THERMOREGULATORY, METABOLIC, AND CARDIOVASCULAR RESPONSES OF PREGNANT IDMEN TO IMMERSION AND IMMERSION EXERCISE
I•
By /
Daniel Gunther Graetzer
A Thesis submitted to the faculty of The University of North Carolina at Chapel Hill in partial fulfillment of the requirements for the degree of Master
of Arts in the Department of Physical Education.
Chapel Hill
1986
himsel£ against the water and we can gain more support in water which contains more solid matter? For
sea-water contains more solid matter than river-water. For it is denser and more capable 0£ o££ering
resistance.
Aristotle (384-322 B.C.
>,
Problems XXIII:13Why do extremities become most chilled? Is it because they are so narrow? The channels in them, because they are narrow, have but little blood, so that they contain but little heat, £or blood is hot.
DANIEL GUNTHER GRAETZER. Thermoregulatory, Metabolic, and Cardiovascular Responses of Pregnant Women to Immersion and Immersion Exercise. (Under the Direction of DR. ROBERT G. MCMURRAY>
Twelve healthy women underwent 20 minute periods of
immersion rest (THao = 30° C) and immersion exercise (60¾ predicted VOa max) during weeks 15, 25, and 35 of
gestation and 8 weeks postpartum. Maternal core temp, heat storage, immersion tissue insulation, and exercise
tissue conductance were maintained across trials. temps were 1.1° Clower during pregnancy to prevent heat losses during immersion and heat gains during
exercise. Oxygen uptake, heart rate, and systolic
Skin
blood pressure did not change with gestational timing during nonweight-bearing immersion exercise. The
hydrostatic pressure of the water enabled exercise
heart rates to be 17 bts/min lower than the predicted
max land trials at the same energy expenditure. The
heat dissipating and weight-supporting capacities of
the water environment provided safe thermal, metabolic,
and cardiovascular responses in pregnant women during
The author wishes to express his appreciation to
his advisor, Dr. Robert G. McMurray, £or continued
guidance and encouragement during this project.
Sincere thanks is extended to Dr. Vern L. Katz and Dr.
Edgar W. Shields as committee members. Special thanks
is given to Dr. Charles J. Hardy and £ellow graduate students Steven C. Fox and David L. Wright £or their
technical assistance.
Gratitude is also acknowledged to the twelve
unborn babies and their mothers who participated as
subjects in this investigation. Finally, I wish to
thank my parents, Hans and Miriam Graetzer, £or their
support and to the Lord in heaven £or making all this
TABLE OF CONTENTS
LIST OF TABLES
LIST OF FIGURES
CHAPTER
I. THE PROBLEM ..
Introduction
Statement 0£ the Problem
Null Hypotheses ..
De£inition 0£ Terms.
Abbreviations
Delimitations
Limitations.
Signi£icance 0£ the Problem.
II. REVIEW OF LITERATURE III. METHODOLOGY.
Subjects
Predicted Max Land Trials.
Immersion Trials
Instrumentation ..
Statistical Analysis
IV. RESULTS • .
iv
PAGE vi vii
1 1 2
2 4
7 8
9
9 10
21 21 22
23 25
Conclusions.
Directions £or Future Research BIBLIOGRAPHY
APPENDIX
A. HUMAN SUBJECTS CONSENT FORM.
B. INDIVIDUAL DATA . . . .
50
51
53
63
66 C. DESCRIPTIVE STATISTICS/ ANOVA TABLES .. 95
D. SMITH GRADUATE RESEARCH FUND APPLICATION 103
E. AUTHOR'S VITAE ... 106
LIST OF TABLES
TABLE PAGE
1. Anthropometric Characteristics of Subjects . 21
2. Mean (~SD> Thermal Responses during 20 Minute Periods of the 1 Hour Trial as Related to
Pregnancy Status • 96
3. Mean (~SD> Cardiovascular and Metabolic Responses during 20 Minute Periods of the 1 Hour Trial as Related to Pregnancy Status. . . • 97
FIGURE
1. Core Temperature ..
2. Mean Skin Temperature
3. Mean Body Temperature
4. Heart Rate
. . . . .
5. Systolic Blood Pressure6. Heat Storage
.
7. Immersion Heat Insulation
8. Exercise Heat Conductance
9. Basal Energy Expenditure.
10. Immersion Energy Expenditure
11. Exercise Energy Expenditure
vii
PAGE
32 33 34
35 36
37
38
39
40 41
Introduction
CHAPTER I THE PROBLEM
Many women are participating in exercise programs during pregnancy due to increasing awareness 0£ the bene£its 0£ physical £itness. The increased metabolic rate during physical exertion elevates body core temperature. Maternal hyperthermia has been
substantiated as a teratogen in both animals (Edwards, 1967; Kilham & Ferm, 1976) and humans (Fisher & Smith, 1981; Miller, Smith, & Shepard, 1978; Smith, 1982; Smith, Clarren, & Harvey, 1978).
Aerobic exercise in the water is not associated
with as dramatic a rise in core temperature as on land (Nielsen & Davies, 1976). The thermal conductivity 0£
water is 25 times that 0£ air enabling cardiovascular training at a high metabolic rate without the
associated hyperthermia. The physiological responses to swim conditioning have been extensively explored
<McArdle, Glaser, & Magel, 1971; McMurray & Horvath, 1979). More recently, the exercise responses
evaluation of the thermal and metabolic responses of
pregnant women to aquatic exercise has not been
directly assessed. Therefore, this investigation will
examine the thermoregulatory, metabolic, and
cardiovascular responses of pregnant women throughout
advancing gestation to immersion exercise.
Statement of the Problem
The purpose of this study is to investigate the
thermoregulatory, metabolic, and cardiovascular
consequences of head-out immersion and
moderate-intensity exercise in water on healthy,
pregnant women across weeks 15, 25, and 35 of gestation and 8 weeks post parturition.
Null Hypotheses
HO: There will be no statistically significant
difference (p<.05) in the core temperature, mean skin
temperature, mean body temperature, heart rate, and
systolic blood pressure of 12 healthy women as a
£unction 0£ pregnancy status (15, 25, and 35 weeks 0£ gestation and 8 weeks postpartum).
HO: There will be no statistically significant
difference (p<.05) in the core temperature, mean skin
temperature, mean body temperature, heart rate, and
function of activity (immersion rest, immersion
exercise, and recovery).
3
HO: There will be no statistically significant
difference (p<.05) in the heat storage of 12 healthy
women as a function of pregnancy status (15, 25, and 35
weeks of gestation and 8 weeks postpartum).
HO: There will be no statistically significant
difference Cp<.05) in the heat storage of 12 healthy
women as a function of time (minute 5 of immersion to minute 20 of immersion, minute 20 of immersion to
minute 20 of exercise, and minute 20 of exercise to
minute 20 of recovery>.
HO: There will be no statistically significant
difference (p<.05) in the tissue insulation during
immersion rest of 12 healthy women as a function of
pregnancy status (15, 25, and 35 weeks of gestation and 8 weeks postpartum).
HO: There will be no statistically significant
difference (p<.05) in the tissue conductance during
immersion exercise of 12 healthy women as a function of
HO: There will be no statistically significant
difference Cp<.05) in the tissue conductance during
immersion exercise of 12 healthy women as a function of
time (minutes 5, 10, 15, and 20 of immersion exercise>.
HO: There will be no statistically significant
difference Cp<.05) in the caloric energy expenditure
during 20 minute periods of land rest, immersion rest,
and immersion exercise of 12 healthy women as a
function of pregnancy status (15, 25, and 35 weeks of
gestation and 8 weeks postpartum>.
Definition of Terms
(1) Core Temperature - The rectal temperature in
degrees Celsius as measured approximately 15 ems into
the rectum.
