Genetic and Behavioral Impact of Bromocriptine Mesylate Quick-Release and Dopamine Functioning on Food Reward-Related Behavior
By Peter Dinh
Senior Honors Thesis
Gillings School of Public Health - Nutrition University of North Carolina at Chapel Hill
July 25, 2019
ABSTRACT
TABLE OF CONTENTS
CHAPTER 1: STUDY AIMS AND HYPOTHESES………..……..…………4
CHAPTER 2: INTRODUCTION………..………..………6
2.1 Individual drivers of obesity……….………...6
2.2 DRD2 TaqIA A1 allele………..……….………….7
2.3 Bromocriptine………..……….………...8
2.4 Summary..……….…….………..8
CHAPTER 3: METHODS……...………..………...……10
3.1 Participants……...………..………..………..10
3.2 Overall study Visit………...……….……….…………10
3.3 Bromocriptine/placebo administration……….……….11
3.4 Genotyping..………..……….……….…..11
3.5 Measures………..………….12
3.6 Statistical analysis……….………14
CHAPTER 4: RESULTS……...………..……....………14
4.1 Ad lib intake………...15
4.2 Taste test………..……….………15
4.3 Food craving and liking (FCI) ..………..…………15
4.4 Self-reported impulsivity (BIS-15)……….….………16
CHAPTER 5: DISCUSSION……...……….………..……….17
APPENDIX A: DATA TABLES………..….…………20
APPENDIX B: FIGURES………..……..……….22
STUDY AIMS AND HYPOTHESIS
Diabetes is a major global health concern that is a result of insulin resistance, causing elevated fasting and postprandial plasma glucose concentrations (Shivaprasad & Kalra, 2011). This metabolic disorder increases the risk for cardiovascular disease and manifests health complications, such as nephropathy, retinopathy, and neuropathy, that can lead to mortality if not treated correctly (Lye & Ooi, 2018). Obesity is the leading risk factor for type 2 diabetes (T2DM) (Barnes, 2011) and eating behavior is critical for T2DM risk and management
(Yannakoulia, 2006; Booth et al., 2012). As the prevalence of obesity increases, hedonic eating behavior has become an important theme in obesity research. Hedonically-motivated eating is thought to be attributed to the reward of food consumption instead of metabolic need, and it is particularly relevant for highly palatable energy-dense foods. Intake in the absence of metabolic need is attributed to overeating past the body’s ability to maintain homeostasis to regulate a balance over time without storing excess energy. It is important to consider the brain reward system and how its dysregulation may contribute to obesity (Ziauddeen et al., 2015). Treatment options that improve eating behavior can control hunger and are useful for treatment and management of diabetes through weight regulation.
Bromocriptine mesylate quick-release (Cycloset) is a drug that was approved for
compare it to those without it. Those that carry the risk allele may benefit from bromocriptine, a dopamine agonist, in treatment for weight control.
Aim 1: To determine the effect of bromocriptine on pre-diabetic individuals on hedonically motivated behavior towards food. We expect that bromocriptine will decrease inclination towards food through increased dopamine signaling that regulates hedonically motivated behavior control.
INTRODUCTION
As of 2018, as many as thirty-four million Americans (10.5% of the US population) suffered from T2DM according to the 2020 National Diabetes Statistics Report from the CDC. This metabolic disorder increases the risk for cardiovascular disease and manifests health complications, such as nephropathy, retinopathy, and neuropathy, that can lead to mortality if not treated correctly (Lye & Ooi, 2018). The prevalence of diabetes (type 2 diabetes and type 1 diabetes) is expected to increase by 54% to more than 54 million Americans between 2015 and 2030 (Rowley et al., 2017). Obesity is a major risk factor for the development of T2DM, with key findings showing that increases in visceral and ectopic fat depots is associated with insulin resistance (Golay & Ybarra, 2005). Increased body mass can lead to dysregulation of hormones involved in blood glucose control, including glucagon-like peptide 1 (GLP-1), amylin, and
adiponectin, which may decrease the effectiveness of T2DM treatment options. Dietary
interventions to treat T2DM through weight regulation may be ineffective because adherence to a strict diet plan may be difficult for patients who are overtly susceptible to hedonic motivation to eat. Treatment options that directly target intrinsic motivators of hunger can control dietary intake and may prove to be useful for treatment and management of diabetes.
