HELGOLANDER MEERESUNTERSUCHUNGEN
Hetgol~inder Meeresunters. 49, 223-236 (1995)
J
Correlation of viable cell counts, metabolic activity of
s u l p h u r - o x i d i z i n g bacteria and chemical parameters of
marine s e d i m e n t s
J. F. Imhoff, A. Schneider & L. Podgorsek
Institut ffir Meereskunde, Abteilung Marine Mikrobiologie;
Dfisternbrooker W e g 20, D-24105 Kiel, Germany
ABSTRACT: Viable counts of aerobic a n d a n a e r o b i c chemotrophic sulphur-oxidizers as well as phototrophic s u l p h u r bacteria were d e t e r m i n e d in s e d i m e n t samples t a k e n from two different areas along the Baltic Sea shore which were k n o w n to regularly develop sulphidic conditions. Depth profiles of bacterial cell counts were correlated with concentration profiles of chloride, sulphate, sulphide, nitrate a n d p h o s p h a t e in the pore water of these sediments a n d with potential activities of nitrate reduction, thiosulphate transformation and s u l p h a t e formation. The data r e v e a l e d a complex multilayered structure within the sediments. Sulphide was released into the water from s e d i m e n t s of b o t h sampling areas, but it was found that light a n d the availability of oxygen significantly r e d u c e d this amount. In the highly reduced s e d i m e n t at Hiddensee, the highest n u m b e r s of phototrophic a n d chemotrophic sulphur-oxidizers were found n e a r the s e d i m e n t surface. Therefore, it was c o n c l u d e d that the c o m b i n e d action of both groups of bacteria most efficiently oxidizes r e d u c e d sulphur compounds in the top layers of the sediments. Nitrate may replace oxygen as final electron acceptor a n d will support oxidation of sulphide, in particular w h e n oxygen and light are limiting.
I N T R O D U C T I O N
T h e i n c r e a s i n g e u t r o p h i c a t i o n of m a r i n e c o a s t a l a r e a s , i n p a r t i c u l a r its c o r r e l a t i o n w i t h i n c r e a s e s of s u l p h i d i c l o c a t i o n s i n m a r i n e b a s i n s a n d c o a s t a l a r e a s , h a s r e n e w e d i n t e r e s t i n r e a c t i o n s of t h e m i c r o b i a l s u l p h u r c y c l e a n d its i m p o r t a n c e for t h e s e e n v i r o n - m e n t s . A c o n s i d e r a b l e p a r t of t h e o r g a n i c m a t t e r i n m a r i n e s e d i m e n t s is m i n e r a l i z e d b y s u l p h a t e - r e d u c i n g b a c t e r i a ( J 6 r g e n s e n , 1982). I n g e n e r a l , t h e s e b a c t e r i a h a v e a s e l e c t i v e a d v a n t a g e o v e r m e t h a n o g e n i c b a c t e r i a i n t h e m a r i n e e n v i r o n m e n t . T h e m e t h a n o g e n s c a n c o m p e t e o n l y w h e n s u l p h a t e is l i m i t i n g , w h e n o r g a n i c s u h s t r a t e s a r e s u p p l i e d i n e x c e s s or w h e n a l t e r n a t i v e n o n - c o m p e t i t i v e s u b s t r a t e s , l i k e m e t h y l a m i n e , a r e a v a i l a b l e ( W i d d e l , 1988). S u l p h a t e r e d u c t i o n l e a d s to t h e p r o d u c t i o n of s u l p h i d e , w h i c h is t o x i c to b a c t e r i a , a n i m a l s a n d p l a n t s . S u l p h i d e is r e l e a s e d f r o m s e d i m e n t s i n t o t h e s u p e r n a t a n t w a t e r a n d a l s o i n t o t h e a t m o s p h e r e f r o m c o a s t a l a r e a s l i k e t h e W a d d e n S e a a n d t h e s h a l l o w a r e a s of t h e B a l t i c S e a s h o r e .
M o s t of t h e s u l p h i d e p r o d u c e d i n m a r i n e s e d i m e n t s is s u p p o s e d to b e o x i d i z e d b y b a c t e r i a l a c t i v i t y r a t h e r t h a n c h e m i c a l l y . I m p o r t a n t i n t e r m e d i a t e s i n t h e o x i d a t i o n of s u l p h i d e a r e e l e m e n t a l s u l p h u r a n d t h i o s u l p h a t e ( J S r g e n s e n & B a k , 1991). T h e l a t t e r a l s o i s o n e of t h e m o s t c o n v e n i e n t s o u r c e s of r e d u c e d s u l p h u r for t h e c u l t i v a t i o n of b a c t e r i a
224 J. F. Imhoff, A. S c h n e i d e r & L. P o d g o r s e k
that oxidize r e d u c e d s u l p h u r c o m p o u n d s . A l t h o u g h t h r e e m a j o r p h y s i o l o g i c a l groups of b a c t e r i a with such activities are k n o w n - the a e r o b i c s u l p h u r - o x i d i z e r s , the nitrate- r e d u c i n g sulphur-oxidizers, a n d t h e p h o t o t r o p h i c s u l p h u r b a c t e r i a - only a s m a l l n u m b e r of species of t h e s e b a c t e r i a h a v e b e e n i d e n t i f i e d from the m a r i n e e n v i r o n m e n t . Informa- tion on their distribution a n d f r e q u e n c y in m a r i n e habitats is rare. T h e p r e s e n t study was initiated in o r d e r to correlate v i a b l e n u m b e r s of b a c t e r i a p a r t i c i p a t i n g in the b a c t e r i a l sulphur cycle w i t h c h e m i c a l p a r a m e t e r s from t h e s e d i m e n t s a n d to l o c a t e b a c t e r i a a n d their activities within the s e d i m e n t layers.