(2) Mean Skin Temperature - The average skin
temperature of the body in degrees Celsius. With 27¾
of the body surface area above water and 73¾ immersed
(Hafen & Karren, 1984, p. 263), mean skin temperature
is determined by the formula:
Tsk = (0.27 x Tsh) + (0.73 x Tbk)
Tsk = mean skin temperature
Tsh = shoulder temperature
5
(3) Mean Body Temperature - The average
temperature of the entire body in degrees Celsius using
rectal and skin measurements in the formula advanced by
McMurray & Horvath (1979):
TB= (0.35 x Tsk) + (0.65 x Tr>
TB = total body temperature
Tsk = mean skin temperature
Tr = rectal temperature.
(4) Heat Storage - The rate of body heat storage
in response to a heat stress directly proportional to
body weight and change in mean body temperature in the
formula advanced from Dressendorfer, Smith, Baker, &
Hong (1977):
S = 0.83 x Kg x ~TB x 3
S = body heat storage in Kcals/hour
0.83 = average specific heat of body tissues
Kg =bodyweight in kilograms
~TB = change in mean body temperature in degrees
Celsius over the 20 minute period using
minute 5 of immersion as the baseline for
immersion, minute 20 of immersion as the
baseline for exercise, and minute 20 of
exercise as the baseline for recovery
3 = conversion factor to change mean body
(5) Tissue insulation - The insulation of body
tissues determined assuming peripheral temperature is
represented by mean skin temperature with a peripheral mass equal to 0.4 body weight. Core temperature is assumed to represent the remaining 0.6 body weight and 8¾ of the heat loss is assumed to occur through the
respiratory tract. With the denominator representing the rate of skin heat loss, tissue insulation is
determined by the formula from Rennie, Covino, Howell,
Song, Kang, & Hong (1962):
I=
<Tr - Tsk)/M - (0.08 x M) + SI = tissue insulation in °C x hour/Kcal
Tr = rectal temperature
Tsk = mean skin temperature
M = metabolic heat production in Keala/hour
S = heat storage in Kcals/hour.
(6) Tissue conductance - The rate of heat loss
through the skin dependent upon peripheral blood flow
and the skin-to-ambient thermal gradient. Metabolic
heat production is assumed to be the same as metabolic
energy production as the external work for swimming is
negligible, 1-5¾, (Holmer, 1974) and the net efficiency
for underwater bicycling is unknown. This
7
insulation index <Nadel & Horvath, 1975, p. 105) in the formula:
K = 1 / I
K = tissue conductance in °C x hour/Kcal I= tissue insulation in °C x hour/Kcal.
(7) Caloric energy expenditure - The total energy
expenditure in kilocalories by the subject per 20
minute activity (land rest, immersion rest, immersion
exercise) in the formula:
Kcals/20 min= Keala/minx 20.
(8) Heart Rate - The number of ventricular
contractions per minute as determined by the distance
between R waves on an electrocardiogram strip during the last 10 seconds of the minute.
(9) Systolic Blood Pressure - The maximal pressure
in mm Hg of the blood on the arterial walls as
determined by acoustic sphygmomanometer.
Abbreviations
BMR
s
I
K SBP
DBP
Basal metabolic rate
Heat storage
Tissue insulation
Tissue conductance
Systolic blood pressure
KPM
RPM
L/min
Kilopond meters per minute
Revolutions per minute
Liters per minute
Celsius
Kilocalorie
Millimeters of Mercury
Percent oxygen in expired air
Percent carbon dioxide in expired air
Volume of oxygen uptake per minute C
Kcal
mm Hg
FEOa
FECOa
VOa VCOa Temp
DV
Volume of carbon dioxide expired per minute
Temperature
Dependent variable
SD Standard deviation
Delimitations
(1) This experiment was conducted on 12 healthy
Caucasian women free of evident cardiovascular disease or predisposing problems to ex~rcise with no prior
extensive swim conditioning.
(2) Individual immersion exercise workloads were determined by heart rate response to previous land
bicycle trials throughout pregnancy and postpartum.
(3) A repeated measures research design was
9
as the baseline comparison £or physiological responses
during weeks 15, 25, and 35 of gestation.
Limitations
(1) Only one exercise intensity (60¾ heart
rate-predicted VOa max>, duration (20 minutes>, and
water temperature (30° C> was examined.
(2) Swim conditioning was simulated using an
underwater bicycle ergometer which requires more leg work than normal swimming.
(3) Thermal responses during exercise may be
confounded by the previous 20 minute period of
immersion rest.
(4) Findings will only be valid £or steady state exercise and cannot be generalized to maximal e£forts
or transitional stages of exercise.
Signi£icance 0£ the Problem
This study will investigate aquatic exercise throughout advancing human gestation. The heat
dissipating capacity and hydrostatic pressure of the
water may create a sa£e and beneficial exercise
REVIEW OF LITERATURE
Pregnant women are being encouraged to exercise by everyone £ram "£itness counselors" to movie stars
(DeLyser, 1982). Potential bene£its 0£ aerobic
conditioning include improved cardiovascular endurance, muscle tone, coordination, balance, and a sense 0£ £eeling good and looking better. The American College 0£ Obstetricians and Gynecologists <ACOG, 1985)
introduced preliminary guidelines £or unsupervised
exercise during pregnancy. These recommendations have been criticized as being too vague, not based on hard data, and taking input £rem too £ew pro£essionals
(Gauthier, 1986). Pregnant women remain without well-established guidelines £or exercise prescription.
An elevation in the expectant mother's body core temperature in excess 0£ 38.9° Chas been linked to adverse £etal development (Artal & Wiswell, 1986). The enhanced metabolic rate during aerobic exercise
increases body temperature. The magnitude 0£ elevation is dependent on the exercise type, intensity, and duration, and ambient heat and humidity conditions.
11
approximately .5 degrees Celsius higher than maternal temperature <Adamsons, 1966). The majority of fetal heat is transferred to the mother across the placenta,
while the fetal skin, amniotic fluid, and uterine wall dissipate a smaller proportion of heat away from the fetus to maintain thermal balance (Adamsons, 1966; Abrams, Caton, Clapp, & Barron, 1970). Schroder, Gilbert, & Power (1982) suggest that maternal body temperature is the major influence over fetal
temperature, with changes in uterine blood flow and fetal metabolism being secondary determinents.
Animal research has shown moderate to severe maternal hyperthermia during the first one-half of gestation to be teratogenic. Edwards (1967) found that a 1.5° C elevation in the core temperature of pregnant guinea pigs halted the growth of neural cells in the epidural layer of the embryo's brain and a 3° C elevation can kill these mitotic cells. Edwards also noted a temperature elevation of 2.5° C in pregnant guinea pigs for one hour daily at 18 to 25 days gestation resulted in progeny with microencephaly,
on the fetal guinea pig (Edwards, 1971). Using
pregnant hamsters, Kilham & Ferm (1976) found maternal hyperthermia of 40° C to 41° C for one hour on
gestation day 8 resulted in an increased rate of resorption and a high frequency of anencephaly and encephalocele, with longer exposure often causing maternal death. Maternal hyperthermia as a teratogen has also been substantiated in white mice CLecyk, 1966), rats CSkrebs & Frank, 1963), sheep (Hartley, Alexander,
&
Edwards, 1974; Alexander&
Williams, 1971), and chickens <Alsop, 1919).Retrospective studies on human birth defects have also revealed neural tube defects associated with
hyperthermia. The nature and degree of the anomality appears to be related primarily to the timing of the maternal hyperthermia, rather than its cause. Rubella, a viral disease characterized by a high fever, has long been recognized as being teratogenic when acquired
during the first trimester (Swan, 1949). Smith,
Clarren, & Harvey (1978) examined the cases of 8 women where a fever of 38.9° C occurred at 4 to 6 weeks
13
neurogenic arthrogryposis, and central nervous system dysgenesis occurred. The causes of the hyperthermia varied from febrile illness to prolonged sauna bathing. Similar to the animal studies, no apparent problems in morphogenesis following maternal hyperthermia in the later half of gestation have been reported. The following tabulates the defect and degree of
microcephaly of 24 human dysmorphic offspring with a history of early maternal hyperthermia (Smith, 1982).
