99.998% of
subcutaneously administered OT from reaching the central nervous system (Mens, Witter, & Van
of rats, where it has only a 2 minute half-life. However, in the cerebrospinal fluid, its half-life
can be extended to up to 19 minutes (Mens et al., 1983). For these reasons, intranasal OT may be
more effective in elevating central OT levels for extended periods of time.
Although it is not commonly used in animal studies, intranasal (in non-human primates)
and nasal (in rodents) OT appears to exert similar effects as other routes of administration. As
with injections and intracerebral infusions, nasal OT has been found to increase central and
peripheral OT levels in rats and mice, although the exact route by which this occurs is unclear
(Neumann, Maloumby, Beiderbeck, Lukas, & Landgraf, 2013). Research on the behavioral
effects of intranasal OT in animals appears to mirror findings from the human literature.
Intranasally applied OT reduced the hypothalamic-pituitary-adrenal axis stress response to social
isolation in squirrel monkeys (Parker, Buckmaster, Schatzberg, & Lyons, 2005) and facilitated
social bonding in marmoset monkeys (Smith, Ågmo, Birnie, & French, 2010). However, little
research has been performed investigating how intranasal or nasal OT affects the DA systems in
animals.
Fast Scan Cyclic Voltammetry
The present study examines how nasal OT affects NAc DA release, using fast-scan cyclic
voltammetry (FSCV). FSCV allows for a higher spatial and temporal resolution than other
techniques such as microdialysis. This electrochemical technique involves applying a range of
electrical potentials to an electrode placed in the brain region of interest; as the chemicals around
the electrode undergo oxidation-reduction reactions at the various potentials, the changes in
current are measured to deduce the concentration of specific chemicals (Robinson, Venton,
Heien, & Wightman, 2003; Robinson & Wightman, 2007). For DA, the waveform of the applied
at 10 Hz throughout a recording session. The applied potential creates a large background
current, which can be subtracted from the recorded voltammograms to reveal smaller changes in
current. Because compounds have specific potentials at which they oxidize or reduce, the
background-subtracted voltammogram can be identified as a specific molecule, allowing for
measurement of the chemical of interest. Specifically, DA oxidizes to its ortho-quinone at
approximately 0.6 V and the ortho-quinone reduces back to DA at approximately -0.2 V
(Robinson & Wightman, 2007). This exact pattern is not shared with other compounds such as
serotonin or nitric oxide. However, norepinephrine oxidizes and reduces at the same potentials as
DA, making it important to consider the anatomical position of the electrode when identifying
norepinephrine and DA using voltammetry (Robinson & Wightman, 2007).
FSCV can be used in freely moving rats to measure real-time neurochemical changes as
the animal behaves and interacts with its environment, but it can also be used in anesthetized
animals to answer pharmacological questions. To measure DA release in the NAc in anesthetized
animals, a stimulating current is applied to the VTA and the resulting DA release is measured at
its terminals in the NAc. In this way, the neurobiological effects of a pharmacological agent
(such as OT) can be isolated from its behavioral components and environmental influences can
be eliminated.
Project Plan
To investigate OT’s interactions with the DA system, electrically stimulated DA release
in the NAc was measured before and after nasal OT administration (or its vehicle as a control).
Virgin females and postpartum dams were used to determine whether pregnancy alters OT’s
effect on NAc DA, given that the neuroendocrine system, OT included, undergoes a wide array
increases DA release in the NAc. Past studies have found that infusing OT into the VTA
increases DA release in the NAc of awake animals, in both male and female animals (Melis et
al., 2007; Shahrokh et al., 2010), but whether this occurs after nasal OT in anesthetized animals,
as well as postpartum dams, is yet to be established.
Methods Animals
Virgin female (250-430g) Sprague-Dawley rats (Charles River, Raleigh, NC) were pair
housed on a 12-hour light-dark schedule. Female rats to be bred were single housed on a reverse
12-hour light-dark cycle (lights off at 0900) for a 2-week habituation period, after which they
were housed with breeder males. Gestation day (GD) 0 was designated as the day a sperm plug
was found. Pregnant females were weighed daily throughout their gestation. On GD 1, females
were returned to single housing and kept under the reverse light-dark cycle until GD 7, when
they were moved to a room with regular light cycle (lights on at 0700) for the rest of the
experiment. This procedure generally causes dams to deliver during daytime hours on GD 21
(Mayer & Rosenblatt, 1998). On delivery day (postpartum day (PPD) 0), litters were culled to 4
males and 4 females. The final weights of postpartum animals ranged from 213 to 323g. All rats
received water and food ad libitum. All procedures were conducted under federal and
institutional animal care and use committee guidelines for humane treatment.
Surgery
Virgin females underwent surgery at least one week after arriving in the animal facility, and
postpartum dams underwent surgery on PPD 1. Litters from dams were euthanized 30 minutes
prior to the dams’ anesthesia administration to remain consistent with past studies from the Johns
conducted. Rats were anesthetized (1.5g/kg urethane, i.p.) and placed in a stereotaxic frame.