2.1 Individual drivers of obesity: Food liking, wanting and Impulsivity
Obesity is positively associated with an increased risk of developing T2DM. Being overweight or obese accounts for about 80-90% of all T2DM cases (Astrup & Finer, 2001). Eating habits are dependent on hedonic motivators that are determined by the interplay between homeostatic systems, external food environment, and genetic systems (Lutter, 2009; Morton, 2006). Obesity is commonly associated with overeating, which consists of food consumption past the point of satiation. This suggests that hedonic motivation for food consumption overrides homeostatic satiation signaling leading to food consumption in the absence of hunger. Reward pathways in the brain respond to food and food-related cues, and it has been shown that the mesolimbic dopamine system promotes learning of association
between natural rewards and the environment in which they are received (Berridge & Robinson, 1998). Therefore, food, or cues that predict it, can stimulate rapid firing of dopamine neurons and regulate behaviors toward acquiring that reward. This suggests that dopamine activity is involved in regulation of hedonically motivated behavior (Berridge & Robinson, 1998).
many studies have indicated the importance of the dopamine D2 receptor in food reward regulation in association with obesity. Low dopamine D2 receptor binding and low dopamine release has been shown to be associated with obesity, and morbidly obese individuals with the lowest D2 values had the largest body mass index (Baik, 2013). Additionally, Volkow et al. (2008) has shown that low striatal D2 receptor binding correlates with decreased metabolism in prefrontal regions that are implicated in inhibitory control (Trifilieff & Martinez, 2014). Impulsivity is a category of behaviors encompassing deficits in the ability to delay immediate gratification for future larger rewards. Too much or too little activity of D2-like receptors has been shown to induce impulsive responses (Lee et al., 2009). Studies of bulimia by Waxman (2009) and Broft et al. (2012) associated with poor inhibitory control, and food binging by Wang et al. (2011) suggest that impulsive eating and addiction share the neurobiological marker of reduced D2 receptor binding and dopamine release, especially in the striatal prefrontal region.
2.2 DRD2 TaqIA A1 allele
Variations in genes that affect dopamine signaling in the brain may increase
susceptibility to obesity and T2DM. The dopamine-D2 receptor (DRD2) gene encodes the D2 dopamine receptor, mainly expressed in the striatum. The DRD2 TaqIA A1 allele is a gene polymorphism located 10 kb downstream from the DRD2 gene that affects dopamine D2
receptor density and has been associated with obesity. Compared to the homozygous A2 allele carriers, A1 allele carriers show up to 30% reduced D2 receptor density, particularly prominent in the caudate and putamen. Obese subjects have fewer D2 receptors in the striatum compared to lean subjects, and A1 allele prevalence has been reported to be significantly higher in obese individuals(Ariza et al., 2012). It is disproportionately prevalent in the T2DM population. The A1 allele has been associated with increased food intake in both mouse and human models
2.3 Bromocriptine
Bromocriptine mesylate quick-release (Cycloset) is a drug that was approved for
treatment of T2DM in May 2009. Bromocriptine is believed to reduce post meal plasma glucose levels through suppression of glucose production by the liver. It is a D2-dopamine agonist, acting to increase activity of dopamine-D2 receptors. Bromocriptine is thought to primarily increase low hypothalamic dopamine, which will impact hedonic responsivity to food stimuli (Berridge & Robinson, 1998). In treatment of T2DM patients, it is used as a therapeutic for diet control. Efficacy of bromocriptine treatment is dependent on dopamine signaling capacity, as it is a dopamine agonist. Genetic polymorphisms that affect dopamine signaling may disrupt bromocriptine effectiveness through hormonal mechanisms. Thus, there is a potential for a personalized medicine approach to target the interaction between bromocriptine and those with the DRD2 TaqIA A1 allele, who may be especially susceptible to developing obesity and T2DM. As a dopamine D2 agonist, there is potential for bromocriptine to improve dopamine receptor functionality in A1 allele carriers. Bromocriptine has shown promising results (Cincotta et al., 1999) for its potential benefit as a therapeutic agent for weight loss (Cincotta & Meier, 1996; Kamath et al, 1997).
2.3 Summary
T2DM is a major national health concern affecting at least thirty-four million Americans in 2012. Treatment of T2DM is reliant on healthy eating behavior and healthy lifestyle choices. The determinants of eating behavior involve regulation of hedonically-motivated food signaling and genetics. The TaqIA A1 allele is a genetic polymorphism that affects dopamine D2 receptor density in those who carry it. It is highly prevalent in obese individuals and those diagnosed with T2DM. About 20-35% of the population carries the DRD2 TaqIA A1 allele, while 65% of the population is overweight or obese and at risk for T2DM development. Evidence suggests that decreased dopamine signaling is associated with decreased hedonically-motivated food regulation, which may lead to overeating and obesity. Pharmaceuticals, such as bromocriptine, are commonly used to aid in behavioral change in type 2 diabetes treatment. Bromocriptine has shown mixed results in terms of effectiveness, but there may be promise for its use in
METHODS 3.1 Participants
Seventy-four (n=74) undergraduate and graduate students from a large, public university in North Carolina were recruited through fliers to participate in a taste test of fruit flavored beverages. Forty-nine participants completed all study visits (mean ± SD age: 22.5 ± 3.0 y, 22 men and 27 women) and are included here. Participants had a mean BMI of 28.6 ± 2.6 kg/m2. The sample was 10.2% Asian, 18.4% black, 55.1% white, and 16.3% mixed race or other, based on self-reported race. Inclusion criteria were age (18-35 years), 25 < BMI < 35, as body mass greater than 35 have difficulty with comfort in the MRI, and at high risk for T2DM by virtue of elevated BMI (Gray, Picone, & Yashkin, 2015).Baseline demographics for the study
participants are summarized in Table 1. We excluded participants who reported current: a) contraindications of fMRI (e.g., metal implants, braces, or serious medical problems [e.g., known T2DM, CVD, recent stroke]), b) any previous habitual use of cigarettes or illicit drugs, or c) current major psychiatric disorders (depression, bipolar disorder, panic disorder, generalized anxiety disorder), d) pregnancy e) current weight loss dieting and/or weight fluctuations greater than 10 lbs in the previous 6 weeks. Informed written consent was obtained before data
collection; methods were approved by the University of North Carolina at Chapel Hill’s Institutional Review Board.