PROBLEM
CNS Structural
Neuronal heterotopias l'licropthalmia
Microcephaly Functional
Mental deficiency Hypotonicity
<Hypotonic diplegia) Hypertonicity
Neurogenic contractures Seizures
FACIAL
Microphthalmia Micrognathia
Midfacial hypoplasia External ear anomalies Cleft lip and/or palate OTHER
Micropenis <LH deficiency)
GESTATIONAL TIMING OF HYPERTHERMIA 4-7 WKS 2/2 6/11 1/10 8/8 5/10 (2/10) 3/10 2/11 5/8 6/11 5/11 3/11 3/11 3/11 2/7 8-14 WKS 2/2 1/13 1/13 11/11 7/13 (4/13) 3/10 3/13 2/11 1/13 1/13 1/13 1/13 0/13 0/6
¾ OF TOTAL 100¾ 29¾ 9¾ 100¾ 52¾ 26¾ 30¾ 21¾ 33¾ 29¾ 25¾ 17¾ 17¾ 13¾ 15¾
These hyperthermia-associated anomalities are
Pregnancy causes an increase in body weight along
with complex thermal, hormonal, metabolic, and cardiovascular changes which complicate maternal responses to exercise. Basal body temperature rises approximately .5° C following ovulation with increased progesterone concentration. During pregnancy, this . temperature increase persists until about mid-gestation before declining to normal levels <Wilson, Morganti, Zervoudakis, Letcher, Romney, Von Deyon, Papera, Sealey, & Laragh, 1980). Although progesterone
continues to increase toward term <Tulchinski, Hobel, Yaeger, & Marshall, 1972>, the decline in resting body temperature is attributed to an increase in estrogen
<Wilson, et al., 1980). Progesterone and estrodial concentrations increase during exercise (Jurkowsky, Jones, Walker, Younglai, & Sutton, 1978). Resting
oxygen consumption increases with gestation to peak at 16¾ to 32¾ above non-pregnant status <Emerson, Saxena,
& Poindexter, 1972). A deconditioning effect near term is seen with the increased caloric cost of daily
15
by 32 weeks approximately 20 bts/min higher than the nonpregnant state <Pritchard & MacDonald, 1980).
Resting systolic blood pressure decreases slightly
until mid-gestation be£ore rising back to normal levels during the third trimester <Hytten & Leitch, 1964). These physiological changes 0£ advancing gestation create complicated interactions during aerobic exercise.
Total heat production 0£ the body during exercise may increase to as much as 20 times over rest. Only 20¾ to 25¾ 0£ the additional energy expenditure is used £or metabolism, while the remaining 75¾ to 80¾ is
trans£ormed into heat <Rowell, 1974). During land exercise, most 0£ this heat is lost to the environment through evaporation, convection, and radiation. Some is stored, however, resulting in an elevation in body temperature and upsetting thermal equilibrium. The body's £irst line 0£ de£ense to maintain thermal
balance is a circulatory change adjusting blood £low £ram the body's core to the periphery (Buskirk, 1977). The hypothalamus regulates the body's thermostat by specialized neurons leading to thermal receptors in the skin <McArdle, Katch, & Katch, 1986). The exercise
mode and ambient conditions strongly a££ect the rate 0£ heat dissipation, heat storage, and body temperature
The thermal and metabolic effects of land exercise
during animal pregnancy have been directly studied
(Clapp, 1980; Bell, Hales, King, & Fawcett, 1983).
Lotgering, Gilbert, & Longo (1983) observed that in
pregnant ewes, fetal temperature increases lag behind
maternal temperature increases as a consequence of the
heat diffusion capacity of the fetus and amniotic
fluid. Placental heat transfer remained constant.
Fetal metabolic rate showed only a small increase
supporting the hypothesis that moderate exercise does
not present a stressful or hypoxic event to the fetus.
Uterine blood flow decreases with the intensity and duration of exercise CLotgering, et al., 1983) with the magnitude being variable across animal models (Artal &
Wiswell, 1986). Fetal metabolism is maintained, however, due to a hemoconcentration and increased
oxygen extraction CLotgering, et al., 1983). As with all animal studies, discretion must be employed when extrapolating these findings to humans.
Increases in oxygen consumption and heart rate
while walking have a linear relationship with increases in body weight throughout human pregnancy (Knuttgen &
Emerson, 1974). Weight-supported bicycle exercise,
however, showed no significant differences in oxygen
17
Botti, Anderson, & Bennett (1985) found that heat
storage in conditioned women was not greatly increased
as a result of a mild jogging program throughout
-advancing pregnancy. However, for safety reasons
exercise was stopped when their subjects approached a core temperature of 39° C. This well exceeded the ACOG recommendation of a 38° C limit.
Dressendorfer & Goodlin (1980) found fetal heart
rate will increase approximately 1 bt/min per 10 bt/min rise in maternal heart rate during exercise. Dynamic
muscular activity has also been shown to cause fetal bradycardia or tachycardia (Hon & Wohlgemuth, 1961;
Pomerance, Gluck, & Lynch, 1974) possibly due to
reduced uterine blood flow. Uterine blood flow
reductions could compromise dissipation of heat through
the placenta (Schroder, et al., 1982). Clapp (1985),
however, found no correlation between the increases in
fetal heart rate and the elevation in maternal exercise
temperature (range, 0.1° to 1.3° C) during exercise.
Exercise in the water allows cardiovascular training at a high metabolic rate without the
associated shift in body temperature <Nielsen & Davies,
1976). The skin-to-ambient conductivity gradient and
specific heat of water are about 25 and 1000 times
greater, respectively, than that of air (Miller, 1972).
water temperature (Nunneley, 1978>. Metabolic heat is liberated in proportion to the swimming intensity and temperature of the water enabling heat to be gained in warm water and lost in cold water. Convection is
essentially the only means of heat transfer in the water which increases with water turbulence during higher intensity swimming <Nadel, Holmer, Bergh, Astrand, & Stolwijk, 1974). The thickness of the subcutaneous layer of fat is the major determinent of this heat flux (Nadel, et al., 1974). Women tolerate mild cold changes better than men due to insulation by the female's higher average percentage of body fat
<Rennie, Covino, Howell, Song, Kang, & Hong, 1962). Bodyfat percentage does not change significantly with body weight increases during pregnancy (Pritchard &
MacDonald, 1908). Females have a greater core to skin temperature difference enhancing their heat diffusion capacity <Fox, Lofstedt, Woodward, Eriksson, &
Werkstrom, 1969). Women also have the advantage of a lower metabolic rate per unit surface area and a higher heat conductance index <DuBois, Ebaugh, & Hardy, 1952). The water environment and the natural characteristics
of females create a safe thermal climate for exercise during pregnancy.
A high maternal temperature continuing to elevate
metabolism following land exercise (Q,o effect) could be detrimental. Hypothalamic influence on both ~he
19
temperature-regulating and appetite-control centers
(Guyton, 1981) may cause the increased appetite seen by many after swimming. An enhanced caloric intake is often recommended by obstetricians.
Heart rate response is lower during submaximal swimming than while running or cycling on land at an equalized metabolic rate (McArdle, Glaser, & Magel, 1971) due to the hydrostatic pressure of the water and supine body position of the subject enhancing venous return. This enhanced cardiac filling results in a larger stroke volume so heart rate will not have to increase as high to maintain the needed cardiac output. This adjustment is due to a baroreceptor response
causing a higher cardiac output, higher blood pressure, and reflexly lowered heart rate during immersion (Farhi
& Linnarsson, 1977). The slight but consistent rise in fetal heart rate during land exercise may be reduced by the lower maternal heart rate during swimming. Central blood flow may be increased by the higher cardiac
output, restriction of superficial circulation, and decrease in venous pooling seen during immersion.