Electrodes were implanted as previously described (Shnitko & Robinson, 2014); electrode
placements (from bregma) were -5.0mm and ML -1.2mm for the stimulating electrode and AP
+1.7mm and ML +1.8mm for the carbon fiber electrode. The stimulating electrode (bipolar,
parallel, stainless steel, 0.2 mm diameter/tip; Plastics One, Roanoke, VA) was initially placed in
the VTA at DV -8.0mm, and the carbon fiber electrode (length 60-115μm) was initially placed in
the NAc core DV -6.0mm. Both electrodes were lowered to optimize DA signal, with final
placements ranging from -8.1mm to -9.0mm for the stimulating electrode and -6.0mm to -6.3mm
for the carbon fiber electrode. A reference electrode (Ag/AgCl) was placed in the left hemisphere
and secured with a stainless-steel screw and dental cement.
Voltammetry and Experimental Procedure
The applied potential was ramped from -0.4 V to +1.3 V and back to -0.4 V every 100
ms. A stimulating current was applied to the VTA (24 biphasic square-wave pulses, 2 ms per
phase, 260.3 – 304 μA, 60 Hz); the stimulation current was adjusted to optimize the DA signal
and varied from animal to animal. Three evoked signals, collected 5 minutes apart, were
measured before OT or vehicle administration to determine the animal’s baseline level of
stimulated DA release. Following nasal treatment (10 μL per nostril of 1 mg/ml OT or its
vehicle), 12 evoked signals were collected five minutes apart, totaling an hour of voltammetric
recording. This time frame was chosen because brain OT levels peak at approximately 30 – 60
minutes after either intraperitoneal or nasal OT administration (Neumann et al., 2013). In order
to convert current to DA concentration, electrodes were calibrated in vitro post-experiment, as
previously described (Robinson & Howard, 2009). In short, current was recorded from each
concentration of DA (1μM). Electrodes from 10 animals were preserved for calibration, and
calibration factors (μM/nA) for the remaining electrodes were estimated from these calculations.
Drug Preparation
1mg/mL OT solution was prepared by dissolving synthetic OT (Bachem Americas, Inc.,
Torrance, CA) in a saline and 0.1% acetic acid solution, as OT requires a mildly acidic
environment in order to dissolve. Aliquots were kept frozen at -80°C until the time of the
experiment.
Analysis
Voltammetric analysis. Before analysis, data files were renamed in order to mask the time point at which the file was taken (i.e., before or after drug administration) to reduce researcher
bias during analysis. Voltammetric data was analyzed as previously described (Shnitko &
Robinson, 2014). DA was identified in the voltammetric current using color plots and cyclic
voltammograms. An average background voltammogram was taken from the 5 seconds
preceding stimulation and was subtracted from voltammograms taken during and after
stimulation to create background-subtracted voltammograms. No 2D smoothing was applied to
the data. Current-versus-time traces were used to determine peak current (nA) for each
stimulation. Calibration factors collected post-experiment were used to convert peak current to
peak concentration (μM), which will be referred to as [DA]max.
Dopamine signal modeling. To investigate whether DA release and uptake differed between virgin and postpartum groups, DA release and clearance in baseline recordings were
examined as previously described (Shnitko, Spear, & Robinson, 2015) using the
Michaelis-Menten kinetic equation (Wu, Reith, Wightman, Kawagoe, & Garris, 2001). This equation
pulse of stimulation ([DA]p, nM), the frequency of electrical stimulation (f, Hz), the velocity of
DA transporters (Vmax, nM/sec), and the affinity of those transporters (Km, nM). For each
baseline recording, a model of the current-versus-time plot was generated using this equation. Km
was held at 200nM and [DA]p and Vmax were modified until the model resembled the
experimental data, with a correlation coefficient of at least 0.9. For each animal, average [DA]p
and Vmax measurements were calculated from the three baseline files and were then compared
between virgin and postpartum groups, collapsing across nasal treatment groups.
Statistical analysis. To examine maximum evoked DA concentration over the course of the experiment, each rat’s 15 data points were condensed to five time points by averaging [DA]max
from three consecutive files at a time, leaving each rat with five total measurements: one baseline
measurement and four post-administration measurements. To examine how postpartum status
affects the concentration of DA release, concentration of DA released per pulse of stimulation,
and the velocity of DA transporters, three Mann-Whitney Rank Sum tests were performed (due
to non-equal variance between groups), comparing [DA]max, [DA]p, and Vmax between virgin and
postpartum groups, collapsing across nasal treatment groups.
To examine changes in DA signal following OT treatment, the data for each rat was
normalized to baseline by calculating the percent change from baseline for each of the four
post-administration measurements. A two-way repeated measures ANOVA was performed with
percent change from baseline as the dependent variable, group as a between-subjects factor
(OT/virgin, vehicle/virgin, OT/postpartum, vehicle/postpartum), and time as within-subjects
Results Baseline Dopamine
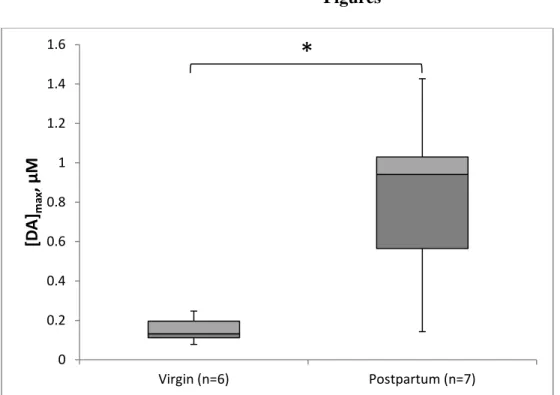
To determine how parity affects evoked DA release, we examined [DA]max at baseline in
virgin animals (n=6) and postpartum dams (n=7), collapsing across nasal treatment groups. Of
the postpartum dams, five were run the day after delivery (PPD 1), and two were run on PPD 3.