3.2 Study Visit Overview
Participants visited the lab in three separate visits. The first visit is the screening visit, which consists of measuring participant’s height and weight, fasting blood glucose, and collecting a saliva sample. If participants met the criteria for being included in the study, they came back to the lab over two testing visits with a two-week washout period in between. During these visits, participants were given a 1.6mg dose of either bromocriptine-QR (CyclosetTM) or a placebo following an overnight fast, paralleling the recommended dosage and time of
walked to the fMRI scan center, a 10-minute walk from the research lab, where they completed an fMRI scan while being fed chocolate milkshake. After the fMRI scan, participants returned to lab and were given 12oz of chocolate milkshake to consume while completing another round of VAS and a survey rating various snack items. 20-25 minutes after they finish their milkshake, a second blood draw was taken and participated completed a stop signal task to test for
behavioral impulsivity. 20-25 minutes after the reaction time game, participants had their blood drawn for a third time before being given a VAS taste test with four snack items and chocolate milkshake, lastly followed by measured ad lib consumption of the remainder of the snacks. An overview of the study visit can be seen in Figure 1.
3.3 Bromocriptine/placebo administration
Participants will be given either bromocriptine or placebo at each testing visit. Order of bromocriptine/placebo will be counterbalanced across the participants. A 52-week trial
assessing the overall safety of bromocriptine compared to placebo (N=3070) found the optimal dose to be 1.6mg-4.8 mg taken within 2 hours of waking (26,27). Therefore, we will schedule testing visits in the morning hours to minimize the time between participants waking and receiving the drug. To allow the drug to reach maximum plasma concentration, participants will be scanned approximately 90 minutes following drug administration (25). Our dosage (1.6mg) is based on previous reports that found a significant effect of one-time bromocriptine use on BOLD signaling in an fMRI (25) and consultation with our endocrinologist, Dr. Klett. Adverse effects related to bromocriptine include nausea, dizziness, fatigue, headache, diarrhea, and
constipation (26), particularly at higher doses than the one proposed here. However, adverse events are generally not severe; only occurring at 4% greater rate than placebo (27). This 4% is accounted for in our power calculation, and if participants indicate negative side effects, their data can be controlled for or removed as necessary. Additional details regarding the prevalence and severity of adverse events can be seen in human subjects’ section.
3.4 Genotyping
Independent investigators will score allele sizes; inconsistencies were reviewed and samples rerun when necessary. For every assay each 96-well plate will include nontemplate and DNA standards of known genotype. Genotyping was be performed by an independent genotyping service at the Nutrition Research Institute.
3.5 Measures
3.5.1 Anthropometric Measures
Body mass index (BMI; kg/m2) was used as a measure of adiposity and participant risk for T2DM. Height was measured to the nearest millimeter using a stadiometer and weight was assessed to the nearest 0.1 kg using a digital scale. Non-fasted blood glucose was assessed using a finger prick during the initial screening.
3.5.2 Taste test
Participants performed a taste test which included perceptual hedonic ratings of a variety of foods and an ad lib intake challenge. Participants first performed a taste test of 1g of each of the 4 foods (e.g. M&Ms, Skittles, popcorn, Doritos). These foods were selected as they are generally considered highly palatable and represent a variety of flavors, mouth feels and nutrient contents (e.g., sugar, fat, salt). Participants will be asked to rinse with water between tasting each of the foods and the order of presentation of the foods will be counter-balanced across the sample. In addition to the snacks, participants performed a taste test for chocolate milkshake. The milkshake was prepared as one part chocolate ice cream and one part Nesquik chocolate milk (218 kcal, 8 g fat, 23 g sugar/8 fl oz). For the taste test, participants were given a 12oz glass. Participants rated their desire to eat/drink and pleasantness for the snacks and milkshake on a visual analog scale (VAS) that ranged from -100 (“least desire imaginable”; “most unpleasant imaginable”) to 100 (“most desire imaginable”; “most pleasant imaginable”).