1978; McMurray, 1983). Exercise in the water may reduce fetal heart rate changes and possible
utero-placental ischemia, providing less stress to the fetus.
An initial exploration of swim conditioning during pregnancy demonstrated that women were able to maintain initial fitness levels over 12 weeks of gestation while pregnant control subjects were not (Sibley, Ruhling, Cameron-Foster, Christensen, & Bolen, 1981). Surveys on scuba diving during pregnancy revealed no
disturbance in fetal well-being (Newhall, 1981; Bolton, 1980). Nonweight-bearing exercise in the water may be more supportive and less jarring to the abdominal
region. With the popularity of swimming, it is surprising that an in-depth evaluation of aquatic
exercise during pregnancy has not been assessed. Swim
conditioning may be the ideal alternative for gaining the benfits of aerobic exercise while minimizing
possible risks during pregnancy.
The American College of Obstetricians and Gynecologists will undoubtedly modify their exercise recommendations with further research. Information regarding the effects of immersion and immersion
Subjects
CHAPTER III
METHODOLOGY
The subjects used £or this study were 12 healthy,
pregnant Caucasian £emales (Table 1). The subjects
were recruited through advertisements in local
newspapers and signs posted in the North Carolina
Memorial Hospital Obstetrics and Gynecology Clinic.
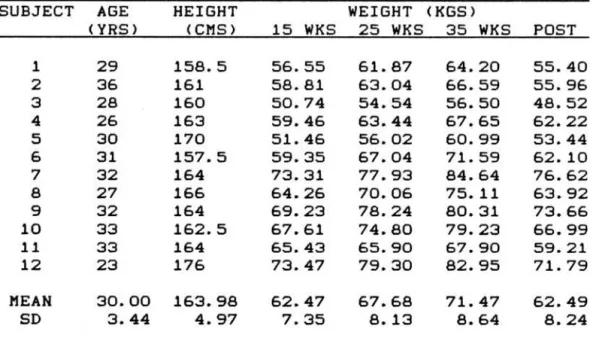
TABLE 1. ANTHROPOMETRIC CHARACTERISTICS OF SUBJECTS
SUBJECT AGE HEIGHT WEIGHT <KGS>
<YRS> (CMS> 15 WKS 25 WKS 35 WKS POST
1 29 158.5 56.55 61.87 64.20 55.40
2 36 161 58.81 63.04 66.59 55.96
3 28 160 50.74 54.54 56.50 48.52
4 26 163 59.46 63.44 67.65 62.22
5 30 170 51.46 56.02 60.99 53.44
6 31 157.5 59.35 67.04 71.59 62.10
7 32 164 73.31 77.93 84.64 76.62
8 27 166 64.26 70.06 75.11 63.92
9 32 164 69.23 78.24 80.31 73.66
10 33 162.5 67.61 74.80 79.23 66.99
11 33 164 65.43 65.90 67.90 59.21
12 23 176 73.47 79.30 82.95 71. 79
MEAN 30.00 163.98 62.47 67.68 71. 47 62.49
SD 3.44 4.97 7.35 8.13 8.64 8.24
Each volunteer was examined by an obstetrician and
completed a £amiliarization and training session with
University of North Carolina Exercise Physiology
Laboratory in Fetzer Gymnasium. 12-lead resting ECG and resting blood pressure screening were obtain~d along with medical history and informed consent documentation (Appendix A>. Each subject was reimbursed $75.00 per session to pay for
transportation, parking, and babysitting. The post partum trials were conducted 8 to 10 weeks post term to ensure all applicable physiological changes of
pregnancy were gone.
Predicted Max Land Trials
The subjects reported to the laboratory for a land bicycle ergometer test one to seven days prior to each immersion trial. A target heart rate of 60¾ predicted maximum was estimated by the Karvonen's formula
(Karvonen, Kentala, & Mustala, 1957):
Hr.. = HR,.•., + O. 60 ( 220 - age - HR,.•.,
>.
Incremental exercise starting at O workload andincreasing 150 Kpm every 3 minutes was conducted until the target heart rate was reached. Oxygen uptake was taken continuously with the VOe at the final minute of target heart rate used for the immersion exercise
f = a/VOa - 0.274/0.000008 f = pedal frequency
VDe = oxygen uptake in L/min.
23
Individual 60¾ predicted maximal heart rates and oxygen uptakes are presented in Appendix 8.
Immersion Trials
Upon reporting to the laboratory for an immersion trial, the subject lay undisturbed on her side on a cushioned table for 20 minutes. Following minute 20 of rest, basal metabolic rate was obtained by having the subject breathe through a mouthpiece attached to a meteorlogic balloon for 7 minutes. The subject then left the room to empty her bladder and insert the
rectal probe. Upon return, ECG leads were attached in the CMs position. Skin temperature thermistors on nonexercising areas of the shoulder (out of water> and back (in water> were then attached with water proof
tape by the investigators. The shoulder thermistor was attached to the right trapezius muscle 2 to 4 ems below the acromium process of the scapula. The back
thermistor was attached to the left flank of the latissimus dorsi muscle 8 to 10 ems above the illiac crest.
tank's water temperature was set at 30~.5 degrees Celsius to ensure a thermoneutral exercise environment £or untrained swimmers <KcMurray & Horvath, 1979 _>. The water level was set at the height of the xiphoid
process to closely approximate the same circulatory ef£ect on the heart as horizontal swimming <Risch, Koubenec, Beckman, Lange, & Gauer, 1978). Core and skin temperatures, heart rate, and blood pressure measurements were recorded every 5 minutes. Oxygen uptake was taken £or 1 minute at the 18th minute of immersion.
Immersion exercise was then per£ormed £or 20 minutes using an underwater bicycle ergometer as
modified by Morlock & Dressendorfer (1974). Based on the previous land trial, pedal frequency was set to achieve exercise at 60¾ of predicted maximal aerobic capacity. A metronome regulated the pedal frequency during exercise. Core and skin temperature, heart rate, blood pressure, and oxygen uptake were monitored every 5 minutes.
25
Instrumentation
A calibrated Monarch Model 668 stationary bicycle ergometer was used £or the predicted max land trials. Heart rate was recorded by a Hewlett-Packard Model
1500B electrocardiogram and monitored continuously with a Nihon-Kohden Li£escope 8. Land exercise, immersion rest, and immersion exercise oxygen uptake were monitored by having the subject breathe through a Collins triple J valve with the inspired side 0£ the valve connected to a Parkinson-Cowan dry gas meter. The expired side was connected to a mixing chamber which was continuously sampled by an Applied
Electrochemical Inc. S-3A oxygen analyzer and Beckman LB-2 in£rared carbon dioxide analyzer leading to
Quintron and Fisher chart recorders. All equipment on the metabolic measurement cart was pre and post
calibrated £or accuracy using gases 0£ known Oa and COa content. Oxygen uptake was obtained £rem measurements 0£ ventilation, FEOa, and FECOa by standard £ormulas. Basal metabolic rate was determined by having the
A modified Monarch stationary bike with the friction belt removed was used for the immersion
exercise trials <Morlock & Dressendorfer, 1974). The modification involved removing the friction belt and using changes in pedal frequency <RPM) to induce
resistance and control energy expenditure. The water in the 17,625 liter immersion tank was circulated and filtered to ensure adequate mixing and maintenance of temperature before the immersion trials and during immersion rest. The water heater and filter were turned off during exercise so the subject could hear the metronome as the water was being sufficiently mixed by the subject's leg movement. A Franz Model 6648
electric metronome with auditory and visual stimuli was used to set pedal frequency. Blood pressure was
monitored by acoustic sphygmomanometer. Core, back, and shoulder temperatures were measured by
thermocouples leading to a Yellow Springs Model 44 JAX telethermometer. The telethermometer was post
calibrated using known water temperatures similar to the temperatures recoided during testing.