All animals included in this analysis received 2.24-2.26V stimulation (289-304μA); two animals
(both OT-treated virgin females) received 2.2V (260.3μA) stimulation and were removed from
this analysis. The baseline concentration of evoked DA release in postpartum dams (Mdn=0.941
μM) was over five times larger than virgin females (Mdn=0.123 μM), as shown in Figure 1. This
difference was statistically significant (Mann-Whitney U=2.0, p=0.005). As shown in Figure 2,
[DA]p also varied significantly between virgin and postpartum groups as well (Mann-Whitney
U=2.0, p=0.008), with postpartum dams releasing more DA per stimulation pulse (Mdn=62 nM)
than virgin animals (Mdn=8.833 nM). Also illustrated in Figure 2, although Vmax did not vary
significantly between parity groups, there was a trend toward significance (Mann-Whitney
U=7.0, p=0.051), with postpartum animals showing reduced velocity of DA transport
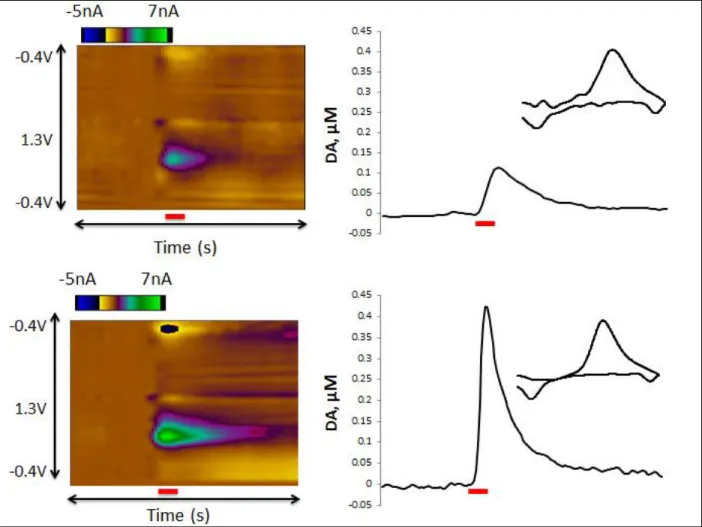
(Mdn=1600 nM/sec) compared to virgins (Mdn=826.667 nM/sec). Figure 3 shows color plots
from representative animals to illustrate these differences.
Change from Baseline
To examine the effect of nasal administration of OT or its vehicle on DA release, the
percent change from baseline was determined for each 15 minute averaged epoch. Regardless of
nasal treatment group or parity status, there was no change from baseline, as shown in Figure 4.
time [F(3, 30) = 0.587, p=0.628] on percent change from baseline, nor was there an interaction of
group and time [F(9, 30) = 0.361, p=0.945].
Discussion
Nasal Oxytocin Does Not Modulate Stimulated Phasic Dopamine Release
Although the interactive role of OT and DA in behavior is under thorough investigation,
the direct pharmacological effect of OT administration on the DA system is relatively unclear. In
the present study, the effect of nasal OT administration on stimulated DA release in the NAc was
examined. Although we predicted that OT would increase DA release, no effect was found. This
runs in contrast to the behavioral studies upon which this project was designed, which found that
OT exerted either a facilitatory or inhibitory effect on the DA system, depending on the
behavioral context under investigation.
We anticipated a simple, direct effect of nasal OT on DA release, but it appears that the
process may be more complex. Indeed, even the basic assumption that OT directly facilitates
social behavior has been questioned in the literature. Though OT is often defined as the “moral
molecule” or the “hug hormone,” its effects on behavior may be less than straightforward. Many
behavioral studies tend to consider only a top-down effect of OT on social behavior; i.e., changes
in behavior are due to OT’s direct effect on the central nervous system. However, OT’s effect on
peripheral organs is not to be dismissed, and bottom-up processes should be taken into
consideration. Beyond the central nervous system, OT receptors are present throughout the body,
including the heart and cardiovascular system, the kidneys, mammary tissues, and the
reproductive tract of both males and females, all of which could be affected by systemic OT
administration (Gimpl et al., 2001). Indeed, OT administration has been known to reduce body
Additionally, organs in the periphery may influence OT release in the nervous system, as both
the heart (van Lieshout, Wieling, Karemaker, & Eckberg, 1991) and the kidneys (Ciriello, 1998)
have afferent projections to the hypothalamus. Vaginocervical stimulation has been found to
increase OT release in the olfactory bulb (OB) and enhance social recognition memory
(Larrazolo-López et al., 2008), so it is entirely possible that systemic OT’s effect on organs can
indirectly influence central nervous system activity and behavior by extension.