3.5.3 Ad lib intake
After the taste test, participants will be told that they are free to eat as many of the leftover snacks as they like, as we have to discard them after each participant (they will not be allowed to have backpacks, purses, or bags, so they do not take snacks from the room).
3.5.4 Food Craving
The Food Craving Inventory (FCI) was used to assess the degree of craving and liking of high- and low-fat/sugar foods (White et al., 2002. This scale has shown internal consistency (α = .93), 2-week test-retest reliability (r = .86), and sensitivity to detecting change (55). For the assessment of craving, it uses a rating scale from 1-5, with 1 being the lowest craving level and 5 being the highest level of craving for a specific food. For the assessment of liking, it uses a rating scale from 1-4, with 1 being the lowest liking score and 4 being the highest liking score for a specific food. Responses for all foods were averaged to create a total craving and total liking score.
3.5.5 Self-report Impulsivity
The Barratt Impulsivity Scale (BIS-15) is a 15-item self-report assessment used to assess total impulsivity, as well as the subdomains of attentional, motor, and non-planning impulsivity (Spinella, 2007). This scale has shown internal consistency (α=.81), 2-week test-retest reliability (r =.88), and discriminates between psychiatric patients and controls
(56). Individual items were scored from 1 (“Rarely/Never”) to 4 (“Almost Always/Always”) and responses were summed to create the total and subdomain scores:
3.5.6 Visual Analogue Scales
Participants will complete 5 cross-modal visual analog scales (VAS) per visit for to assess hunger, fullness, nausea and thirst and 2 additional VAS to assess the hedonics of the milkshake (pre-scan & pre-fixed consumption). The cross-modal VAS assesses these
-100mm and “Greatest imaginable hunger” at -100mm. Hedonic VAS measures of milkshake pleasantness will be used to examine the effects of bromocriptine on self-reported enjoyment of the milkshake taste. Additional VAS ratings will be used as control variables of no interest, as well as outcome variable in secondary analyses.
3.6 Statistical Analysis
Statistical analyses were performed using R (version 3.6.1, The R Foundation for Statistical Computing). Descriptive statistics to summarize means, standard deviation, and percentages were generated for variables of interest. Repeated measures ANOVAs were calculated using multilevel modeling with the nmle R package. The models tested the main effects of and interaction between drug administration (bromocriptine vs. placebo) and TaqIA allele status (A1 carriers vs. A2/A2) on the outcome variables (ad-lib snack and milkshake intake, snack and milkshake liking/pleasantness, snack and milkshake wanting/desire to eat, FCI total craving and FCI total liking score, and BIS-15 impulsivity). In all models, self-reported race and ethnicity were included to adjust for population stratification. Sex was added as a between-subjects factor to probe for interactions with drug administration and TaqIA allele status. Random effects were included at the subject level to account for individual differences in outcomes. Additional confounders (age, BMI, hunger, nausea) were adjusted for in all models. Statistical significance was set at p < 0.05.
RESULTS
4.1 Ad Lib Intake
4.1.1 Main Effects of Drug, TaqIA allele status, Sex
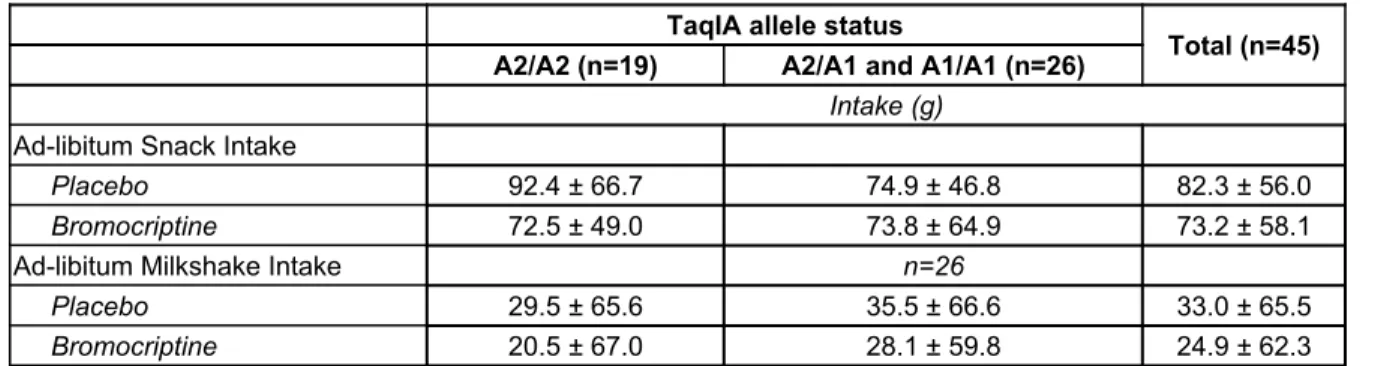
Ad lib snack and milkshake intake amounts did not significantly differ by bromocriptine vs. placebo administration or between carriers and non-carriers of the TaqIA A1 risk allele. Ad lib snack intake was higher for males compared to females (β = 50.48, t(34) = 2.11, p = 0.042), as outlined in Figure 2. There was no significant difference found for ad-lib milkshake intake between males vs. females.