Statistical Analysis
temperature, mean body temperature, heart rate, and systolic blood pressure. The independent variables were pregnancy status, activity, and time.
Results and discussion will £ocus on pregnancy and activity main e££ects and pregnancy* activity
27
interactions. Data had to be collected every 5 minutes to get an accurate representation 0£ these variables. Heat storage was analyzed using 4 x 3 completely within-subjects analysis 0£ variance <ANOVA>. The independent variables were pregnancy status and
activity. Exercise heat conductance was analyzed using 4 x 4 completely within-subjects analysis 0£ variance
CANOVA). The independent variables were pregnancy status and exercise time. The £allowing dependent variables were analyzed using one-way
subjects-by-trials analysis 0£ variance CANOVA) with pregnancy status as the independent variable:
immersion heat insulation and caloric energy
expenditure during 20 minute periods 0£ land rest, immersion rest, and immersion exercise. Measures of statistical signi£icance were reported to the .05 level 0£ con£idence. A student's Newman-Keuls post hoc test was applied to determine where the signi£icant
RESULTS
Data for all dependent variables are presented
graphically in Figures 1 to 11. Descriptive statistics
and ANOVA summary tables aie given in Appendix C and individual data in Appendix 8.
Results
Core temperatures (°C) during immersion rest,
immersion exercise, and recovery did not change
significantly as a function of pregnancy status <Figure
1) but recovery core temps (37.3~.38) were higher (p<.0096) than both immersion (37.2~.34) or exercise
(37.2~.41). Twenty minutes of immersion showed a
similar decline in all trials. Core temps increased
appoximately .5 °C during exercise to peak at minute 20
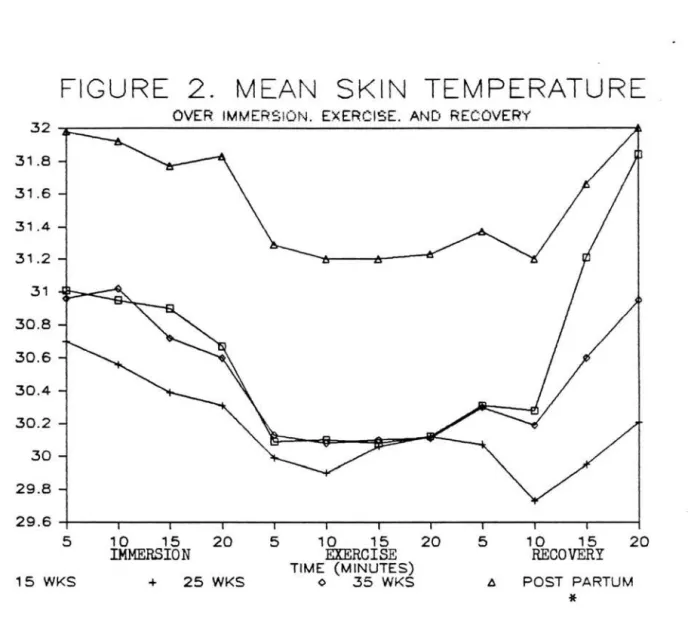
or early in recovery. Mean skin temperatures (Figure
2) during postpartum (31.6~.85) were higher (p<.0001) by approximately 1.13 °C than during any of the
pregnancy trials. Immersion (31.0~.89>, exercise
(30.4~.91), and recovery (30.7~1.18) skin temps were all significantly different (p<.0001). A gradual
decrease during immersion was followed by skin temps
back to resting levels occured after minute 10 of
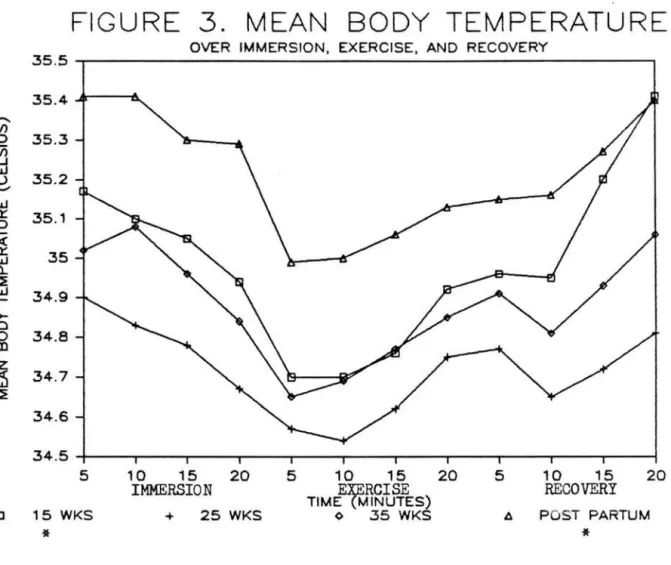
recovery. Postpartum (35.2~.37) and 15 week
(35.0~.45) mean body temperatures (Figure 3) were
higher (p<.0031) than weeks 25 (34.7~.50) or 35 (34.9~.35) of gestation. Exercise body temps
29
(34.8~.43) were lower (p<.0001) than both immersion
(35.1~.42) or recovery (35.0~.49). Mean body and mean
skin temps showed similar trends of postpartum
remaining higher and 25 weeks remaining slightly lower
across activities.
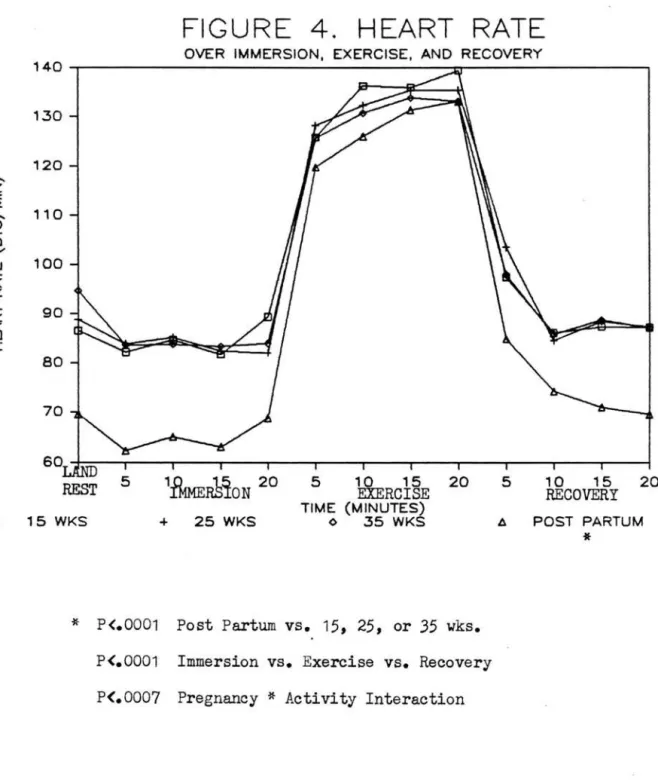
Heart rates <Figure 4) were lower (p<.0001) by approximately 13 bts/min during postpartum (89.1~29.2>
than during the 3 pregnancy trials. Immersion
(79.3~13.1>, exercise (131.5~11.0), and recovery
(86.5~14.4) heart rates were all different (p<.0001)
while remaining nearly identical with gestational
timing. A significant pregnancy* activity interaction
(p<.0007) revealed that while immersion and recovery heart rates were lower during postpartum, exercise heart rates did not change with pregnancy status.