In addition to OT’s peripheral effects, OT has been found to exert an anxiolytic effect
(Yoshida et al., 2009). Like its effects on social behavior, this reduction in anxiety may be due to
the complex interaction of its central and peripheral effects. It is also important to consider
whether OT’s social behavior effects may be partly due to its anxiolytic nature (for review, see
Churchland & Winkielman, 2012). As Churchland and Winkielman note, the basis of social
behavior is one’s willingness to approach another, which is sensitive to anxiety levels. In this
way, an increase in social behavior may be just one result of a general reduction in anxiety.
All of the above illustrate the complex nature of OT’s effects on social behavior. Using
nasal OT as our route of administration does little to simplify the problem, as nasal OT’s route
into the nervous system has been disputed. The potential benefits of nasal administration of
peptides were discussed earlier, but the exact route by which nasally administered neuropeptides
enter the brain has been disputed (for review, see Leng & Ludwig, 2015; Quintana, Alvares,
Hickie, & Guastella, 2015). The most likely route of access is the olfactory pathway, in which
molecules reach the OB via olfactory fibers, and then are transferred to brain tissue or
cerebrospinal fluid (Quintana et al., 2015). Through this pathway, it is theoretically possible that
nasal OT could reach more centralized brain regions, as the OB has projections to the amygdala
under what specific circumstances. A neuroimaging study found that nasal OT only activated the
primary olfactory system, some brain stem regions, and sensory afferents, suggesting that OT
was not able to penetrate into more centrally located brain regions (Ferris et al., 2015). In
contrast, a microdialysis study found that nasal OT increased OT levels in the amygdala and
hippocampus, suggesting that it is indeed reaching central brain regions (Neumann et al., 2013).
However, the implantation of microdialysis probes causes significant tissue damage, and plasma
OT may have been unintentionally measured as blood vessels were ruptured.
This raises the question of how nasal OT has been found to alter social behavior, if it may
not reach past the OB, brain stem, and sensory afferents. Again, it is important to note the
interplay of top-down and bottom-up processes. Even if nasal OT is unable to penetrate into
deeper brain regions, its superficial effects may be sufficient to elicit social behavior changes. Its
effects on the OB may alter how olfactory stimuli is processed, leading to altered social
behavior. Indeed, infusion of OT into the OB is sufficient to enhance social recognition (Dluzen,
Shang, & Landgraf, 2001).
Beyond the complexity of the OT system, a few factors of our study may have
contributed to the lack of effect. A significant constraint is our use of normal virgin and
postpartum animals; it is possible that we encountered a ceiling effect, whereby administering
OT to a normal OT system had little impact. One next step will be to look at animals with
disrupted OT systems. For example, postpartum dams treated with cocaine during gestation show
a number of OT disruptions, including lower levels of OT in various areas of the brain, including
the VTA (Johns, Lubin, Walker, Meter, & Mason, 1997), and reductions in OT receptor binding
(Jarrett, McMurray, Walker, & Johns, 2006). These changes may be partly responsible for the
Noonan, & Zimmerman, 1994). If we administer the same experimental procedure to
cocaine-exposed dams and see no effect, it will provide further evidence that nasal OT does not directly
affect the mesolimbic DA system. However, if an effect is found, it would suggest that a ceiling
effect was responsible for the results from the present study.
In addition to the possible ceiling effect of OT, the present study contained some
methodological limitations. First, we measured evoked DA release by electrically stimulating the
VTA. Although this provides the benefit of controlling the environment under which each
measurement is taken, it is in no way a realistic representation of normal brain activity, and the
relatively large current applied to the VTA may mask more subtle changes that could be
occurring. Secondly, we used one acute administration of nasal OT, which may not be sufficient
to elicit noticeable changes, and follow-up experiments may utilize repeated treatments before
the experiment.
Pregnancy Affects Stimulated Dopamine Release at Baseline
When comparing baseline measurements between animal groups, we found significantly
higher DA concentrations in early postpartum dams compared to virgin animals. This is a
plausible finding in consideration with other studies examining the role of the DA system in the
postpartum period. In order to prepare for the onslaught of novel pup-stimuli and the
energy-intensive task of caring for a litter, the peripartum period is rife with changes in the DA system.
However, the precise changes that occur in this time are still under investigation. Postpartum
animals have been found to exhibit reductions in 3,4-Dihydroxyphenylacetic acid (DOPAC), a
metabolite of DA, indicating alterations in DA turnover (Byrnes, Bridges, Scanlan, Babb, &
Byrnes, 2007; Olazábal, Abercrombie, Rosenblatt, & Morrell, 2004). A similar effect is found
Similarly, we found a trend toward a reduction in the velocity of DA transporters in postpartum
dams. Together, these findings suggest that DA is not cleared as readily from the synapse in
postpartum dams as it is in virgin animals. Additionally, we found that more DA was released
per pulse of electrical stimulation in postpartum dams. It is unclear why this may be, as little
research has been performed examining basal DA content in the NAc in the early postpartum
period. Tissue studies have found similar levels of DA content between virgin animals and
postpartum dams (Byrnes et al., 2007), but in vivo experiments have revealed lower basal DA
levels in postpartum dams (Afonso et al., 2009; Afonso, King, Novakov, Burton, & Fleming,
2011; Afonso et al., 2013). Our findings provide additional information as to how the
mesolimbic DA system changes in the early postpartum period, and follow-up experiments can
investigate the exact mechanisms at play.
