Copyright © 2000, American Society for Microbiology. All Rights Reserved.
Expression of a Gene Encoding the Major Antigenic Protein 2
Homolog of
Ehrlichia chaffeensis
and Potential
Application for Serodiagnosis
A. RICK ALLEMAN,
1* ANTHONY F. BARBET,
2MICHAEL V. BOWIE,
2HEATHER L. SORENSON,
1SUSAN J. WONG,
3ANDMYRIAM BE´LANGER
2Departments of Physiological Sciences
1and Pathobiology,
2College of Veterinary Medicine, University of Florida,
Gainesville, Florida 32610, and Wadsworth Center, New York State Department of Health, Albany, New York 12201
3Received 8 February 2000/Returned for modification 3 May 2000/Accepted 10 July 2000
The major antigenic protein 2 (MAP2) homolog of
Ehrlichia chaffeensis
was cloned and expressed. The
recombinant protein was characterized and tested in an enzyme-linked immunosorbent assay (ELISA) format
for potential application in the serodiagnosis of human monocytic ehrlichiosis. The recombinant protein, which
contained a C-terminal polyhistidine tag, had a molecular mass of approximately 26 kDa. The antigen was
clearly identified by Western immunoblotting using antihistidine antibody. However, immune sera failed to
react with the recombinant on immunoblots when the antigen was denatured by heat or reduced using

-mercaptoethanol. The recombinant MAP2 (rMAP2) was used in an ELISA format with 60 blinded serum
samples. Twenty of the serum samples were previously demonstrated to contain antibodies reactive with
E.
chaffeensis
by indirect immunofluorescence assays (IFAs). The remaining 40 samples were seronegative. All
samples negative by IFA were also found to be negative for antibodies against the rMAP2 of
E. chaffeensis
by
using the ELISA. Only 1 of 20 IFA-positive samples tested negative in the rMAP2 ELISA. There was 100%
agreement using IFA-negative samples and 95% agreement using IFA-positive samples, resulting in a 97.5%
overall agreement between the two assays. These data suggest that the rMAP2 homolog of
E. chaffeensis
may
have potential as a test antigen for the serodiagnosis of human monocytic ehrlichiosis. To our knowledge, this
recombinant is unique because it is thus far the only
E. chaffeensis
recombinant antigen that has been shown
to work in an ELISA format.
Clinical findings associated with
Ehrlichia chaffeensis
infec-tion (human monocytic ehrlichiosis [HME]) and infecinfec-tion with
the human granulocytic ehrlichiosis agent are similar,
consist-ing of fever, headache, and myalgia. Common laboratory
find-ings for both diseases include leukopenia, thrombocytopenia,
and elevated levels of hepatic enzymes (6). There is
consider-able antigenic cross-reactivity between the
Ehrlichia
spp. and
other closely related species such as
Cowdria ruminantium
(4,
5, 8, 10, 13, 14, 19). Although a newly developed recombinant
enzyme-linked immunosorbent assay (ELISA) for the
diagno-sis of human granulocytic ehrlichiodiagno-sis appears to have good
specificity (7), the presently available serologic tests for HME
are unable to discriminate between infections caused by
dif-ferent
Ehrlichia
spp. Although several recombinant proteins
have been evaluated for the serologic diagnosis of HME (16,
20, 21), the immunofluorescence assay (IFA), using in
vitro-cultured, whole
Ehrlichia
organisms, is still the preferred
lab-oratory method of serodiagnosis (18). Improved specificity for
diagnosis of
E. chaffeensis
infection may be available by use of
immunoblot analysis (3); however, DNA sequence analysis is
presently the only reliable method for specific identification of
Ehrlichia
spp.