Systolic blood pressures (mm Hg> did not change
significantly with advancing pregnancy <Figure 5). Immersion (95.1~7.8), exercise (118.7~12.3), and
interaction (p<.0243) revealed that 35 week immersion
systolic blood pressures dropped slightly lower than recovery.
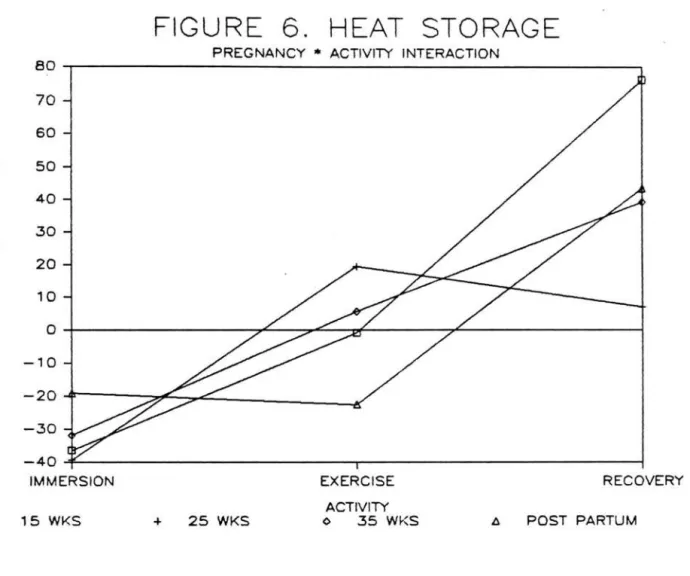
Heat storage (Keala/hr) did not change
signi£icantly with advancing gestation (Figure 6). Immersion (-31.8~40.7), exercise (0.35~54.2>, and
recovery (42.0~72.3) heat storage were all
signi£icantly di££erent (p<.0001). All trials showed a
heat loss during immersion and a heat gain during
recovery. Postpartum and 15 weeks showed a slight
heat loss during exercise while 25 and 35 weeks showed a slight heat gain. Large standard deviations were noted indicating considerable variance across subjects.
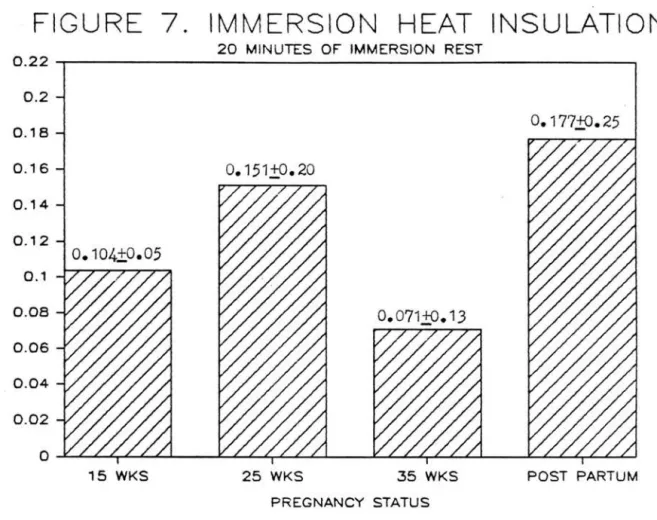
Immersion tissue insulation <Figure 7) did not
signi£icantly change with pregnancy status (overall mean, 0.128~.18 °C
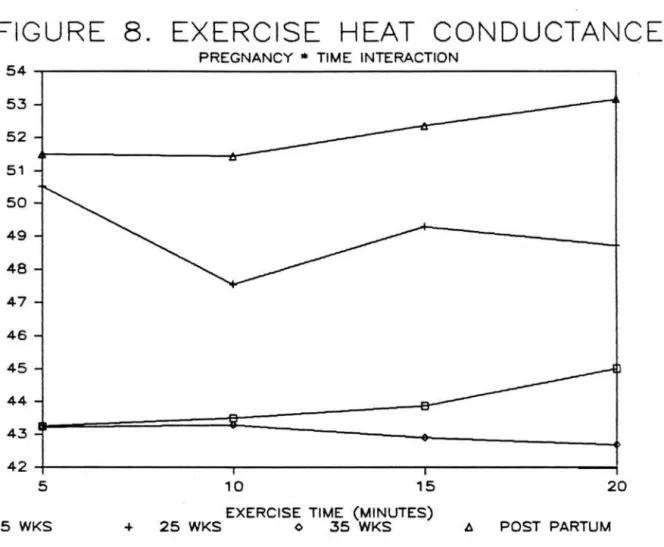
*
hr/Kcal). Exercise tissueconductance (Figure 8) showed no di££erence with pregnancy status or exercise time (overall mean,
46.94~14.1
°c
*
hr/Kcal). Postpartum and 25 week heat exchanges during immersion and exercise were slighly higher than responses during 15 or 35 weeks.Land rest energy expenditures (Kcals/20 min) were lower (p<.0001) during postpartum (19.8~3.2> and 15 weeks (19.4+4.2) than during weeks 25 (25.5+2.5) or 35
(27.8~3.6) 0£ gestation (Figure 9). Immersion rest
approximately 10.5 Kcals/20 min during postpartum while remaining nearly identical across gestation
(pregnancy mean, 39.9~6.9). Exercise energy
31
37.6
37.5
,-...,
(/) :)
vi 37.4
...J w
(.) '-"
w 37.3 a:
~
w 37 .2CL
~
~
w 37.1
a:
0 (.)
37
5
C 15 WKS
FIGURE 1. CORE TEMPERATURE
OVER IMMERSION, EXERCISE, AND RECOVERY
10 15 20 5 10 15 20
EXERCISE 5 10 15 20
IMMERSION
TIME (MINUTES) RECOVERY
+ 25 WKS o 35 WKS POST PARTUM
33
FIGURE 2. MEAN SKIN TEMPERATURE
OVER IMMERS1m-..J. EXERCISE. ANC:, RECOVERY 32
31.8
,,,,...,.
31.6
(/)
:,
~
31.4w
u
31.2
'--" w Q:
31 ::::>
~
30.8w Q_
::E 30.6
~
z
30.4~
(/)
z
30.2 ~30
::E
29.8
29 .6
5 10 15 20 5 10 15 20 5 10 15 20
IMMERSION
EXERCISE
RECOVERY
TIME (MINUTES)
□ 15 WKS + 25 WKS 0 35 WKS ll POST PARTUM
*
35.4 r"'>..
(/)
:> 35.3
ln
ci
U 35.2
'-'
w
~ 35.1
~
ffi
35 Cl.~
~ 34 .9
b
~ 34.8
z
~ 34.7 ~
34 .6
FIGURE 3. MEAN BODY TEMPERATURE
5
OVER IMMERSION, EXERCISE, AND RECOVERY
10 15 20
IMMERSION 5 10
EXERCISE
15 20 TIME (MINUTES)5 10 15 20
RECOVERY
C 15 WKS -+- 25 WKS ◊ .35 WKS POST PARTUM
*
*
,..._
z
~'
~
'-"'
~
~
~ ~ I
a
35
FIGURE 4. HEART RATE
OVER IMMERSION, EXERCISE, AND RECOVERY 140
1.30
120
110
100
90
80
70
60
REST 5 1 0
EXERCISE
1 5 20 5 1 0RECOVERY
1 5 20TIME (MINUTES)
15 WKS + 25 WKS <> .35 WKS !!,. POST PARTUM
*
* P<.0001 Post Partum vs. 15, 25, or
35
wks.
P<.0001 Immersion vs.
E
xercise vs. Recovery
-
C1'I
E E
..., w er: ::> V) U') w
0::
Cl. 0 0
g
CD () :J
~
>-U')
□
FIGURE 5. SYSTOLIC BLOOD PRESSURE
OVER IMMERSION, EXERCISE, AND RECOVERY 130 ---,
125
,20
115
110
105
REST
15 WKS
5 10 15 20 5 10 15 20
IMMERSION
EXERCISE
TIME (MINUTES)
5 10 15 20
RECOVERY
+ 25 WKS <> 35 WKS l:l POST PARTUMP<.OOO1 Immersion vs.