4.1.2 Interaction of Drug, TaqIA allele status, Sex
There were no significant interactions between drug administration, TaqIA A1 allele status, or sex for ad lib intake of either snacks or milkshake.
4.2 Taste Test
4.2.1 Main Effects of Drug, TaqIA allele status, Sex
Food liking scores were lower in A1 allele carriers compared to non-carriers, although the results failed to reach significance (β = 15.78, t(34) = 1.92, p = 0.063). Food wanting scores were marginally lower during bromocriptine administration compared to placebo although the results were not statistically significant (β = 14.16, t(37) = 1.86, p = 0.071).
Neither snack nor milkshake liking or desire to consume were significantly different during bromocriptine vs. placebo administration. Snack and milkshake liking and desire to consume scores were also not significantly different between carriers and non-carriers of the TaqIA A1 risk allele or between males and females.
4.2.2 Interaction of Drug, TaqIA allele status, Sex
The interaction between drug administration and A1 allele status was marginally significant for food liking scores such that carriers of the A1 allele had decreased food liking during bromocriptine administration vs. placebo and non-carriers of the A1 allele had increased food liking during bromocriptine vs. placebo administration, β = -18.46, t(37) = -2.00, p = 0.052, as shown in Figure 3a. Findings are more prominent in females, although not significant (Figure 3b).
4.3 Food Craving and Liking (FCI)
Total craving and liking scores for a variety of foods as assessed by the FCI did not differ during bromocriptine vs. placebo administration. FCI scores were also not significantly different between carriers and non-carriers of the TaqIA A1 risk allele or between males and females.
4.3.2 Interaction of Drug, TaqIA allele status, Sex
For craving scores, there was a significant interaction between TaqIA allele status and sex, such that, among males, craving scores were overall higher for the risk allele carriers compared to male non-carriers (β = -0.57, t(38) = -2.06, p = 0.047; Figure 4).
The interaction between drug and TaqA1 was significant for total liking scores such that A1 had a larger decrease as a function of bromocriptine administration (β = -0.23, t(40) = -2.34, p = 0.024), as displayed in Figure 5a. For total liking scores, there was a three-way interaction between drug administration, TaqIA allele status and sex, such that male carriers of the A1 allele, during both bromocriptine and placebo administration, had lower liking scores compared to male with the A2/A2 variant. Female carriers of the A1 allele showed decreased liking scores after bromocriptine administration compared to placebo, while female non-carriers showed increased liking ratings after bromocriptine administration compared to placebo (β = 0.33, t(40) = 2.18, p = 0.035) as displayed in Figure 5b.
4.4 Self-reported Impulsivity (BIS-15)
4.4.1 Main Effects of Drug, TaqIA allele status, Sex
Total impulsivity scores for attentional, motor and non-planning as measured using the BIS-15 scale did not differ by bromocriptine vs. placebo administration. Impulsivity scores were also not significantly different between carriers and non-carriers of the TaqIA A1 risk allele or between males and females.
4.4.2 Interaction of Drug, TaqIA allele status, Sex
The interaction between TaqIA A1 allele status and sex was marginally significant for total impulsivity scores such that male A1 allele carriers had increased total impulsivity
DISCUSSION
The purpose of this study was 1) to investigate the effects of bromocriptine
administration on hedonic motivation towards food and 2) to determine the interactive effects between bromocriptine drug administration and TaqIA A1 allele status on hedonically-motivated food eating behavior and impulsivity.
For all measures (ad lib intake, liking or desire to consume snacks or milkshake, food craving, and impulsivity), we found no significant difference between within-subject measures of bromocriptine vs. placebo administration. We are unable to draw any conclusions regarding the general effect of bromocriptine on appetite from our data.
Similarly, for all measures, there was no significant difference between average scores of TaqIA A1 allele carriers compared to non-carriers. We are unable to draw any conclusions regarding the general effect of TaqIA A1 allele status on appetite from our data. Additionally, there were no significant interactions between drug administration and TaqIA A1 allele status for any of the measures tested.
We identified significant interactions between bromocriptine administration, TaqAI A1 allele status and sex. Male carriers of the TaqAI A1 allele had higher impulsivity, higher food cravings, and lower food liking scores compared to male non-carriers. Bromocriptine was found to increase food cravings in male carriers of the TaqIA A1 allele, which is the opposite of the effect that we expected from administration of bromocriptine. For females, we found that carriers of the TaqIA A1 allele had lower impulsivity compared to non-carriers. Bromocriptine was found to decrease food liking for female carriers of the A1 allele, but increase food liking for non-carriers of the A1 allele. This finding is significant for the whole sample, including both males and females, but graphical representation of the results show that the interaction is driven by females.
the finding that bromocriptine increased food liking in non-carriers was unexpected. This suggests that there may be a potential difference in interaction between the drug and carriers vs. non-carriers of the TaqIA A1 allele, but we only discovered marginal significance and further studies must be done to confirm this finding. Additionally, other covariates, such as nausea, might have played a role in these results, as nausea status had a significant effect, β = -0.20, t(38) = -2.97, p = 0.005, on milkshake wanting status.