A 19-kDa protein, major surface protein 5 (MSP5), of
Anaplasma marginale
has been produced as a recombinant
protein and is currently being used as a diagnostic test to
identify
A. marginale
-infected cattle (9). This protein was
found to be conserved in all recognized
Anaplasma
species,
and when used with monoclonal antibodies in a competitive
ELISA format, it was able to detect cattle persistently infected
with
A. marginale
but not other closely related organisms (9,
17). A 21-kDa protein with 55.5% amino acid identity to MSP5
of
A. marginale
was identified in the closely related rickettsia
C.
ruminantium
(11). The gene encoding this protein, major
an-tigenic protein 2 (MAP2), was isolated, cloned, and expressed
in
Escherichia coli
. The recombinant MAP2 (rMAP2) was
shown to be recognized by sera from cattle, sheep, and goats
infected with
C. ruminantium
(11). The
map
2 gene, like the
msp
5 gene of
A. marginale
, was also found to be well conserved
between various isolates of
C. ruminantium
, and homologs of
the
map
2 and
msp
5 genes from both
Ehrlichia canis
and
E.
chaffeensis
were identified (2). Amino acid sequence analysis of
the MAP2 homologs from
E. chaffeensis
and
E. canis
revealed
83.4 and 84.4% identities, respectively, with MAP2 from
C.
ruminantium
(2).
In this study, we report the cloning and expression of the
map
2 homolog from
E. chaffeensis
and examine the potential
value of the rMAP2 homolog for the serodiagnosis of HME.
MATERIALS AND METHODS
Source ofE. chaffeensisorganisms and DNA.E. chaffeensis(Arkansas isolate) was kindly provided by Jacqueline E. Dawson and James G. Olsen, Centers for Disease Control, Atlanta, Ga. Organisms were grown in the canine macrophage cell line DH82 in Eagle’s minimum essential medium containing 10% fetal bovine serum, 26 mM sodium bicarbonate, and 2 mML-glutamine at 34°C. Cells
were harvested when 90 to 100% of them were infected, and ehrlichiae were purified as previously described (5). Genomic DNA ofE. chaffeensiswas isolated by treatment of purified organisms with 5 mg of lysozyme per ml, 100g of proteinase K per ml, and 2% (wt/vol) sodium dodecyl sulfate (SDS), followed by phenol-chloroform extraction and ethanol precipitation (11).
* Corresponding author. Mailing address: Department of
Physiolog-ical Sciences, College of Veterinary Medicine, University of Florida,
Box 100103C, Gainesville, FL 32610. Phone: (352) 392-4700, ext. 5858.
Fax: (352) 392-1769. E-mail: [email protected]
.EDU.
3705
on May 15, 2020 by guest
http://jcm.asm.org/
Amplification of themap2 gene ofE. chaffeensis.Primers, which corresponded to the sequences encoding the predicted mature protein of themap2 gene, were synthesized by Genosys Biotechnologies Inc., The Woodlands, Tex. The forward primer ARA3 (5⬘GGCAATTTTTCTAGGATATTCCTACGTAACA 3⬘) and reverse primer ARA4 (5⬘GAGATATTGTTTTATTATAGATAGTAACTT 3⬘) were designed for in-frame insertion of amplicons into the pTrcHis2-TOPO vector (Invitrogen Corporation, Carlsbad, Calif.). The beginning of the predicted mature protein corresponds to nucleotide 46 of the open reading frame. The nucleotide sequence of themap2 gene ofE. chaffeensishas been previously reported (2) and assigned GenBank accession number AF117731. Amplification was performed usingTaqDNA polymerase in order to produce amplicons with the necessary 3⬘A overhangs needed for ligation into the TOPO vector. Briefly, genomic DNA (10 ng) was amplified using 0.5M concentrations of primers ARA3 and ARA4 and 1.25 U ofTaqDNA polymerase in 2 mM deoxynucleoside triphosphates–10 mM Tris-HCl (pH 8.8)–50 mM KCl–1.5 mM MgCl2. PCR assays were performed at 94°C for 3 min, followed by 10 cycles of denaturing at 94°C for 15 s, annealing at 43°C for 1 min, and extension at 72°C for 7 min. This was followed by 25 cycles of denaturing at 94°C for 15 s, annealing at 49°C for 1 min, and extension at 72°C for 7 min. A final extension step at 72°C was performed for 7 min. Amplicons were analyzed by gel electrophoresis on a 1% agarose gel in 1⫻TBE buffer (89 mM Tris, 89 mM boric acid, and 2 mM disodium EDTA).