Conclusion
Although we anticipated a direct excitatory effect of nasal OT on NAc DA release, this
was not the case, suggesting a more complex relationship between the two systems. However,
we did find an effect of postpartum status on basal DA release, contributing to the understanding
of changes in the mesolimbic DA system in the early postpartum period. Further experiments
using cocaine-treated dams will reveal whether nasal OT exerts a different effect on animals with
References
Afonso, V. M., King, S., Chatterjee, D., & Fleming, A. S. (2009). Hormones that increase maternal responsiveness affect accumbal dopaminergic responses to pup- and food-stimuli in the female rat. Hormones and Behavior, 56(1), 11–23.
http://doi.org/10.1016/j.yhbeh.2009.02.003
Afonso, V. M., King, S. J., Novakov, M., Burton, C. L., & Fleming, A. S. (2011). Accumbal dopamine function in postpartum rats that were raised without their mothers. Hormones and Behavior, 60(5), 632–643. http://doi.org/10.1016/j.yhbeh.2011.08.016
Afonso, V. M., Shams, W. M., Jin, D., & Fleming, A. S. (2013). Distal pup cues evoke dopamine responses in hormonally primed rats in the absence of pup experience or ongoing maternal behavior. The Journal of Neuroscience : The Official Journal of the Society for
Neuroscience, 33(6), 2305–12. http://doi.org/10.1523/JNEUROSCI.2081-12.2013
Baracz, S. J., & Cornish, J. L. (2013). Oxytocin modulates dopamine-mediated reward in the rat subthalamic nucleus. Hormones and Behavior, 63(2), 370–375.
http://doi.org/10.1016/j.yhbeh.2012.12.003
Baskerville, T. A., Allard, J., Wayman, C., & Douglas, A. J. (2009). Dopamine-oxytocin interactions in penile erection. European Journal of Neuroscience, 30(11), 2151–2164. http://doi.org/10.1111/j.1460-9568.2009.06999.x
Brom, M., Both, S., Laan, E., Everaerd, W., & Spinhoven, P. (2014). The role of conditioning, learning and dopamine in sexual behavior: A narrative review of animal and human studies.
Neuroscience & Biobehavioral Reviews, 38, 38–59. http://doi.org/10.1016/j.neubiorev.2013.10.014
Byrnes, E. M., Bridges, R. S., Scanlan, V. F., Babb, J. a, & Byrnes, J. J. (2007). Sensorimotor gating and dopamine function in postpartum rats. Neuropsychopharmacology, 32(5), 1021– 31. http://doi.org/10.1038/sj.npp.1301222
Carson, D. S., Hunt, G. E., Guastella, A. J., Barber, L., Cornish, J. L., Arnold, J. C., …
McGregor, I. S. (2010). Systemically administered oxytocin decreases methamphetamine activation of the subthalamic nucleus and accumbens core and stimulates oxytocinergic neurons in the hypothalamus. Addiction Biology, 15(4), 448–463.
http://doi.org/10.1111/j.1369-1600.2010.00247.x
Champagne, F. a, Chretien, P., Stevenson, C. W., Zhang, T. Y., Gratton, A., & Meaney, M. J. (2004). Variations in nucleus accumbens dopamine associated with individual differences in maternal behavior in the rat. The Journal of Neuroscience, 24(17), 4113–23.
Churchland, P. S., & Winkielman, P. (2012). Modulating social behavior with oxytocin: How does it work? What does it mean? Hormones and Behavior, 61(3), 392–399.
http://doi.org/10.1016/j.yhbeh.2011.12.003
Ciriello, J. (1998). Afferent renal inputs to paraventricular nucleus vasopressin and oxytocin neurosecretory neurons. The American Journal of Physiology, 275(6 Pt 2), R1745–54. Retrieved from http://www.ncbi.nlm.nih.gov/pubmed/9843863
Da Silva, M. P., Cedraz-Mercez, P. L., & Varanda, W. a. (2014). Effects of nitric oxide on magnocellular neurons of the supraoptic nucleus involve multiple mechanisms. Brazilian Journal of Medical and Biological Research = Revista Brasileira de Pesquisas Médicas E Biológicas / Sociedade Brasileira de Biofísica ... [et Al.], 47(2), 90–100.
http://doi.org/10.1590/1414-431X20133326
Day, J. J., Roitman, M. F., Wightman, R. M., & Carelli, R. M. (2007). Associative learning mediates dynamic shifts in dopamine signaling in the nucleus accumbens. Nature Neuroscience, 10(8), 1020–8. http://doi.org/10.1038/nn1923
Di Chiara, G., Bassareo, V., Fenu, S., De Luca, M. A., Spina, L., Cadoni, C., … Lecca, D. (2004). Dopamine and drug addiction: The nucleus accumbens shell connection.