The differences between males and females suggest that there may be an interaction between sex that contributes to the improved effectiveness of the drug in female participants compared to male participants. Male participants displayed mixed results for impulsivity, food craving, and food liking scores in response to bromocriptine. Further research should be done to explore this finding, but the use of bromocriptine as a means of weight regulation may be more promising for females compared to males.
We hypothesized that bromocriptine would decrease inclination towards food and lower outcomes for measures of liking, desire to consume, ad lib intake, and food craving. However, in our sample, bromocriptine decreased food liking only, driven mainly by female subjects. These data suggest that there may be a separate mechanism that bromocriptine acts upon to improve metabolism and promote weight and body fat loss in obese subjects, as discovered in previous studies (Cincotta & Meier, 1996; Kamath et al, 1997). We also hypothesized that bromocriptine would decrease impulsivity for carriers of the TaqIA A1 allele, while not significantly affecting impulsivity in non-carriers. Although we found no significant results for drug administration on impulsivity, we actually observed that A1 carriers had lower impulsivity than non-carriers. Research measuring the effects of varying levels of dopamine in the brain on impulsivity may help to clarify this relationship. Given the small sample size of this study, we cannot disregard the potential interaction between bromocriptine and A1 allele status, as the true effect of this relationship may not have been detected with only 49 participants.
Further limitations of this study are that we did not screen individuals for HbA1c. Instead, we used BMI as a measure of T2DM risk, which may be inaccurate. Additionally, we did not screen for eating disorders, nor could we fully control for outside environmental factors, both of which could have affected ad libitum intake, desire to eat, and hedonic ratings of beverages.
regarding the true nature of the relationship between the effects of bromocriptine and TaqIA A1 allele status on eating behavior, but we have found some potentially significant interactions. Future research should continue to explore the potential interaction between the bromocriptine drug and TaqIA A1 allele status as a means of diet control and how dopamine levels affect that relationship. Other remaining areas of interest include clarifying the effect bromocriptine has on improving reduced dopamine D2 receptor density, as well as identifying the interactions
APPENDIX A: DATA TABLES Table 1. Baseline characteristics of participants (n=49)
TaqIA allele status
Total (n=49) A2/A2 (n=20) A2/A1 and A1/A1 (n=29)
Count (Percent) Sex
Male 10 (50%) 12 (41.4%) 22 (44.9%)
Female 10 (50%) 17 (58.6%) 27 (55.1%)
Race
Black or African American 5 (25.0%) 4 (13.8%) 9 (18.4%)
Asian 0 (0%) 5 (17.2%) 5 (10.2%)
White 14 (70.0%) 13 (44.8%) 27 (55.1%)
American Indian or Alaska
Native 0 (0%) 1 (3.4%) 1 (2.0%)
More than one race 0 (0%) 2 (6.9%) 2 (4.1%)
Other or chose not to report 1 (5.0%) 4 (13.8%) 5 (10.2%)
Ethnicity
Hispanic 3 (15.0%) 5 (17.2%) 8 (16.3%)
Non-Hispanic 17 (85.0%) 24 (82.8%) 41 (83.7%)
Mean ± SD
Age (years) 22.4 ± 3.1 22.6 ± 2.9 22.5 ± 3.0
BMI (kg/m2) 28.5 ± 2.1 28.7 ± 2.9 28.6 ± 2.6
Table 2. Hedonic Ratings of Snacks and Milkshake TaqIA allele status
Total (n=47) A2/A2 (n=19) A2/A1 and A1/A1(n=28)
VAS Score (mm) Snack liking/pleasantness*
Placebo 33.1 ± 11.5 33.5 ± 18.1 33.4 ± 15.6