Cloning and sequencing ofE. chaffeensis map2.Amplicons were inserted into the pTrcHis2-TOPO vector (Invitrogen Corporation) according to the manufac-turer’s recommendations. Recombinant plasmids were transformed intoE. coli (One Shot cells; Invitrogen Corporation), and transformants were grown on Luria-Bertani (LB) agar plates in the presence of ampicillin (50g/ml). Colonies were selected and incubated in LB broth in the presence of ampicillin (50g/ml) overnight at 37°C with vigorous shaking. Plasmid DNA was extracted by a rapid miniprep method (22), reconstituted in Tris-EDTA buffer (pH 8.0) containing 1.0g of DNase-free RNase per ml, and analyzed on a 1% agarose gel. Recom-binant clones containing themap2 homologs ofE. chaffeensiswere digested with restriction enzymes (NcoI andDraI) to ensure the correct orientation of the insert in the plasmid vector. Digested DNA was analyzed on a 1% agarose gel. The DNA sequences of both strands of the 570-bp insert of pTrcHis2-TOPO K1 were determined by the DNA Sequencing Core Laboratory (University of Flor-ida, Gainesville) using forward and reverse primers based on vector sequences in flanking regions.
Production and purification ofE. chaffeensisrecombinant MAP2.Transformed cells containing themap2 gene homologs ofE. chaffeensiswere incubated with vigorous shaking at 37°C in LB broth containing 50g of ampicillin per ml to an optical density at 600 nm of 0.6. Protein production was induced with 1 mM isopropyl--D-thiogalactopyranoside, and cells were incubated with vigorous
shaking at 37°C for an additional 5 h. Recombinant proteins were purified by immobilized metal affinity chromatography (ProBond resin; Invitrogen Corpo-ration) and isolated under native, nondenaturing conditions using 50, 200, 350, and 500 mM imidizole elution buffers according to the manufacturer’s recom-mendations. Fractions containing the rMAP2 homolog were identified by SDS-polyacrylamide gel electrophoresis and staining with Coomassie blue. The rMAP2 protein contained a C-terminal polyhistidine tag for purification using affinity chromatography. The authenticity of the rMAP2 homolog ofE. chaffeen-siswas evaluated by Western immunoblot and slot blot analysis using a horse-radish peroxidase (HRP)-conjugated antihistidine antibody (Invitrogen Corpo-ration) and knownE. chaffeensisIFA-positive human serum. This serum sample was collected from an individual with a history of tick exposure, clinical and laboratory finding consistent with HME, and acute- and convalescent-phaseE. chaffeensis, IFA titers of 1:160 and 1:320, respectively.
SDS-polyacrylamide gel electrophoresis.The rMAP2 protein concentrations were determined by the Coomassie blue G dye-binding assay as previously described (15). The proteins were dissolved in a 3⫻sample buffer containing 0.1 M Tris (pH 6.8), 5% (wt/vol) SDS, 50% glycerol, and 0.00125% bromophenol blue, either with or without 7.5%-mercaptoethanol. Samples were either heat denatured at 100°C for 3 min prior to electrophoresis or were electrophoresed without heat denaturation on SDS–10% (wt/vol) polyacrylamide gels. Proteins were electrophoretically transferred to nitrocellulose membranes (Hybond ECL; Amersham International plc, Little Chalfont, Buckinghamshire, England) and fixed in 25 mM Tris–191.8 mM glycine–20% methanol as described previously (1).
Antibodies and antisera. HRP-labeled antihistidine antibody (Anti-His(C-term)-HRP; Invitrogen Corporation) was used as a positive control for the rMAP2 homolog on immunoblot assays. Sixty serum samples from the Wads-worth Center, New York State Department of Health, Albany, N.Y., and the University of Florida College of Medicine, Gainesville, Fla., were evaluated for antibodies to the rMAP2 homolog ofE. chaffeensis. Investigators were blinded as to the immune reactivities of the samples. Twenty of the serum samples were previously demonstrated to contain antibodies reactive withE. chaffeensis (Ar-kansas strain) by IFA testing, and 40 samples were IFA negative. The IFA-positive samples were obtained from patients with presenting clinical signs con-sistent with human ehrlichioses. Ten of these samples had IFA titers of 1:160, and the other 10 had titers of 1:320 or greater. Of the 40 IFA-negative samples, 20 were from clinically ill patients, none of whom were diagnosed with HME, and 20 samples were from clinically normal individuals during well-patient visits.