M A T E R I A L A N D M E T H O D S
T h e s a m p l i n g s i t e s : T w o s a m p l i n g sites that w e r e k n o w n to d e v e l o p r e g u - larly sulphidic conditions w e r e s e l e c t e d for our analyses. T h e first site, close to the Island of H i d d e n s e e , r e p r e s e n t s an o p e n r e e d b e l t w i t h highly r e d u c e d s e d i m e n t , w h i c h has a l r e a d y b e e n d e s c r i b e d by S u c k o w (1966). T h e s a m p l i n g o c c u r r e d during a t i m e of highly fluctuating w a t e r levels (from 0 - 2 0 cm) a b o v e the s e d i m e n t . T h e other site is l o c a t e d n e a r F6hrdorf, close to Poel, a n d m a y b e d e s c r i b e d as a small, n e a r l y closed r e e d bight. During the s a m p l i n g period, it a p p e a r e d o x i d i z e d at the surface, b u t w a s r e d u c e d at d e e p e r levels. At that t i m e the w e a t h e r w a s c a l m a n d t h e r e w a s a constant w a t e r l e v e l a b o v e the s e d i m e n t of 1-5 cm. T h e structure of the s e d i m e n t n e a r F6hrdorf w a s m o r e h o m o g e n e o u s a n d c o n s i s t e d of a s m a l l e r g r a i n size t h a n that of H i d d e n s e e .
S a m p l i n g o f s e d i m e n t s : T h e s a m p l i n g o c c u r r e d during J u n e 1992. Sedi- m e n t cores w e r e directly cut into discs of 1 to 5 cm thickness, as n o t e d in T a b l e s 1 a n d 2. C o r r e l a t i o n of the different p a r a m e t e r s r e q u i r e d an i m m e d i a t e t r e a t m e n t of the discs a c c o r d i n g to the m e t h o d s listed below. T h e r e f o r e , a n d b e c a u s e of the g r e a t n u m b e r of s i m u l t a n e o u s a n a l y s e s a n d e x p e r i m e n t s , statistical t r e a t m e n t of the s e d i m e n t cores was not possible. H o w e v e r , c o r r e s p o n d i n g disks from 3 - 5 s e d i m e n t cores t a k e n f r o m the s a m e location w e r e c o m b i n e d , carefully m i x e d a n d u s e d for further t r e a t m e n t s .
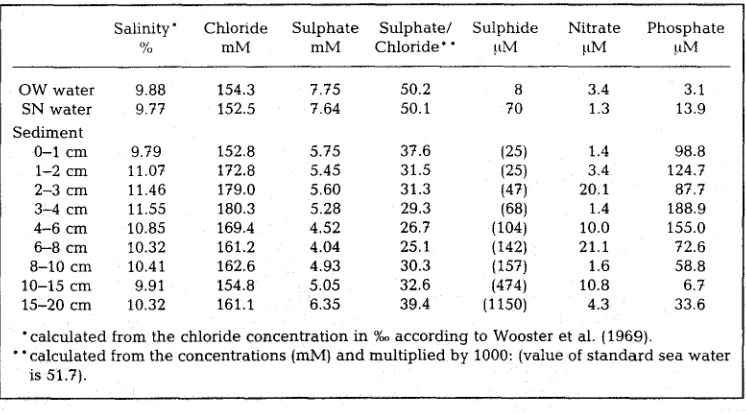
C h e m i c a 1 a n a 1 y s e s : Pore w a t e r from the s e d i m e n t discs was r e c o v e r e d after c e n t r i f u g a t i o n a n d sterilized by filtration (0.45 ~m pore size cellulose a c e t a t e m e m b r a n e filters). This w a t e r was u s e d to d e t e r m i n e c o n c e n t r a t i o n s of chloride, s u l p h a t e , nitrate a n d p h o s p h a t e . S e p a r a t i o n a n d q u a n t i f i c a t i o n of t h e s e anions was a c h i e v e d by ion c h r o m a t o g r a p h y u s i n g a D i o n e x DX 300, e q u i p p e d w i t h conductivity- a n d U V - d e t e c t o r s a n d an a n i o n e x c h a n g e column (Dionex*AS4A). C h e m i c a l a n a l y s e s w e r e m a d e at t h r e e different sensitivity r a n g e s of the c o n d u c t i v i t y detector. T h e salinity w a s c a l c u l a t e d from the c o n c e n t r a t i o n of chloride (salinity = 1.80655 chlorinity) a c c o r d i n g to W o o s t e r et al. (1969).