Bromocriptine 36.6 ± 14.5 25.7 ± 22.0 30.1 ± 19.9
Snack wanting/desire to eat*
Placebo 13.4 ± 25.2 17.8 ± 22.0 16.1 ± 23.2
Bromocriptine 11.3 ± 24.9 8.0 ± 24.3 9.4 ± 24.3
Milkshake liking/pleasantness
Placebo 12.2 ± 27.2 29.8 ± 28.8 22.7 ± 29.2
Bromocriptine 15.1 ± 38.8 28.1 ± 31.0 22.9 ± 34.6
Milkshake wanting/desire to eat
Placebo -21.6 ± 36.2 6.6 ± 41.0 -4.8 ± 41.2
Bromocriptine -18.1 ± 40.3 1.6 ± 38.4 -6.4 ± 40.0
Table 3. Ad-lib intake of Snacks and Milkshake
TaqIA allele status
Total (n=45) A2/A2 (n=19) A2/A1 and A1/A1 (n=26)
Intake (g)
Ad-libitum Snack Intake
Placebo 92.4 ± 66.7 74.9 ± 46.8 82.3 ± 56.0
Bromocriptine 72.5 ± 49.0 73.8 ± 64.9 73.2 ± 58.1
Ad-libitum Milkshake Intake n=26
Placebo 29.5 ± 65.6 35.5 ± 66.6 33.0 ± 65.5
Bromocriptine 20.5 ± 67.0 28.1 ± 59.8 24.9 ± 62.3
Table 4. FCI Total Craving and Liking Scores
TaqIA allele status
Total (n=49) A2/A2 (n=20) A2/A1 and A1/A1 (n=29)
FCI Total Craving Score
Placebo 2.08 ± 0.40 2.31 ± 0.47 2.21 ± 0.46
Bromocriptine 2.09 ± 0.38 2.38 ± 0.49 2.26 ± 0.47
FCI Total Liking Score
Placebo 2.73 ± 0.27 2.76 ± 0.38 2.75 ± 0.34
Bromocriptine 2.75 ± 0.28 2.72 ± 0.32 2.73 ± 0.30
Table 5. BIS-15 Impulsivity Measures
TaqIA allele status
Total (n=49) A2/A2 (n=20) A2/A1 and A1/A1 (n=29)
BIS-15 Total Impulsivity
Placebo 27.0 ± 4.6 27.9 ± 5.7 27.5 ± 5.3
Bromocriptine 27.6 ± 3.7 28.3 ± 4.6 28.0 ± 4.2
BIS-15 Non-planning Impulsivity
Placebo 8.9 ± 2.1 9.9 ± 2.6 9.5 ± 2.4
Bromocriptine 9.4 ± 1.7 9.5 ± 1.9 9.5 ± 1.8
BIS-15 Motor Impulsivity
Placebo 8.7 ± 2.2 8.5 ± 2.1 8.6 ± 2.1
Bromocriptine 8.6 ± 2.1 8.9 ± 2.0 8.8 ± 2.1
BIS-15 Attentional Impulsivity
Placebo 9.4 ± 2.3 9.4 ± 2.1 9.4 ± 2.2
APPENDIX B: FIGURES
(a)
(b)
Figure 2. ANOVA plots comparing ad-lib snack intake between (a) drug vs. placebo and TaqIA A1 carriers vs. carriers and (b) drug vs. placebo and TaqIA A1 carriers vs.
(a)
(b)
Figure 3. ANOVA plots comparing snack liking ratings between (a) drug vs. placebo and TaqIA A1 carriers vs. non-carriers and (b) drug vs. placebo and TaqIA A1 carriers vs. non-carriers
(a)
(b)
Figure 5. ANOVA plots comparing FCI total food liking scores between (a) drug vs. placebo and TaqIA A1 carriers vs. carriers and (b) drug vs. placebo and TaqIA A1 carriers vs.
(a)
(b)
REFERENCES
1. Baik, J.-H. (2013). Dopamine signaling in reward-related behaviors. Frontiers in Neural Circuits, 7, 152.
2. Barnard, N. D., Noble, E. P., Ritchie, T., Cohen, J., Jenkins, D. J. A., Turner-McGrievy, G., Gloede, L., et al. (2009). D2 dopamine receptor Taq1A polymorphism, body weight, and dietary intake in type 2 diabetes. Nutrition, 25(1), 58–65.
3. Barnes, A. S. (2011). The epidemic of obesity and diabetes: trends and treatments. Texas Heart Institute Journal, 38(2), 142–144.
4. Berridge, K. C., & Robinson, T. E. (1998). What is the role of dopamine in reward: hedonic impact, reward learning, or incentive salience? Brain Research. Brain Research Reviews, 28(3), 309–369.
5. Booth, D. A., Jarvandi, S., & Thibault, L. (2012). Food after deprivation rewards the earlier eating. Appetite, 59(3), 790–795.
6. Broft, A., Shingleton, R., Kaufman, J., Liu, F., Kumar, D., Slifstein, M., Abi-Dargham, A., et al. (2012). Striatal dopamine in bulimia nervosa: a PET imaging study. The
International Journal of Eating Disorders, 45(5), 648–656.
7. Burger, K. S., & Stice, E. (2011). Variability in reward responsivity and obesity: evidence from brain imaging studies. Current drug abuse reviews, 4(3), 182–189.
8. Cincotta, A. H., & Meier, A. H. (1996). Bromocriptine (Ergoset) reduces body weight and improves glucose tolerance in obese subjects. Diabetes Care, 19(6), 667–670.