Western immunoblot and slot blot analysis.Nitrocellulose membranes con-taining electrophoretically transferred proteins or proteins directly blotted using a SlotBlot (Hoefer Scientific Instruments, San Francisco, Calif.) were blocked for 1 h with 5% (wt/vol) skim milk in 1⫻phosphate-buffered saline (PBS) with 0.25% Tween 20 and washed with 1% (wt/vol) milk in 1⫻PBS with 0.25% Tween 20 as described previously (1). The membranes were probed with either HRP-labeled antihistidine antibodies at a dilution of 1/15,000 orE. chaffeensis IFA-positive immune sera at a dilution of 1/300. As a negative control, noninfected human serum was used at a dilution of 1/300. Membranes were then washed with 1% (wt/vol) milk in 1⫻PBS as described previously (1) and reacted with a secondary antibody, HRP-conjugated goat anti-human immunoglobulin G (whole molecule; Sigma Chemical Co., St. Louis, Mo.), at a dilution of 1/40,000. Membranes were processed for enhanced chemiluminescence with detection reagents containing luminol (SuperSignal Substrate; Pierce, Rockford, Ill.) as a substrate and were exposed to X-ray film (Hyperfilm-MP; Amersham Interna-tional plc) to visualize the bound antibody.
Indirect ELISA.Polystyrene microtiter plates (Maxi Sorp; Nunc, Roskilde, Denmark) were coated with 100l (per well) of purified rMAP2 homolog ofE. chaffeensis(2g/ml) in 0.05 M carbonate-bicarbonate buffer (pH 9.6) (Sigma Chemical Co.) and incubated overnight at 4°C. The wells were then washed four times with wash buffer containing 1⫻PBS and 0.5% (vol/vol) Tween 20 and blocked for 60 min at room temperature with 1% (wt/vol) bovine serum albumin (BSA) in 1⫻PBS. The plates were washed four times as described above and incubated for 60 min at room temperature with test sera at 1:100, 1:300, 1:1,000, 1:3,000, and 1:10,000 dilutions in 1.0% (wt/vol) BSA in 1⫻PBS (100l). The wells were again washed four times and incubated at room temperature for 60 min in the presence of alkaline phosphatase-conjugated goat anti-human immu-noglobulin G (whole molecule; Sigma Chemical Co.) at a dilution of 1:5,000 in 1% (wt/vol) BSA in 1⫻PBS. The wells were again washed four times, and the substrate,p-nitrophenylphosphate (1 mg/ml) in 0.05 M carbonate-bicarbonate buffer (pH 9.6) (Sigma Chemical Co.), was added at 100l per well and incu-bated for 60 min at room temperature. Absorbance was measured at 405 nm using a Rainbow plate reader (Tecan U.S. Inc., Durham, N.C.). Serum samples from four clinically healthy individuals were used to establish the cutoff values for determining if a test sample was positive or negative. Negative controls were used each time an ELISA was performed. A test sample was considered positive if the absorbance reading was at least double that of the negative controls.
RESULTS
Analysis of the predicted amino acid sequence of the rMAP2
homolog indicated that it differed from the previously reported
amino acid sequence of the
E. chaffeensis
MAP2 at two
posi-tions. Position 17 in the native
E. chaffeensis
MAP2 contains
the polar amino acid serine, while the rMAP2 has a nonpolar
amino acid, tryptophan, at this position. A conserved amino
acid substitution also occurred at position 195 of the reported
sequence, where an aspartic acid was replaced with asparagine
in the recombinant protein. Both the native and recombinant
map
2 sequences were derived from the Arkansas strain of
E.
chaffeensis
. We have not investigated whether these differences
were the result of PCR amplification errors or whether they
represent true polymorphism in regions of the gene.
Coomassie blue-stained SDS-polyacrylamide gels were used
to evaluate each of five fractions of 50, 200, 350, and 500 mM
imidizole elution buffer for the presence of the rMAP2
ho-molog. A single protein with a molecular mass of
approxi-mately 26 kDa was identified in fractions 2 through 5 of the 350
mM imidizole buffer and fractions 1 and 2 of the 500 mM
imidizole buffer (Fig. 1). This protein contains a fusion
pep-tide, increasing the size of the recombinant seen on Coomassie
blue-stained gels. Based on the amino acid sequence, the
cal-culated mass of the rMAP2 is approximately 21 kDa.