V i a b l e cells, s u l p h u r - o x i d i z i n g bacteria, c h e m i c a l p a r a m e t e r s 225 a l l o w e d to s t a n d for at least 30 min. After colour d e v e l o p m e n t w a s c o m p l e t e d , the s a m p l e v o l u m e w a s a d j u s t e d to a total of 20 ml a n d the optical density m e a s u r e d at 668 nm a g a i n s t a r e a g e n t blank.
B a c t e r i a 1 a n a 1 y s e s : In order to o v e r c o m e s m a l l - s c a l e h e t e r o g e n e i t y of the sediments, c o r r e s p o n d i n g s e d i m e n t discs of 3 - 5 cores from e a c h s a m p l i n g sites w e r e h o m o g e n o u s l y mixed, and v o l u m e s of 0.5 cm 3 s e d i m e n t w e r e u s e d for dilution series. T h e dilution w a s p e r f o r m e d in 70 % filter-sterilized, n a t u r a l s e a w a t e r (taken from the North S e a n e a r List/Sylt). From t h e s e dilution series, various m e d i a w e r e i n o c u l a t e d . T h e m a t e r i a l was e i t h e r s p r e a d on plates (for a e r o b i c bacteria) or i n o c u l a t e d in d e e p - a g a r (for a n a e r o b i c bacteria).
P h o t o t r o p h i c b a c t e r i a w e r e c u l t i v a t e d in P f e n n i g ' s m e d i u m , s u p p l e m e n t e d with a c e t a t e solution (1 ml/100 ml) a n d salt solution (5 ml/100 ml) a c c o r d i n g to P f e n n i g & Tr~iper (1992). Cell counts w e r e o b t a i n e d from dilution series in agar.
A n e w m e d i u m ("Lith" m e d i u m ) w a s d e s i g n e d for m a r i n e a n a e r o b i c sulphur- oxidizing bacteria, using different e l e c t r o n acceptors. T h e composition of the basic m i x t u r e of the m e d i u m c o n t a i n e d in final a m o u n t s p e r litre: 700 ml of a modified, s u l p h a t e - f r e e s e a w a t e r (MSM), 1.68 g N a H C O 3 , 0.68 g KH2PO4, 0.25 g NH4C1, 1 ml trace e l e m e n t solution SLB (modified SLA with only 1 m g N a 2 S e O 3 - 5 H 2 0 p e r litre) a n d 1 ml v i t a m i n solution VA (Imhoff, 1988). T h e modified, s u l f a t e - f r e e sea w a t e r (MSM) was a u t o c l a v e d s e p a r a t e l y and c o n t a i n e d the f o l l o w i n g a m o u n t s p e r litre: 27.3 g NaC1, 2.7 g MgC12 - 6 H20, 0.75 g KC1, 0.4 g CaC12 . 2 H 2 0 , a n d 0.14 g KBr. T h e p H w a s a d j u s t e d to 7.2. A m i x t u r e of Na2S203 " 5 H20, Na2S . 9 H 2 0 a n d p o l y s u l p h i d e ( p r e p a r e d a c c o r d i n g to F e h e r & Laue, 1956) was a p p l i e d as a source of r e d u c e d s u l p h u r in final c o n c e n t r a t i o n s of 5 mM, 0.5 m M a n d 13 raM, respectively. N a H C O 3 a n d s o d i u m a c e t a t e (5 mM) w e r e a d d e d as the only carbon sources. Either KNO3 (10 mM) or Na2SO4 (10 mM) s e r v e d as e l e c t r o n acceptors.
A e r o b i c b a c t e r i a w e r e c o u n t e d on a g a r p l a t e s u s i n g a m i n e r a l salts m e d i u m ("Thio" medium). T h e basic m e d i u m c o n t a i n e d 700 ml n a t u r a l sea water, 0.4 g NH4C1 a n d 0.5 g KH2PO4. N a H C O 3 (1.0 g/l) and sodium a c e t a t e (5 mM) w e r e a d d e d as c a r b o n sources, a n d s o d i u m t h i o s u l p h a t e (3.2 raM) and Na2 S - 9 H 2 0 (0.5 mM) as sulphur sources and electron donors. T h e p H was a d j u s t e d to 7.2
M e a s u r e m e n t s o f b a c t e r i a l a c t i v i t i e s : T r a n s f o r m a t i o n of thiosulfate was m e a s u r e d u n d e r aerobic conditions in s h a k e n E r l e n m e y e r flasks a n d u n d e r a n a e r o b i c conditions with KNO3 (7 mM) as a d d e d e l e c t r o n acceptor. A n a e r o b i c condi- tions w e r e a c h i e v e d in H u n g a t e t u b e s by r e p e a t e d e v a c u a t i o n a n d flushing w i t h n i t r o g e n gas. Assays w e r e p e r f o r m e d u n d e r n i t r o g e n a n d in the dark. For all activity m e a s u r e - ments, the b a s a l m e d i u m for a n a e r o b i c b a c t e r i a w a s s u p p l e m e n t e d with s o d i u m a c e t a t e (5 raM) a n d Na2S203 9 5 H 2 0 t5 mM). T h e assays w e r e started by the a d d i t i o n of the s e d i m e n t s a m p l e (1 c m 3) to 15 ml of m e d i u m . S u l p h a t e r e d u c t i o n w a s i n h i b i t e d by addition of Na2MoO4 ' 2 H 2 0 (20 mM). S a m p l e s w e r e w i t h d r a w n after 4.5 h of i n c u b a t i o n in the dark at r o o m t e m p e r a t u r e a n d sterilized b y filtration.