Exercise
vs. Recovery
70 60
50
40
30 20
10
-10
-30
FIGURE 6. HEAT STORAGE
PREGNANCY • ACTIVITY INTERACTION-40---37
IMMERSION EXERCISE
ACTIVITY
RECOVERY
□ 15 WKS + 25 WKS <> 35 WKS A POST PARTUM
*
z
0~
::,
(/)
z
~
I
w
::,
(/) (/)
~
FIGURE 7. IMMERSION
HEAT INSULATION
20 MINUTES OF IMMERSION REST 0.22
-r---,
0.2
0.18
0.16
0.14
0.12
0.1
0.08
0.06
0.04
0.02
15 WKS
o.
177,±0. 2525 WKS 35 WKS POST PARTUM
39
FIGURE 8. EXERCISE HEAT CONDUCTANCE
PREGNANCY*
TIME INTERACTION54
-
...J 53<
(.)
~
'
0::: 52I 51
*
0(.) 50 ...,
w 49
(.) z
~ 48
(.) =>
0 47
z 0
(.) 46
~
45I
w 44 => (/) (/)
.:=
4342
5 10 15 20
EXERCISE TIME (MINUTES)
z
~ 0N
'
(/)w ~
0
5
g
S2
FIGURE 9.
35
30
25
20
15
10
5
0
15 WKS
*
BASAL ENERGY EXPENDITURE
20 MINUTES OF LAND REST27. 81±,3. 61
25 WKS 35 WKS
PREGNANCY STATUS
POST PARTUM
*
41
FIGURE 10. IMMERSION
ENERGY EXPENDITURE
z
~
0
N
'
(/)w
~
0
~
g
~
20 MINUTES OF IMMERSION REST
50---,
40
30
20
10
4
1
.47±6.6
6
15 WKS 25 WKS 35 WKS
PREGNANCY STATUS
*
P(.0005 Post Partu m vs. 15,25,
or35
wks.POST PARTUM
FIGURE
1 1 .
EXERCISE
ENERGY EXPENDITURE
20 MINUTES OF IMMERSION EXERCISE
150
140
130
120 117.85±9.29
, , 0
z
~ 100
0 90
N
'
(/) 80 w0: 70 0
-;i
60 (.)g
50~ 40
30
20
10
0
15 WKS 25 WKS 35 WKS POST PARTUM
CHAPTER V
DISCUSSION
The purpose of this investigation was to determine
if the thermoregulatory, metabolic, and cardiovascular
responses of women to immersion and immersion exercise change with advancing gestation in comparison to post parturition (nonpregnant status). Pregnancy causes an
increase in body weight along with complex thermal,
metabolic, hormonal, and cardiovascular changes. The
heat dissipating capacity and hydrostatic pressure of
the water environment may effect maternal responses to
rest and exercise.
Resting heart rates on land progressively
increased with gestation to peak 25 bts/min higher at 35 weeks above postpartum (70 bts/min). Basal
metabolic rates increased from 0.97 Keala/min during
postpartum to 1.39 Keala/min at 35 weeks to meet increasing demands of the fetus and placenta <Pritchard
& MacDonald, 1980).
Twenty minutes of immersion rest in 30° C water
produced lower heart rates than land rest with post partum remaining 19 bts/min lower than the pregnancy
venous return and stroke volume allowing heart rate to
decrease and still maintain the needed cardiac output
(Farhi & Linnarsson, 1977). It is interesting to note
that immersion heart rates did not change with
gestational timing. The normal fetal position applies
pressure to the inferior vena cava <Pritchard &
MacDonald, 1980) which may contribute to increase resting heart rates with advancing gestation. The weight-supporting capacity of the water may have reduced this pressure, stabilizing immersion heart
rates throughout pregnancy.
Immersion rest produced higher energy expenditures
than land rest in all trials. The elevated resting
metabolic rates were caused by the water environment
being cooler than body temperature, thus increasing the
need for heat production (DuBois, Ebaugh, & Hardy,
1952). Although pregnancy resulted in progressive
increases in BMR and heart rate on land, caloric costs
in the water remained nearly identical throughout gestation. The lowest immersion energy costs during
pregnancy occured at 35 weeks even though BMR was the
highest. The greater mass 0£ the pregnant female
results in a smaller surface area/mass ratio which
reduces heat loss in the water CMcMurray & Horvath,
1979). Coupled with the increase in £etal metabolism,
45
maintain thermal balance. Water immersion provided smaller maternal metabolic increases near term with progeaaively larger bradycardiaa to maintain constant metabolic and cardiovascular functioning.
Core temperature (°C), heat storage (Kcals/hr), and tissue insulation (°C x hr/Kcal) during immersion rest did not significantly change with pregnancy
status. Core and mean skin temps during all pregnancy trials decreased slowly over 20 minutes indicating
vascular resistence was attempting to maintain inner body heat. This probably represented a redirection 0£ blood flow from peripheral to central, thus maintaining fetal equilibrium. Mean skin temps remained close to the water temp (30~5° C) as expected <Nadel, et al., 1974). However, akin temps were 1.13° C higher during postpartum indicating more skin blood flow as a heat loss was not as critical. The water worked against the increased cutaneous blood flow normally seen during
pregnancy (Spetz, 1964) to regulate heat dissipation generated by increased fetal metabolism. Maternal vascular compliance reduced skin blood flow and heat loss during immersion in water cooler than body temperature.
rates normally produce proportionally lower predicted oxygen uptakes using Karvonen's formula (Karvonen, et al., 1957
>.
Although resting heart rates increased with pregnancy, 60¾ predicted VOe 's did not change during weight-supported bicycle exercise.Exercise heart rates in the water did not change across trials due to use of Karvonen's formula. With the higher resting heart rates and blood volumes of pregnancy (Pritchard, 1965), smaller increases (HR •• -HRr•••) were required to adjust to the same relative
steady state work load. Heart rates during immersion exercise averaged 17 bts/min lower than the land trials at the same oxygen uptake. This difference was related to the hydrostatic pressure of the water enhancing
venous return and stroke volume. This may provide less stress to the fetus as fetal heart rate will increase 1 bt/min per 10 bt/min rise in maternal heart rate during land exercise <Dressendorfer & Goodlin, 1980).
Exercise energy expenditures in the water did not significantly change with pregnancy status (overall
mean, 113 Kcals/20 min). This concurs with Knuttson &
47
Systolic blood pressures did not change with
pregnancy status during either immersion (overall mean, 95 mm Hg) or exercise (overall mean, 119 mm Hg). _ Blood volume is increased during pregnancy as more fluid is mobilized to meet demands of the added body weight
(Pritchard, 1965). Immersion causes large fluid shifts from peripheral to central and increases plasma volume
(Kloska & DuBois, 1978). Vascular functioning was
apparently maintained by trade offs of the increases in blood volume and cardiac output and a vasodilation to supply the working muscles and maintain body
temperature <Rowell, 1974). The slightly lower
systolic blood pressures during exercise at 25 weeks reflect the enhanced vasodilation normally seen during the second trimester (Pritchard & MacDonald, 1980; Hytten & Leitch, 1964).
Immersion exercise at 60¾ of predicted VOa max for 20 minutes over advancing pregnancy did not effect core temperature responses. Core temps increased less than .5° Cover 20 minutes suggesting thermal balance was maintained. The 3 pregnancy trials showed lower
(p<.0001) skin temps than postpartum indicating a central vascular shift keeping blood around the fetus and placenta. The heat dissipating capacity of the
following 20 minutes of immersion. Exercise heat
conductance (°C • hr/Kcal) did not change significantly
but was slightly higher during postpartum.