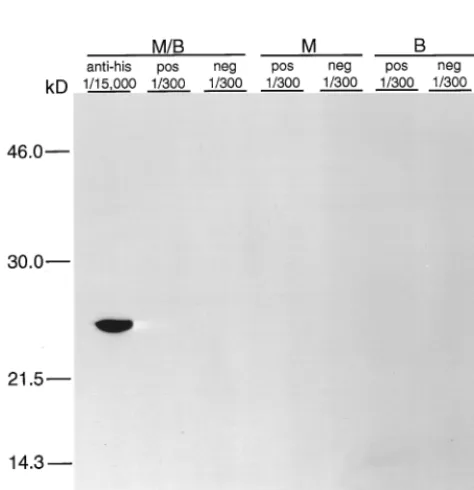
Western immunoblot analysis was done to evaluate
reactiv-ity of the rMAP2 homolog with immune serum from a patient
who was seropositive for infection with
E. chaffeensis
. In these
experiments, purified proteins were run on
SDS-polyacryl-amide gel electrophoresis under denaturing conditions.
Nor-mal human serum was used as a negative control on
immuno-blots, and HRP-labeled antihistidine antibody was used as a
positive control for detection of the recombinant protein. The
26-kDa recombinant protein was clearly identified using the
antihistidine antibody (Fig. 2). However, known positive
on May 15, 2020 by guest
http://jcm.asm.org/
mune serum failed to react with the recombinant antigen in
Western immunoblots.
In order to determine if the denaturing conditions were
affecting the protein reactivity, slot blotting was performed
using the rMAP2 homolog in sample buffers with and without

-mercaptoethanol, with and without boiling, and with and
without fixing the membrane in 20% methanol. rMAP2 was
recognized by antihistidine antibody even when it was
dena-tured by boiling in

-mercaptoethanol (Fig. 3). However,
im-mune serum recognized rMAP2 only when it was prepared in
sample buffer without

-mercaptoethanol and without boiling
prior to blotting on the membrane. rMAP2 that was either heat
denatured or reduced using

-mercaptoethanol failed to react
with immune serum. Fixation of the protein to the membrane
using 20% methanol did not abolish the reactivity of the
pro-tein with the immune serum but enhanced the signal visualized
on the immunoblot (Fig. 3).
[image:3.612.316.547.69.305.2]ELISAs were performed on 60 serum samples which were
previously evaluated for antibodies against
E. chaffeensis
by
IFA testing. All samples negative by IFA were also found to be
negative for antibodies against the rMAP2 homolog using the
ELISA (data not shown). A comparison of rMAP2 ELISA
results with
E. chaffeensis
IFA-positive samples is presented in
Table 1. Only 1 of 20 IFA-positive samples tested negative in
the rMAP2 ELISA. Therefore, there was 100% agreement
using negative samples and 95% agreement using
IFA-positive samples, resulting in a 97.5% overall agreement
be-tween the two assays. In samples that were positive by both
assays, the rMAP2 ELISA was usually able to detect antibodies
at a higher dilution of serum (Table 1).
DISCUSSION
We have successfully cloned the
map
2 gene from
E.
chaf-feensis
and have purified rMAP2 translated from the open
reading frame encoding the predicted mature protein. This
recombinant is unique and to our knowledge is thus far the
only
E. chaffeensis
recombinant antigen that has been shown to
work in an ELISA format. The predicted amino acid sequence
of this protein has significant homology with the sequences of
FIG. 1. Coomassie blue-stained gel of eluted fractions containing the rMAP2 homolog ofE. chaffeensis. Lanes 2, 3, 4, and 5, respective fractions eluted with 350 mM imidizole elution buffer; lanes 1 and 2, respective fractions eluted with 500 mM imidizole buffer. Molecular size (M.W.) standards are in the unmarked lane on the left.