226 J. F. I m h o f f , A. S c h n e i d e r & L. P o d g o r s e k R E S U L T S
In o r d e r to e v a l u a t e t h e r o l e of b a c t e r i a i n v o l v e d i n t h e o x i d a t i o n of r e d u c e d s u l p h u r c o m p o u n d s i n m a r i n e s e d i m e n t s , w e a n a l y s e d t h e t o p 20 c m of s e d i m e n t f r o m t w o d i f f e r e n t s a m p l i n g s i t e s a l o n g t h e c o a s t of t h e B a l t i c S e a . A l t h o u g h t h e s e d i m e n t s f r o m b o t h a r e a s s h o w e d a c t i v e s u l p h a t e r e d u c t i o n a n d r e l e a s e of s u l p h i d e i n t o t h e s u p e r n a t a n t w a t e r , r e m a r k a b l e d i f f e r e n c e s w e r e f o u n d i n t h e d e p t h p r o f i l e s of t h e i r c h e m i c a l c o m p o - s i t i o n a n d i n t h e d i s t r i b u t i o n of b a c t e r i a l a c t i v i t i e s . T h e s a m p l i n g s i t e a t H i d d e n s e e o b v i o u s l y w a s c h a r a c t e r i z e d b y t r e m e n d o u s a c t i v i t y of s u l p h a t e - r e d u c i n g b a c t e r i a a n d r e l e a s e of s u l p h i d e i n t o t h e w a t e r ; w h e r e a s s u l p h a t e r e d u c t i o n at t h e s a m p l i n g s i t e n e a r F ~ h r d o r f a p p e a r e d to b e q u i t e low. N e v e r t h e l e s s , s u l p h i d e w a s m e a s u r a b l e i n t h e t h i n w a t e r l a y e r c o v e r i n g t h i s s e d i m e n t .
T h e
s e d i m e n t
a tH i d d e n s e e
T h e c h e m i c a l c o m p o s i t i o n of t h i s s e d i m e n t is s h o w n i n T a b l e 1. M i n o r s a l i n i t y c h a n g e s , b e t w e e n c a 9 a n d 12 %o, o c c u r r e d t h r o u g h o u t t h e s e d i m e n t . M o s t o b v i o u s l y , t h e s u l p h a t e c o n t e n t of t h e s e d i m e n t w a s s i g n i f i c a n t l y r e d u c e d c o m p a r e d to t h e s u p e r n a t a n t w a t e r . P a r t i c u l a r l y i n d e p t h s b e t w e e n 3 - 8 cm, t h e m i n i m u m v a l u e s of t h e s u l p h a t e / c h l o r i d e r a t i o of 25.1 to 29.3 ( c a l c u l a t e d a s i n d i c a t e d i n T a b l e 1) p o i n t t o a z o n e of c o n t i n u o u s s u l p h a t e d e p l e t i o n b y s u l p h a t e r e d u c t i o n . T h i s z o n e w a s a c c o m p a n i e d b y a m a x i m u m of t h e p h o s p h a t e c o n c e n t r a t i o n . T h e n i t r a t e p o o l w a s a b o u t 2 - 3 o r d e r s of m a g n i t u d e l o w e r t h a n t h e s u l p h a t e p o o l , b u t s h o w e d s i g n i f i c a n t c h a n g e s t h r o u g h o u t t h e s e d i m e n t c o l u m n , w h i c h i n d i c a t e s a c t i v e t u r n o v e r . A c c o r d i n g to t h e c o n c e n t r a t i o n g r a -
Table 1. Chemical p a r a m e t e r s of a s e d i m e n t from Hiddensee. Values in brackets w e r e t a k e n from a dialysis pore w a t e r sampler. Parallel m e a s u r e m e n t s by V61kel & Schmidt (pets. comm.) r e v e a l e d c o m p a r a b l e concentrations of sulphide in the sediment. OW = surface water: SN = w a t e r close to
the s e d i m e n t surface
Salinity" Chloride S u l p h a t e S u l p h a t e / Sulphide Nitrate P h o s p h a t e
% mM m M Chloride * * !tM [tM ttM
OW w a t e r 9.88 154.3 7.75 50.2 8 3.4 3.1
SN w a t e r 9.77 152.5 7.64 50.1 70 1.3 13.9
S e d i m e n t
0-1 cm 9.79 152.8 5.75 37.6 [25) 1.4 98.8
1-2 cm 11.07 172.8 5.45 31.5 [25~ 3.4 124.7
2-3 cm 11.46 179.0 5 60 31.3 (47) 20.1 87.7
3-4 cm 11.55 180.3 5.28 29.3 (68) 1.4 188.9
4 - 6 cm 10.85 169.4 4.52 26.7 (104b 10.0 155.0
6-8 cm 10.32 161.2 4.04 25.1 (142) 21.1 72.6
8-10 crn 10.41 162.6 4.93 30.3 (157) 1.6 58.8
10-15 cm 9.91 154.8 5.05 32.6 [4741 10.8 6.7
15-20 cm 10.32 161.1 6.35 39.4 (1150) 4.3 33.6
9 calculated from the chloride c o n c e n t r a t i o n in %o according to Wooster et al. (1969).