Calculations of heat conductance are based on skin to
water temperature differences, therby reflecting the higher postpartum skin temps. Skin temps were lowest
during 25 weeks gestation with exercise heat
conductance the highest of the pregnancy trials as the
body adjusted to maintain the already elevated body
temperature (Wilson, et al, 1980).
Exercise heat storage, dependent upon body weight
and the change in mean body temp, did not change
significantly with pregnancy status (overall mean, 0.35 Kcals/hr). Large standard deviations were noted due to
anthropometric differences in the subjects. irends of
slight heat losses during postpartum and 15 weeks and
slight heat gains during 25 and 35 weeks were
influenced by the body weight and basal temperature
changes of pregnancy. As maternal weight increased
with pregnancy, mean body temps changed less to maintain thermal equilibrium. Heat dissipation was
mostly by convection <Nadel, et al., 1974) with the high water turbulence during underwater cycling.
Convective heat loss was higher during exercise than
during immersion rest as heat transfer was assisted by
49
immersion exercise was maintained by vascular
adjustments to the water temperature and metabolic heat production.
Twenty minutes 0£ land rest with subject wrapped in a blanket immediately £allowed exercise. No signi£icant di££erences in thermal £unctioning across trials indicated normal recoveries with advancing gestation. Core temps decreased slowly a£ter peaking at minute 20 0£ exercise. With the body no longer being cooled by the water, skin temps slowly increased. The combined e££ect 0£ the reduced heat loss and the slightly elevated core temps increased heat storage
(Keels/hr) in all trials. Skin temps did not increase, however, until a£ter minute 10 ensuring that inner body heat would be maintained under the new ambient
conditions. Although not signi£icant, 25 weeks gestation showed the lowest heat gains in recovery
£allowing the highest heat gains and lowest skin temps during exercise. This can be attributed to the drop in vascular tone <Hytten & Leitch, 1964) and increase in body temperature (Wilson, et al., 1980) during the second trimester.
Heart rates recovered normally remaining
This small vasodilation can be attributed to the higher blood volumes of later pregnancy remaining out in the periphery.
Conclusions
Thermal, metabolic, and cardiovascular functioning was not hindered by the physiological changes of
pregnancy during immersion rest and exercise in 30° C
water. This sharply contrasts data by Jones, et al. (1985) where 25 minutes of treadmill running elevated
.
rectal temperatures to the dangerous level of 39° C in conditioned pregnant women during moderate aerobicexercise. Maternal core temperatures and heat storage
were maintained with advancing gestation due to the heat dissipating capacity of the water and maternal skin temperature adjustments. Skin temperatures were 1.1° Clower during gestation to prevent heat losses during immersion rest and heat gains during immersion exercise. Metabolic costs and heart rates were similar across pregnancy trials during weight-supported
immersion exercise. This concurs with Knuttson &
Emerson (1974) that nonweight-bearing bicycle exercise
will not increase VOa with increasing body weight during pregnancy and extends their results to exercise in the water. The hydrostatic pressure of the water
predicted max land trials at the same exergy expenditure. The heat dissipating capacity and hydrostatic pressure of the water provided a safe environment for aerobic exercise during pregnancy.
Directions for Future Research
51
This study examined immersion exercise throughout pregnancy at one intensity (60¾ predicted VOe max), duration (20 minutes), and water temperature (30° C). Research expanding on these parameters and directly comparing responses to tethered swimming and land exercise bouts would further establish swim
conditioning as beneficial and safe for pregnancy women.
The use of target heart rate has become a popular method of regulating exercise intensity. Sixty to ninety percent of maximal heart rate is required to achieve cardiovascular conditioning (American College of Sports Medicine, 1980). Wiswell, Artal, Romen, &
Kammula (1985) suggest that maximal heart rates during pregnancy may be lower than estimated by the standard Karvonen's formula (Karvonen, et al., 1957). Exercise heart rates are already lower in water than on land
<McArdle, et al., 1971). This presents a major
rates during immersion and land exercise of various
intensities throughout gestation would set ranges for
BIBLIOGRAPHY
Abrams, R., Caton, D., Clapp, J., & Barron D. H.
(1970). Thermal and metabolic £eatures 0£ li£e in utero. Clinical Obstetrics and Gynecology, 13, 549-564.
Adamsons, K. (1966). The role 0£ thermal £actors in £etal and neonatal life. The Pediatric Clinics of North America. The Newborn I, 13(3), 599-619.
Adamsons, K., & Towell, M. E. (1965). Thermal homeostasis in the £etus and newborn. Anesthesiology, 26, 531-548.
Alexander, G., & Williams, D. (1971). Heat stress and development 0£ the conceptus in domestic sheep. Journal 0£ Agricultural Science, 76, 53-58. Alsop, F. M. (1919). The ef£ect of abnormal
temperatures upon the developing nervous system in the chick embryo. Anatomical Record, 15, 307-331. American College 0£ Obstetricians and Gynecologists.
(1985). Exercise during pregnancy and the postnatal period. Washington, D. C.: ACOG. American College of Sports Medicine. (1978). Position
statement on the recommended quantity and quality of exercise £or developing and maintaining fitness in healthy adults. Medicine and Science in Sports and Exercise, 10(3), vii-x.
American College 0£ Sports Medicine. (1980). Guidelines £or graded exercise testing and exercise
prescription. Philadelphia: Lea & Febiger.
Artal, R., & Wiswell, R. (1985). Exercise in pregnancy. Baltimore: Williams & Wilkins.
Artal, R., Platt, L. D., Sperling, M., Kammula, R. K., Jilek, J., & Nakamura, R. (1981). Exercise in pregnancy I: Maternal cardiovascular and
Artal, R., Romen, Y., Paul, R. H., & Wiswell, R. (1984). Fetal bradycardia induced by maternal exercise. Lancet, 82, 258-260.
Astrand, P. 0., & Rodahl, K. (1986). Textbook 0£ work physiology (3rd ed.). New York: McGraw-Hill.
Avellini, B. A. , Shapiro, Y. , & Pando!£, K. < 1983 >. Cardio-respiratory physical training in water and on land. European Journal 0£ Applied Physiology, 50, 255-263.
Bell, A. W., Hales, J. R. S., King, R. B., & Fawcett,
A. A. (1983). In£luence 0£ heat stress on
exercise-induced changes in regional blood £low in sheep. Journal 0£ Applied Physiology:
Respiratory, Environmental, and Exercise Physiology, 55, (6), 1916-1923.
Boulton, M. E. (1980). Scuba diving and £eta! well-being: A survey 0£ 208 women. Undersea Biomedical Research, Z<3>, 183-189.
Bullard, J. A. (1981). Exercise and pregnancy. Canadian Family Physician, 27, 977-982.
Buskirk, E. R. (1977>. Temperature regulation with
exercise. Exercise and Sport Sciences Reviews, §.,
45-88.
Ce£alo, R. C., & Hellegers, A. E. (1978). The e££ects 0£ maternal hyperthermia on maternal and £etal cardiovascular and respiratory £unction. American Journal 0£ Obstetrics and Gynecology, 131(6), 687-694.
Clapp, J. F. (1985). Fetal heart rate responses to running in midpregnancy and late pregnancy. American Journal 0£ Obstetrics and Gynecology, 153(3), 251-252.
Clapp, J. F. (1980). Acute exercise stress in the pregnant ewe. American Journal 0£ Obstetrics and Gynecology, 136(4), 489-494.
Clapp, J. F., & Dickstein, S. (1984). Endurance exercise and pregnancy outcome. Medicine and Science in Sports and Exercise, 16(6), 556-562. Clarren, S.
R. H.,
K., Smith, D. W., Harvey,
& Myrianthopoulos, N. C.
M.A. S., ( 1979