Neuropharmacology, 47(SUPPL. 1), 227–241. http://doi.org/10.1016/j.neuropharm.2004.06.032
Dluzen, D., Shang, Y., & Landgraf, R. (2001). Oxytocin, norepinephrine and olfactory bulb mediated recognition. In A. Marchlewska-Koj, J. Lepri, & D. Müller-Schwarze (Eds.),
Chemical Signals in Vertebrates (9th ed., pp. 107–115). New York: Springer US.
Domes, G., Heinrichs, M., Kumbier, E., Grossmann, A., Hauenstein, K., & Herpertz, S. C. (2013). Effects of intranasal oxytocin on the neural basis of face processing in autism spectrum disorder. Biological Psychiatry, 74(3), 164–171.
http://doi.org/10.1016/j.biopsych.2013.02.007
Fahrbach, S. (1984). Oxytocin induction of short-latency maternal behavior in nulliparous, estrogen-primed female rats. Hormones and Behavior, 18(3), 267–286.
http://doi.org/10.1016/0018-506X(84)90016-3
Feifel, D., MacDonald, K., Nguyen, A., Cobb, P., Warlan, H., Galangue, B., … Hadley, A. (2010). Adjunctive intranasal oxytocin reduces symptoms in schizophrenia patients.
Biological Psychiatry, 68(7), 678–680. http://doi.org/10.1016/j.biopsych.2010.04.039
Gimpl, G., Fahrenholz, F., & Gene, C. (2001). The oxytocin receptor system: Structure, function, and regulation. Physiological Reviews, 81(2), 629–683.
Gregory, R., Cheng, H., Rupp, H. a., Sengelaub, D. R., & Heiman, J. R. (2015). Oxytocin
increases VTA activation to infant and sexual stimuli in nulliparous and postpartum women.
Groppe, S. E., Gossen, A., Rademacher, L., Hahn, A., Westphal, L., Gründer, G., &
Spreckelmeyer, K. N. (2013). Oxytocin influences processing of socially relevant cues in the ventral tegmental area of the human brain. Biological Psychiatry, 74, 172–179. http://doi.org/10.1016/j.biopsych.2012.12.023
Guastella, A. J., Einfeld, S. L., Gray, K. M., Rinehart, N. J., Tonge, B. J., Lambert, T. J., & Hickie, I. B. (2010). Intranasal oxytocin improves emotion recognition for youth with autism spectrum disorders. Biological Psychiatry, 67(7), 692–694.
http://doi.org/10.1016/j.biopsych.2009.09.020
Hicks, C., Ramos, L., Reekie, T., Misagh, G. H., Narlawar, R., Kassiou, M., & McGregor, I. S. (2014). Body temperature and cardiac changes induced by peripherally administered oxytocin, vasopressin and the non-peptide oxytocin receptor agonist WAY 267,464: A biotelemetry study in rats. British Journal of Pharmacology, 171(11), 2868–2887. http://doi.org/10.1111/bph.12613
Insel, T. R. (2003). Is social attachment an addictive disorder? Physiology & Behavior, 79(3), 351–357. http://doi.org/10.1016/S0031-9384(03)00148-3
Jarrett, T., McMurray, M., Walker, C., & Johns, J. (2006). Cocaine treatment alters oxytocin receptor binding but not mRNA production in postpartum rat dams. Neuropeptides, 40(3), 161–167. http://doi.org/10.1016/j.biotechadv.2011.08.021.Secreted
Johns, J., Lubin, D., Walker, C., Meter, K., & Mason, G. (1997). Chronic gestational cocaine treatment decreases oxytocin levels in the medial preoptic area, ventral tegmental area and hippocampus in Sprague-Dawley rats. Neuropeptides, 31(5), 439–43. Retrieved from http://www.pubmedcentral.nih.gov/articlerender.fcgi?artid=3101369&tool=pmcentrez&ren dertype=abstract
Johns, J., Noonan, L., & Zimmerman, L. (1994). Effects of chronic and acute cocaine treatment on the onset of maternal behavior and aggression in Sprague-Dawley rats. Behavioral Neuroscience, 108(1), 107–12. Retrieved from
http://www.ncbi.nlm.nih.gov/pubmed/8192835
Keer, S. ., & Stern, J. . (1999). Dopamine receptor blockade in the nucleus accumbens inhibits maternal retrieval and licking, but enhances nursing behavior in lactating rats. Physiology & Behavior, 67(5), 659–669. http://doi.org/10.1016/S0031-9384(99)00116-X
Kovàcs, G. L., Sarnyai, Z., Barbarczi, E., Szabó, G., & Telegdy, G. (1990). The role of oxytocin-dopamine interactions in cocaine-induced locomotor hyperactivity. Neuropharmacology,
29(4), 365–8. http://doi.org/10.1016/0028-3908(90)90095-9
Lee, H. J., Macbeth, A. H., Pagani, J. H., & Scott Young, W. (2009). Oxytocin: The great facilitator of life. Progress in Neurobiology, 88(2), 127–151.