V i a b l e c e l l s , s u l p h u r - o x i d i z i n g b a c t e r i a , c h e m i c a l p a r a m e t e r s 2 2 7
sediment
depth (cm)
- 7
2
4
6
8 ~
I
12
14
16
1 8 -
20
----r--r
1 0 4
i i
h
i
~ ,
aerobic SOB
--[]-- anaerobic SOB
PSB
105
~ - - [ ' I
106
107
108
i
I I l l l i
viable counts * cm "3 sediment
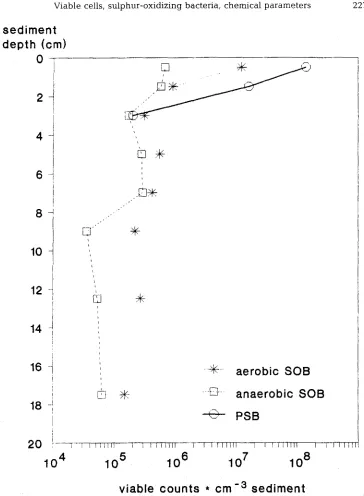
Fig. 1. V i a b l e c o u n t s of s u l p h u r - o x i d i z i n g b a c t e r i a at H i d d e n s e e . SOB = S u l p h u r - o x i d i z i n g b a c t e r i a , PSB = P u r p l e s u l p h u r b a c t e r i a : V i a b l e c o u n t s of a n a e r o b i c SOB w e r e d e t e r m i n e d u n d e r a n o x i c
228
sediment
depth
(cm)
0
J. F. I m h o f f , A. S c h n e i d e r & L. P o d g o r s e k
2
4
6
8
10
12
14
16
18
20
_
... : ...
9
[]
2~
S o 4
formation
S 2 0 32- consumption
I I I I
0
2
4
6
8
10
transformation rates
V i a b l e cells, s u l p h u r - o x i d i z i n g b a c t e r i a , c h e m i c a l p a r a m e t e r s 229
sediment
depth (cm)
0
I
2 -
4~
~
8 ~
lo I
I
J
12
!
J
14
I
)
P
P
2o F
@ ~ i i : : "
9
[]
S 0 4 2 - formation
---~--S2032-consumption
-/~- NO 3- reduction
I I [ I I
0
2
4
6
8
10
transformation rates
230 J. F. Imhoff, A. S c h n e i d e r & L. Podgorsek
dients d u r i n g the s a m p l i n g time, it c a n be c o n c l u d e d that the s e d i m e n t is a p p a r e n t l y a source of p h o s p h a t e a n d p r o b a b l y a sink for nitrate. Sulphide was r e l e a s e d from this s e d i m e n t i n significant amounts. Low c o n c e n t r a t i o n s of sulphide could b e d e t e c t e d e v e n in the oxic surface water of this site (Table 1, OW). Laboratory s t u d i e s with n a t u r a l s e d i m e n t cores r e v e a l e d that u n d e r conditions of b l o c k e d o x y g e n transfer a n d in the dark, the c o n c e n t r a t i o n of s u l p h i d e in t h e s u p e r n a t a n t water i n c r e a s e d c o n s i d e r a b l y w i t h i n 12 h. T h e su!phide c o n c e n t r a t i o n a m o u n t e d to 325 uM directly at the s e d i m e n t surface, 220 ~M at 3 cm a b o v e the s e d i m e n t a n d 200 ,ttM at 6 cm. Using t h e s e data, a n e x c h a n g e rate of sulphide b e t w e e n s e d i m e n t a n d w a t e r of 14.4 mmol m -2 s e d i m e n t d -1 was calculated. Thus o x y g e n availability is e x p e c t e d to b e a limiting factor for s u l p h i d e oxidation in the top layers of this s e d i m e n t , at least d u r i n g dark periods.