[image:3.612.94.253.72.237.2]FIG. 2. Immunoblot of the rMAP2 homolog ofE. chaffeensiseither reduced in sample buffer containing-mercaptoethanol and heat denatured by boiling (M/B), reduced in sample buffer containing-mercaptoethanol without boiling (M), or boiled in sample buffer without-mercaptoethanol (B) and reacted with antihistidine antibodies (anti-his), E. chaffeensisIFA-positive immune serum (pos), orE. chaffeensisIFA-negative serum (neg). The fractions (1/300 and 1/15,000) indicate the dilutions of antibody or serum used. Molecular size stan-dards are given on the left.
FIG. 3. Slot blot of rMAP2 ofE. chaffeensiseither reduced in sample buffer containing-mercaptoethanol and heat denatured by boiling (M/B), reduced in sample buffer containing-mercaptoethanol without boiling (M), boiled in sam-ple buffer without-mercaptoethanol (B), in PBS without boiling or -mercap-toethanol (NT), or in PBS without boiling or mercap-mercap-toethanol and with fixing in 20% methanol (Meth). Recombinant antigens were reacted with antibody con-trols (antihistidine antibodies [anti-his] orE. chaffeensisIFA-negative serum [neg]) or immune sera (E. chaffeensisIFA-positive immune serum [Pos]). The fractions (1/300 and 1/15,000) indicate the dilutions of antibody or serum used.
on May 15, 2020 by guest
http://jcm.asm.org/
[image:3.612.55.292.411.656.2]the 21-kDa MAP2 protein of
C. ruminantium
and the 19-kDa
MSP5 protein of
A. marginale
(2), and the protein is distinctly
different in size and sequence from the 28-kDa outer
mem-brane protein of
E. chaffeensis
, the p28 antigen. Genes
encod-ing both the MAP2 of
C. ruminantium
and the MSP5 of
A.
marginale
were found to be well conserved between various
isolates of organisms within the respective species, and
recom-binant proteins developed from these genes have been used as
diagnostic test antigens to identify infected animals (2, 9, 11,
17). Importantly, development of a competitive ELISA using a
monoclonal antibody against MSP5 resulted in an assay that
was capable of detecting both acutely and persistently infected
cattle. This MSP5 ELISA did not cross-react with sera from
cattle infected with closely related organisms (9, 17).
There-fore, although these antigens are well conserved within a
spe-cies, areas of sequence variability between species may provide
a way to enhance test specificity.
Interestingly, we were not able to demonstrate reactivity of
immune sera with rMAP2 of
E. chaffeensis
using Western
im-munoblot assays. Using slot blot assays, we further confirmed
that heat denaturation or reduction of disulfide bonds by

-mercaptoethanol destroys important structural epitopes of
the antigen needed for antibody recognition. Fixing the
mem-brane-bound protein in 20% methanol did not inhibit reactivity
with immune sera but resulted in enhancement of the signal,
probably associated with increased amounts of protein being
fixed to the nitrocellulose membrane. Similarly,
conformation-ally dependent epitopes were also identified on MSP5 of
A.
marginale
(12). In that study, reduction of disulfide bonds using
dithiothreitol, followed by treatment with iodoacetamide to
modify sulfhydryl groups, completely abolished monoclonal
antibody binding to MSP5 in dot blot assays. In addition, Chen
et al. identified 20 immunogenic proteins of
E. chaffeensis
using
rabbit and human
E. chaffeensis
antisera in Western
immuno-blot assays (4) and showed that one protein of approximately
22 kDa was heat labile, while all of the other identified
immu-nogens were heat stable. Based on its size and heat-labile
nature, the 22-kDa protein could be identical to the MAP2
protein described here. In the same study (4), the 22-kDa
protein of
E. chaffeensis
cross-reacted only with anti-
E. canis
immune serum, whereas the other major immunogenic
anti-gens of 44, 55, and 66 kDa all cross-reacted with antisera
against
E. canis
,
Ehrlichia ewingii
,
Ehrlichia risticii
, and
Ehrli-chia sennetsu
. This gives further indication that the rMAP2
protein could be used as a sensitive and specific antigen in a
diagnostic test.