http://doi.org/10.1016/j.pneurobio.2009.04.001
Leng, G., & Ludwig, M. (2015). Intranasal Oxytocin: Myths and Delusions. Biological Psychiatry, (14), 1–8. http://doi.org/10.1016/j.biopsych.2015.05.003
Lonstein, J. S., Maguire, J., Meinlschmidt, G., & Neumann, I. D. (2014). Emotion and mood adaptations in the peripartum female: Complementary contributions of GABA and oxytocin. Journal of Neuroendocrinology, 26(10), 649–664.
http://doi.org/10.1111/jne.12188
Love, T. M. (2014). Oxytocin, motivation and the role of dopamine. Pharmacology, Biochemistry, and Behavior, 49–60.
http://doi.org/10.1016/j.biotechadv.2011.08.021.Secreted
Marazziti, D., Baroni, S., Giannaccini, G., Betti, L., Massimetti, G., Carmassi, C., & Catena-Dell’Osso, M. (2012). A link between oxytocin and serotonin in humans: Supporting evidence from peripheral markers. European Neuropsychopharmacology, 22(8), 578–583. http://doi.org/10.1016/j.euroneuro.2011.12.010
Mayer, a D., & Rosenblatt, J. S. (1998). A method for regulating the duration of pregnancy and the time of parturition in Sprague-Dawley rats (Charles River CD strain). Developmental Psychobiology, 32(2), 131–6. Retrieved from
http://www.ncbi.nlm.nih.gov/pubmed/9526688
McGregor, I. S., & Bowen, M. T. (2012). Breaking the loop: Oxytocin as a potential treatment for drug addiction. Hormones and Behavior, 61(3), 331–339.
http://doi.org/10.1016/j.yhbeh.2011.12.001
Melis, M. R., Melis, T., Cocco, C., Succu, S., Sanna, F., Pillolla, G., … Argiolas, A. (2007). Oxytocin injected into the ventral tegmental area induces penile erection and increases extracellular dopamine in the nucleus accumbens and paraventricular nucleus of the hypothalamus of male rats. European Journal of Neuroscience, 26, 1026–1035. http://doi.org/10.1111/j.1460-9568.2007.05721.x
Mens, W. B. J., Witter, a., & Van Wimersma Greidanus, T. B. (1983). Penetration of
neurohypophyseal hormones from plasma into cerebrospinal fluid (CSF): Half-times of disappearance of these neuropeptides from CSF. Brain Research, 262(1), 143–149. http://doi.org/10.1016/0006-8993(83)90478-X
Neumann, I. D., Maloumby, R., Beiderbeck, D. I., Lukas, M., & Landgraf, R. (2013). Increased brain and plasma oxytocin after nasal and peripheral administration in rats and mice.
Psychoneuroendocrinology, 38, 1985–1993. http://doi.org/10.1016/j.psyneuen.2013.03.003
Numan, M., & Stolzenberg, D. S. (2009). Medial preoptic area interactions with dopamine neural systems in the control of the onset and maintenance of maternal behavior in rats. Frontiers in Neuroendocrinology, 30(1), 46–64. http://doi.org/10.1016/j.yfrne.2008.10.002
Olazábal, D. E., Abercrombie, E., Rosenblatt, J. S., & Morrell, J. I. (2004). The content of dopamine, serotonin, and their metabolites in the neural circuit that mediates maternal behavior in juvenile and adult rats. Brain Research Bulletin, 63(4), 259–268.
http://doi.org/10.1016/j.brainresbull.2004.02.009
Parker, K. J., Buckmaster, C. L., Schatzberg, A. F., & Lyons, D. M. (2005). Intranasal oxytocin administration attenuates the ACTH stress response in monkeys.
Psychoneuroendocrinology, 30(9), 924–929. http://doi.org/10.1016/j.psyneuen.2005.04.002
Quintana, D. S., Alvares, G. A., Hickie, I. B., & Guastella, A. J. (2015). Do delivery routes of intranasally administered oxytocin account for observed effects on social cognition and behavior? A two-level model. Neuroscience and Biobehavioral Reviews, 49, 182–192. http://doi.org/10.1016/j.neubiorev.2014.12.011
Riem, M. M. E., van IJzendoorn, M. H., Tops, M., Boksem, M. a S., Rombouts, S. a R. B., & Bakermans-Kranenburg, M. J. (2012). No laughing matter: Intranasal oxytocin
administration changes functional brain connectivity during exposure to infant laughter.
Neuropsychopharmacology. Nature Publishing Group. http://doi.org/10.1038/npp.2012.27
Robinson, D., & Howard, E. (2009). Disparity between tonic and phasic ethanol induced dopamine increases in the nucleus accumbens of rats. Alcoholism: Clinical and Experimental Research, 33(7), 1187–1196. http://doi.org/10.1111/j.1530-0277.2009.00942.x.Disparity
Robinson, D., Venton, B., Heien, M., & Wightman, R. (2003). Detecting subsecond dopamine release with fast-scan cyclic voltammetry in vivo. Clinical Chemistry, 1773, 1763–1773. Retrieved from http://www.clinchem.org/content/49/10/1763.short
Robinson, D., & Wightman, R. . (2007). Rapid dopamine release in freely moving rats. In A. Michael & L. Borland (Eds.), Electrochemical Methods for Neuroscience (pp. 17–34). Boca Raton: CRC Press. Retrieved from
http://www.ncbi.nlm.nih.gov/books/NBK2575/?report=printable
Rutherford, H. J. V, Williams, S. K., Moy, S., Mayes, L. C., & Johns, J. M. (2011). Disruption of maternal parenting circuitry by addictive process: Rewiring of reward and stress systems.