Aerobic a n d anaerobic, n i t r a t e - r e d u c i n g sulphur-oxidizers as well as phototrophic s u l p h u r bacteria h a d m a x i m a of v i a b l e cell n u m b e r s in the top layer of this s e d i m e n t (Fig. 1). N u m b e r s of phototrophic p u r p l e s u l p h u r b a c t e r i a e x c e e d e d those of all others, e v e n those of aerobic sulphur-oxidizers. B e c a u s e of their l i g h t - d e p e n d e n c e , they formed a p i n k to r e d - c o l o u r e d layer in the top millimetres of the sediment, just u n d e r n e a t h a very thin surface layer. Viable counts d e c r e a s e d b y o n e order of m a g n i t u d e i n the s e c o n d (1-2 cm) a n d by two more orders in the third s e d i m e n t disc (2-4 cm). V i a b l e cells of phototrophic g r e e n s u l p h u r bacteria were several orders of m a g n i t u d e lower, ca 2 - 4 . 10 4
cells cm -3 in the top layers of the s e d i m e n t .
Also cell n u m b e r s of c h e m o t r o p h i c sulphur-oxidizers u s i n g o x y g e n or nitrate as electron acceptors were h i g h e s t in the top disc of the sediment. They d r o p p e d b y a b o u t o n e order of m a g n i t u d e b e l o w 2 cm, b u t r e m a i n e d relatively high over the rest of the s e d i m e n t clown to 10-15 cm d e p t h (Fig. 1).
T h e capability to oxidize t h i o s u l p h a t e u n d e r aerobic conditions w a s high in this s e d i m e n t (Fig. 2a). A l t h o u g h s u l p h a t e was the major oxidation product, t r a n s f o r m a t i o n rates of t h i o s u l p h a t e I1.4-6.5 ~tmol h -1 c m -3) w e r e not q u a n t i t a t i v e l y c o r r e l a t e d with the formation of s u l p h a t e (1.1-5.2 amol h - l cm-3}. Obviously, thiosulphate is n o t exclusively oxidized [o sulphate b u t to some e x t e n t t r a n s f o r m e d to other still u n k n o w n products.
U n d e r a n a e r o b i c conditions in the p r e s e n c e of nitrate, t h i o s u l p h a t e t u r n o v e r rates (0.3-5.2 LtmO1 h -1 cm -3) w e r e in the s a m e order as u n d e r aerobic c o n d i t i o n s (Fig. 2b). However, lower a m o u n t s of s u l p h a t e (0.1-0.5 ~mol h -1 cm -3) w e r e formed. Thus. anaerobically, s u l p h a t e was a m m o r oxidation product. Sulphite was n o t a n i n t e r m e d i a t e in t h i o s u l p h a t e transformations that a c c u m u l a t e d d u r i n g the i n c u b a t i o n period. Other possible transformations of t h i o s u l p h a t e i n c l u d e oxidation to products other t h a n sul- phate, r e d u c t i o n or disproportionation. H i g h n i t r a t e - r e d u c i n g activity (up to 10.8 ~tmol h - cm -3} was p r e s e n t in this s e d i m e n t a n d the m a x i m a of nitrate r e d u c t i o n c o r r e l a t e d with m a x i m a of t h i o s u l p h a t e t r a n s f o r m a t i o n (Fig. 2b). T h e highest v a l u e s of b o t h reactions were f o u n d in the first 1-2 cm of the s e d i m e n t , a s e c o n d m a x i m u m was at 3 - 4 cm depth. Both of these m a x i m a correlated well w i t h m i m m u m values of n i t r a t e in the s e d i m e n t (Table 1).
T h e s e d i m e n t n e a r F~ihrdorf
V i a b l e cells, s u l p h u r - o x i d i z i n g b a c t e r i a , c h e m i c a l p a r a m e t e r s 231
sediment
depth (cm)
0
_
4
6
8
10
12
16
1 8 -
20
1 0 4
N
~
L
\
D
/
I l i i
z
i l
i
I 111111 L I 111111
105
106
--~-- aerobic SOB
---E~-- anaerobic SOB
, 0
PSB
I I i Ilkll I I t I illtl [ ['"['1 IFli
10 7
10 8
viable counts * c m - 8 sediment
Fig. 3. Viable counts of sulphur-oxidizing bacteria at F~ihrdorf. S O B = S u l p h u r - o x i d i z i n g bacteria; PSB = Purple s u l p h u r bacteria. Viable counts of a n a e r o b i c SOB were d e t e r m i n e d u n d e r anoxic
232
sediment
depth (cm)
0
2,
6 -
_
1~ 1
12
J. F. I m h o f f , A. S c h n e i d e r & L. P o d g o r s e k
\ .-
@
.~,J
I
-(.~
\
\
\
%
1~ 1
[3-
2-
t
9
SO 4
formation
18
2-