ELISAs performed using the rMAP2 homolog of
E.
chaffeensis
were in 100% agreement with the IFA test results
when IFA-negative serum samples were evaluated. There was
95% agreement between these two assays using
E. chaffeensis
IFA-positive samples. Differences in reactivity between the
ELISA and IFA could be explained by the use of a single
recombinant protein versus the cultured, whole organisms
used in the IFA. This could result in false-negative results
using the rMAP2 ELISA. Alternately, IFA testing may result
in false-positive results due to the difficulty in distinguishing
true-positive reactions from nonspecific antibody binding
and/or to an increased likelihood of cross-reactive antigens. In
samples that were positive by both assays, the rMAP2 ELISA
was usually able to detect antibodies at a higher dilution of
serum. This may indicate enhanced sensitivity of the ELISA
over the
E. chaffeensis
IFA and in the future may provide for a
more sensitive assay.
These data, combined with information gathered from
pre-vious work involving the MSP5 of
A. marginale
and MAP2 of
C. ruminantium
, provide evidence that the MAP2 homolog of
E. chaffeensis
is a potential candidate for the serologic
diagno-sis of human ehrlichioses. Because of the antigenic similarities
between organisms within the
Ehrlichia
species (4), and
possi-bly closely related genera (8), it may be necessary to design a
competitive ELISA based on the inhibition of monoclonal
antibodies, similar to the assay developed using MSP5 of
A.
marginale
. An assay such as this, although presently not
avail-able, would provide a convenient and cost-effective way to
analyze large numbers of samples for clinical use and
epide-miologic studies. Further investigation is needed to better
characterize the MAP2 antigen of
E. chaffeensis
with regard to
the sensitivity and specificity of this protein as a diagnostic
antigen.
ACKNOWLEDGMENTS
This study was supported by a grant from the University of Florida,
Division of Sponsored Research, project UPN 98062369. This work
was supported in part by a grant from the Centers for Disease Control
and Prevention (815-3478A) of the Public Health Service.
REFERENCES
1.Alleman, A. R., and A. F. Barbet.1996. Evaluation ofAnaplasma marginale major surface protein 3 (MSP3) as a diagnostic test antigen. J. Clin. Micro-biol.34:270–276.
2.Bowie, M. V., G. R. Reddy, S. M. Semu, S. M. Mahan, and A. F. Barbet.1999. Potential value of major antigenic protein 2 for serological diagnosis of heartwater and related ehrlichial infections. Clin. Diagn. Lab. Immunol. 6:209–215.
3.Brouqui, P., C. Lecam, J. Olson, and D. Raoult.1994. Serologic diagnosis of human monocytic ehrlichiosis by immunoblot analysis. Clin. Diagn. Lab. Immunol.1:645–649.
4.Chen, S. M., J. S. Dumler, H. M. Feng, and D. H. Walker.1994. Identifica-tion of the antigenic constituents ofEhrlichia chaffeensis. Am. J. Trop. Med. Hyg.50:52–58.
5.Chen, S. M., V. L. Popov, H. M. Feng, and D. H. Walker.1996. Analysis and ultrastructural localization ofEhrlichia chaffeensisproteins with monoclonal antibodies. Am. J. Trop. Med. Hyg.54:405–412.
6.Fishbein, D. B., J. E. Dawson, and L. E. Robinson.1994. Human ehrlichiosis in the United States, 1985 to 1990. Ann. Intern. Med.120:736–743. 7.Ijdo, J. W., W. U. Caiyun, L. A. Magnarelli, and E. Fikrig.1999.
Serodiag-nosis of human granulocytic ehrlichiosis by a recombinant HGE-44-based enzyme-linked immunosorbent assay. J. Clin. Microbiol.37:3540–3544. 8.Kelly, P. J., L. A. Matthewman, S. M. Mahan, and T. Peter.1994. Serologic
evidence for antigenic relationships betweenEhrlichia canisandCowdria ruminantium. Res. Vet. Sci.56:176–174.
9.Knowles, D. P., S. Torioni de Echaide, G. H. Palmer, T. C. McQuire, D. Stiller, and T. McElwain.1996. Antibody against anAnaplasma marginale MSP5 epitope common to tick and erythrocyte stages identifies persistently infected cattle. J. Clin. Microbiol.34:2225–2230.