Salamone, J. D., Correa, M., Farrar, A., & Mingote, S. M. (2007). Effort-related functions of nucleus accumbens dopamine and associated forebrain circuits. Psychopharmacology,
191(3), 461–82. http://doi.org/10.1007/s00213-006-0668-9
Shahrokh, D. K., Zhang, T. Y., Diorio, J., Gratton, A., & Meaney, M. J. (2010). Oxytocin-dopamine interactions mediate variations in maternal behavior in the rat. Endocrinology,
151(May 2010), 2276–2286. http://doi.org/10.1210/en.2009-1271
Shnitko, T. A., & Robinson, D. L. (2014). Anatomical and pharmacological characterization of catecholamine transients in the medial prefrontal cortex evoked by ventral tegmental area stimulation. Synapse, 68(4), 131–143. http://doi.org/10.1002/syn.21723
Smith, A. S., Ågmo, A., Birnie, A. K., & French, J. a. (2010). Manipulation of the oxytocin system alters social behavior and attraction in pair-bonding primates, Callithrix penicillata.
Hormones and Behavior, 57(2), 255–262. http://doi.org/10.1016/j.yhbeh.2009.12.004
Triana-Del Rio, R., Tecamachaltzi-Silvarán, M. B., Díaz-Estrada, V. X., Herrera-Covarrubias, D., Corona-Morales, A. a., Pfaus, J. G., & Coria-Avila, G. a. (2015). Conditioned same-sex partner preference in male rats is facilitated by oxytocin and dopamine: Effect on sexually dimorphic brain nuclei. Behavioural Brain Research, 283, 69–77.
http://doi.org/10.1016/j.bbr.2015.01.019
Vaccari, C., Lolait, S. J., & Ostrowski, N. L. (1998). Comparative distribution of vasopressin V1b and oxytocin receptor messenger ribonucleic acids in brain. Endocrinology, 139(12), 5015–5033.
Van Lieshout, J. J., Wieling, W., Karemaker, J. M., & Eckberg, D. L. (1991). The vasovagal response. Clinical Science, 81, 575–586.
Yoshida, M., Takayanagi, Y., Inoue, K., Kimura, T., Young, L. J., Onaka, T., & Nishimori, K. (2009). Evidence that oxytocin exerts anxiolytic effects via oxytocin receptor expressed in serotonergic neurons in mice. Journal of Neuroscience, 29(7), 2259–2271.
Figures
Figure 1. Evoked DA at baseline between virgin and postpartum animals. Postpartum animals
released significantly higher concentrations of DA at baseline to the same electrical stimulation
compared to virgin animals (p=0.005). The second and third quartiles are represented by dark
and light gray, respectively. 0
0.2 0.4 0.6 0.8 1 1.2 1.4 1.6
Virgin (n=6) Postpartum (n=7)
[D
A
]max
, μ
M
Figure 2. DA released per pulse ([DA]p) and velocity of DA transporters (Vmax) derived from
electrically evoked DA signals in virgin and postpartum animals. VTA-NAc neurons in
postpartum animals released significantly more DA per pulse of electrical stimulation than virgin
animals (p=0.008). The velocity of DA transporters (Vmax) in postpartum animals was reduced
0 20 40 60 80 100 120
Virgin (n=6) Postpartum (n=7)
[D
A]
p, nM
0 500 1000 1500 2000 2500 3000
Virgin (n=6) Postpartum (n=7)
V
max
, nM/
se
c
compared to virgin animals. This difference, although not statistically significant, was a strong
Figure 3. Representative color plots (left) and current vs. time traces (right) from a virgin female
(top) and postpartum dam (bottom). Color plots display changes in current (color) at a range of
applied potentials (y-axis) over time (x-axis). Inset, DA cyclic voltammogram from respective
Figure 4. Evoked DA concentration as a percent of basal measurements in virgin and postpartum
rats after nasal treatment of either OT or vehicle. [DA]max was averaged into four 15-minute
epochs. No differences were observed between groups or over time. 50%
60% 70% 80% 90% 100% 110% 120%
0-15 15-30 30-45 45-60
Perc
en
t
C
h
an
ge
from
B
as
el
in
e
Time After Treatment (min)
OXT/Virgin (n=4)
OXT/Postpartum (n=4)
VEH/Virgin (n=4)
![Figure 2. DA released per pulse ([DA] p ) and velocity of DA transporters (V max ) derived from electrically evoked DA signals in virgin and postpartum animals](https://thumb-us.123doks.com/thumbv2/123dok_us/8334112.2211752/28.918.108.664.104.867/figure-released-velocity-transporters-derived-electrically-signals-postpartum.webp)