---0- $ 2 0 3
consumption
2 0
0
2
4
6
8
10
transformation rates
V i a b l e ceils, s u l p h u r - o x i d i z i n g b a c t e r i a , c h e m i c a l p a r a m e t e r s
sediment
depth (cm)
0
2
~9
233
4
6
8
1~
14
'~
18
\
10
L i i h i' - , ,
9
J J /
J
J J J
. / . f l -
j /
[]
SO4 2-
/
formation
J . i / / "
---~:~--S2032-consumption
z~
N 0 3
r e d u c t i o n
20
0
2
4
6
8
10
transformation rates
234 J. F. I m h o f f , A. S c h n e i d e r & L. P o d g o r s e k
Table 2. Chemical parameters of a s e d i m e n t from F~ihrdorf. SN = water close to the s e d i m e n t surface; n.d. not d e t e c t e d
Salinity* Chloride Sulphate Sulphate/ Sulphide Nitrate Phosphate % mM m M Chloride" " ~M aM HM SN water I2.85 200.7 11.01 54.9 8 9.1 8.7 Sediment
0 - I cm I3.00 202.8 10.42 51.4 14 5.7 17.8 1-2 cm 13.30 207.7 11.10 53.4 12 45.6 0.I 2-3 cm 14.53 227.0 13.34 58.8 14 6.0 n.d. 3-4 cm 14.51 226.6 13.47 59.5 276 1.7 654.3 4-6 cm 15.17 236.8 13.36 56.4 139 34.6 169.8 6-8 cm 13.26 207.1 11.20 54.1 232 3.4 184.9 8-10 cm 13.92 217.3 10.65 49.0 29 3.4 394.1 10-15 cm 12.52 195.5 8.80 45.0 154 5.2 11.4 15-20 cm 10.49 170.8 8.21 48,1 - 27.0 6.1 9 calculated from the concentration of chloride m % according to Wooster et al. (I969L
* * calculated from the concentrations (mM) a n d multiplied by 1000: (value of s t a n d a r d sea water
is 51.7).
r a t i o w e r e u n u s u a l l y h i g h ( T a b l e 2), i n p a r t i c u l a r a t t h e 2 - 6 c m d e p t h . D i s t i n c t l a y e r s w i t h c h a n g i n g c o n c e n t r a t i o n s of s u l p h i d e I m a x i m u m a t 3 - 8 cm), n i t r a t e ( m i n i m a at 2 4 c m a n d 6 - 1 2 c m m a x i m a at 1 - 2 c m a n d 4 - 6 c m l , a n d p h o s p h a t e ( s t r o n g m i n i m u m a t 1 - 3 c m , s t r o n g m a x i m u m at 3 - 1 0 cmJ w e r e f o u n d . In p a r t i c u l a r , t h e e n o r m o u s c o n c e n t r a t i o n g r a d i e n t of p h o s p h a t e , b e t w e e n v e r y h i g h v a l u e s of m o r e t h a n 650 u M a t a 3 - 4 c m d e p t h a n d u n d e t e c t a b l e t r a c e s at a 2 - 3 c m d e p t h , p o i n t s to h i g h l y a c t i v e t r a n s f o r m a t i o n s . S i m i l a r to t h e s e d i m e n t of H i d d e n s e e , t h i s s e d i m e n t a p p e a r e d to b e a s o u r c e o f p h o s p h a t e for t h e s u p e r n a t a n t w a t e r .
A l t h o u g h l o w e r n u m b e r s of a e r o b i c a n d n i t r a t e - r e d u c i n g s u l p h u r - o x i d i z e r s w e r e p r e s e n t in t h e t o p l a y e r of t h i s s e d i m e n t c o m p a r e d to t h o s e f r o m H i d d e n s e e (Fig. 3), v i a b l e c e l l n u m b e r s i n t h e d e e p e r s e d i m e n t w e r e i n a s i m i l a r o r d e r of m a g n i t u d e i n b o t h s e d i m e n t s . A e r o b i c s u l p h u r - o x i d i z e r s h a d q u i t e c o n s t a n t c e l l n u m b e r s o v e r t h e w h o l e s e d i m e n t c o r e , w i t h a s l i g h t m a x i m u m a t a d e p t h of 8 - 1 0 c m . N i t r a t e r e d u c e r s h a d a s l i g h t m a x i m u m a t 1 - 2 c m a n d c o n s t a n t l y h i g h v i a b l e c e l l n u m b e r s f r o m 4 - 6 c m . A n a p p a r e n t m a s s d e v e l o p m e n t of p h o t o t r o p h i c p u r p l e s u l p h u r b a c t e r i a w a s n o t v i s i b l e . N e v e r t h e l e s s . v i a b l e n u m b e r s of t h e s e b a c t e r i a w e r e h i g h i n t h e t o p l a y e r of t h e s e d i m e n t (2 - 106 c e l l s c m -3 s e e Fig. 3). C o m p a r e d t o a n e a r l i e r s a m p l i n g i n A p r i l 1992, t h e y h a d i n c r e a s e d b y t h r e e o r d e r s of m a g n i t u d e , b u t still w e r e t w o o r d e r s of m a g n i t u d e l o w e r t h a n t h o s e f r o m H i d d e n s e e . G r e e n s u l p h u r b a c t e r i a w e r e p r e s e n t i n i n s i g n i f i c a n t n u m b e r s a n d c o u l d n o t b e q u a n t i f i e d .