[image:4.612.52.293.91.211.2]10. Magnarelli, L. A., K. C. Stafford, T. N. Mather, M. T. Yeh, K. D. Horn, and J. S. Dumler.1995. Hemocytic rickettsia-like organisms in ticks: serologic reactivity with antisera toEhrlichiaeand detection of DNA of agent of
TABLE 1. Comparison of rMAP2 ELISA results with 20
E. chaffeensis
IFA-positive samples
No. samples rMAP2 ELISAtitera E. chaffeensisIFA titer
1
Negative
1:320
1
1:100
1:160
2
1:100
1:320
1
1:100
⬎1:640
4
1:300
1:160
1
1:1,000
1:160
1
1:1,000
⬎1:640
4
1:3,000
1:160
2
1:3,000
1:320
3
1:3,000
⬎1:640
aA sample was considered positive in the rMAP2 ELISA when the absorbance
reading of the test serum at a specific dilution was two or more times the absorbance reading of identical dilutions of standard negative control sera. A negative ELISA result indicates a titer of⬍1:100.
on May 15, 2020 by guest
http://jcm.asm.org/
human granulocytic ehrlichiosis by PCR. J. Clin. Microbiol.33:2710–2714. 11. Mahan, S. M., T. C. McGuire, S. M. Semu, M. V. Bowie, F. Jongejan, F. R.
Rurangirwa, and A. F. Barbet.1994. Molecular cloning of a gene encoding the immunogenic 21 kDa protein ofCowdria ruminantium. Microbiology 140:2135–2142.
12. Munodzana, D., T. F. McElwain, D. P. Knowles, and G. H. Palmer.1998. Conformational dependence ofAnaplasma marginalemajor surface protein 5 surface-exposed B-cell epitopes. Infect. Immun.66:2619–2624. 13. Nyindo, M., I. Kakoma, and R. Hansen.1991. Antigenic analysis of four
species of the genusEhrlichiaby use of protein immunoblots. J. Am. Vet. Res.52:1225–1230.
14. Ohasi, N., A. Unver, N. Zhi, and Y. Rikihisa.1998. Cloning and character-ization of multigenes encoding the immunodominant 30-kilodalton major outer membrane proteins ofEhrlichia canisand application of the recombi-nant protein for serodiagnosis. J. Clin. Microbiol.36:2671–2680. 15. Read, S. M., and D. H. Northcote.1981. Minimization of variation in the
response to different proteins of the Coomassie blue G dye-binding assay for protein. Anal. Biochem.116:53–64.
16. Unver, A., Y. Rikihisa, N. Ohashi, L. C. Cullman, R. Buller, and G. A. Storch.1999. Western and dot blotting analyses ofEhrlichia chaffeensis
indirect fluorescent-antibody assay-positive and -negative human sera by using native and recombinantE. chaffeensisandE. canisantigens. J. Clin. Microbiol.37:3888–3895.
17. Visser, E. S., T. C. McGuire, G. H. Palmer, W. C. Davis, V. Shkap, E. Pipano, and D. P. Knowles.1992. TheAnaplasma marginale msp5 gene encodes a 19-kilodalton protein conserved in all recognizedAnaplasmaspecies. Infect. Immun.60:5139–5144.
18. Walker, D. H. 2000. Diagnosing human ehrlichioses: current status and recommendations. ASM News66:287–290.
19. Wong, S. J., G. S. Brady, and J. S. Dumler.1997. Serological responses to Ehrlichia equi,Ehrlichia chaffeensis, andBorrelia burgdorferiin patients from New York State. J. Clin. Microbiol.35:2198–2205.
20. Yu, X., P. A. Crocquet-Valdes, L. C. Cullman, and D. H. Walker.1996. The recombinant 120-kilodalton protein ofEhrlichia chaffeensis, a potential di-agnostic tool. J. Clin. Microbiol.34:2853–2855.
21. Yu, X., P. A. Crocquet-Valdes, L. C. Cullman, V. L. Popov, and D. H. Walker. 1999. Comparison ofEhrlichia chaffeensisrecombinant proteins for serologic diagnosis of human monocytic ehrlichiosis. J. Clin. Microbiol.37:2568–2575. 22. Zhou, C., Y. Yang, and A. Y. Jong.1990. MiniPrep in ten minutes.
BioTech-niques8:172–173.