9. Cincotta, A. H., Meier, A. H., & Cincotta, M. (1999). Bromocriptine improves glycaemic control and serum lipid profile in obese Type 2 diabetic subjects: a new approach in the treatment of diabetes. Expert Opinion on Investigational Drugs, 8(10), 1683–1707. 10. Defronzo, R. A. (2011). Bromocriptine: a sympatholytic, d2-dopamine agonist for the
treatment of type 2 diabetes. Diabetes Care, 34(4), 789–794.
11. Eisenberg, D. T. A., Mackillop, J., Modi, M., Beauchemin, J., Dang, D., Lisman, S. A., Lum, J. K., et al. (2007). Examining impulsivity as an endophenotype using a behavioral approach: a DRD2 TaqI A and DRD4 48-bp VNTR association study. Behavioral and Brain Functions, 3, 2.
12. Epstein, L. H., Temple, J. L., Neaderhiser, B. J., Salis, R. J., Erbe, R. W., & Leddy, J. J. (2007). Food reinforcement, the dopamine D2 receptor genotype, and energy intake in obese and nonobese humans. Behavioral Neuroscience, 121(5), 877–886.
14. Kamath, V., Jones, C. N., Yip, J. C., Varasteh, B. B., Cincotta, A. H., Reaven, G. M., & Chen, Y. D. (1997). Effects of a quick-release form of bromocriptine (Ergoset) on fasting and postprandial plasma glucose, insulin, lipid, and lipoprotein concentrations in obese nondiabetic hyperinsulinemic women. Diabetes Care, 20(11), 1697–1701.
15. Lee, J., Parish, C. L., Tomas, D., & Horne, M. K. (2011). Chronic cocaine administration reduces striatal dopamine terminal density and striatal dopamine release which leads to drug-seeking behaviour. Neuroscience, 174, 143–150.
16. Lutter, M., & Nestler, E. J. (2009). Homeostatic and hedonic signals interact in the regulation of food intake. The Journal of Nutrition, 139(3), 629–632.
17. Morton, G. J., Cummings, D. E., Baskin, D. G., Barsh, G. S., & Schwartz, M. W. (2006). Central nervous system control of food intake and body weight. Nature, 443(7109), 289– 295.
18. Nisoli, E., Brunani, A., Borgomainerio, E., Tonello, C., Dioni, L., Briscini, L., Redaelli, G., et al. (2007). D2 dopamine receptor (DRD2) gene Taq1A polymorphism and the eating-related psychological traits in eating disorders (anorexia nervosa and bulimia) and obesity. Eating and Weight Disorders, 12(2), 91–96.
19. Ooi, C. P., & Lye, M. S. (2018). Bromocriptine for type 2 diabetes mellitus. Cochrane Database of Systematic Reviews.
20. Ryan, A. S. (2000). Insulin Resistance with Aging. Sports Medicine, 30(5), 327–346. 21. Schwartz, S. S., & Zangeneh, F. (2016). Evidence-based practice use of quick-release
bromocriptine across the natural history of type 2 diabetes mellitus. Postgraduate medicine, 128(8), 828–838.
22. Shivaprasad, C., & Kalra, S. (2011). Bromocriptine in type 2 diabetes mellitus. Indian journal of endocrinology and metabolism, 15(Suppl 1), S17-24.
23. Spinella, M. (2007). Normative data and a short form of the Barratt Impulsiveness Scale. The International journal of neuroscience, 117(3), 359–368.
24. Thanos, P. K., Michaelides, M., Piyis, Y. K., Wang, G.-J., & Volkow, N. D. (2008). Food restriction markedly increases dopamine D2 receptor (D2R) in a rat model of obesity as assessed with in-vivo muPET imaging ([11C] raclopride) and in-vitro ([3H] spiperone) autoradiography. Synapse, 62(1), 50–61.
26. Wang, G. J., Volkow, N. D., Logan, J., Pappas, N. R., Wong, C. T., Zhu, W., Netusil, N., et al. (2001). Brain dopamine and obesity. The Lancet, 357(9253), 354–357.
27. Wang, G.-J., Geliebter, A., Volkow, N. D., Telang, F. W., Logan, J., Jayne, M. C., Galanti, K., et al. (2011). Enhanced striatal dopamine release during food stimulation in binge eating disorder. Obesity, 19(8), 1601–1608.
28. Waxman, S. E. (2009). A systematic review of impulsivity in eating disorders. European Eating Disorders Review, 17(6), 408–425.
29. White, M. A., Whisenhunt, B. L., Williamson, D. A., Greenway, F. L., & Netemeyer, R. G. (2002). Development and validation of the food-craving inventory. Obesity
research, 10(2), 107–114.
30. Yannakoulia, M. (2006). Eating behavior among type 2 diabetic patients: a poorly recognized aspect in a poorly controlled disease. The review of diabetic studies : RDS, 3(1), 11–16.