Viable cells, s u l p h u r - o x i d i z i n g bacteria, c h e m i c a l p a r a m e t e r s 235
o x i d i z e d to sulphate. T h e s e rates c o i n c i d e d with m a x i m u m v a l u e s of nitrate r e d u c t i o n in the s a m e layers (3.8 ~mol h -1 cm -3 a n d 8.2 ~mol h - l cm -3, respectively) a n d h i g h viable cell counts of nitrate r e d u c i n g bacteria.
D I S C U S S I O N
Q u a n t i t a t i v e e n u m e r a t i o n of s u l p h u r - o x i d i z i n g b a c t e r i a with s e l e c t i v e m e d i a and m e a s u r e m e n t s of potential transformation rates of t h i o s u l p h a t e and nitrate, c o m b i n e d with the c h e m i c a l analyses of p o r e w a t e r i n d i c a t e d the i m p o r t a n c e of b a c t e r i a in the c h e m i c a l p r o c e s s e s occurring w i t h i n the sediments. F u r t h e r m o r e , distinct m a x i m a of s e v e r a l c h e m i c a l p a r a m e t e r s a n d of m e a s u r e d activities r e v e a l e d a c o m p l e x m u l t i l a y e r e d structure of the i n v e s t i g a t e d s e d i m e n t s .
A l t h o u g h nitrate c o n c e n t r a t i o n s in the i n v e s t i g a t e d s e d i m e n t s w e r e q u i t e low (below 10 ttM, at m a x i m a up to 50 ~M), nitrate a p p e a r e d to b e an i m p o r t a n t e l e c t r o n donor for the oxidation of r e d u c e d s u l p h u r c o m p o u n d s in sulphidic sediments. H i g h n u m b e r s of n i t r a t e - r e d u c i n g b a c t e r i a a n d h i g h potential activities of nitrate r e d u c t i o n a n d thiosul- p h a t e oxidation in the s a m e s e d i m e n t layers, i.e. 3-4 cm d e p t h in the H i d d e n s e e s e d i m e n t a n d 1-2 cm d e p t h in the F~hrdorf sediment, point to an active role of t h e s e b a c t e r i a in the r e s p e c t i v e s e d i m e n t layers. A l t h o u g h t h i o s u l p h a t e oxidation is s t i m u l a t e d in the p r e s e n c e of nitrate, s u l p h a t e is not f o r m e d in e q u i v a l e n t quantities. It appears, therefore, that s u l p h a t e is only a m i n o r oxidation p r o d u c t of t h i o s u l p h a t e w h e n nitrate is the e l e c t r o n acceptor. It is c o n c l u d e d that n i t r a t e - r e d u c i n g b a c t e r i a play an i m p o r t a n t role in the t u r n o v e r of r e d u c e d sulphur c o m p o u n d s in anoxic parts of m a r i n e sediments, as l o n g as nitrate is available.
T h e i m p o r t a n c e of aerobic s u l p h u r - o x i d i z i n g b a c t e r i a in the top layer of the H i d d e n - see s e d i m e n t is d e m o n s t r a t e d by significant oxidation rates of t h i o s u l p h a t e to s u l p h a t e a n d by m a x i m a of v i a b l e cell counts in this layer. In the d a r k and u n d e r conditions of b l o c k e d o x y g e n transfer b e t w e e n a t m o s p h e r e a n d w a t e r column, the release of s u l p h i d e from the s e d i m e n t surface into the w a t e r strongly i n c r e a s e d as s h o w n by e x p e r i m e n t s with intact s e d i m e n t cores. T h e r e f o r e . o x y g e n is r e g a r d e d as the most i m p o r t a n t e l e c t r o n a c c e p t o r for o x i d a t i o n of r e d u c e d sulphur c o m p o u n d s in the surface l a y e r of t h e sedi- ments, a n d a e r o b i c s u l p h u r b a c t e r i a play a p r e d o m i n a n t role in t h e s e reactions.
H i g h n u m b e r s a n d e v e n m a x i m a of (facultative) a e r o b i c b a c t e r i a in the a n o x i c parts of the s e d i m e n t s point to their m e t a b o l i c flexibility a n d t h e i r potential of a n a e r o b i c transformations. It is well e s t a b l i s h e d that d e n i t r i f y i n g b a c t e r i a are f a c u l t a t i v e aerobes. A l t h o u g h a g e n e r a l conclusion on the p a r t i c i p a t i o n of facultative a e r o b e s in nitrate r e d u c t i o n in anoxic s e d i m e n t layers is not possible, the o c c u r r e n c e of h i g h n u m b e r s of a e r o b i c s u l p h u r b a c t e r i a in distinct s e d i m e n t layers w i t h m a x i m a of d e n i t r i f y i n g bacteria, active denitrification and m i n i m u m nitrate c o n c e n t r a t i o n s m a y be i n d i c a t i v e of the m e t a b o l i c activity of t h e s e b a c t e r i a u n d e r a n a e r o b i c conditions.
2 3 6 J. F. I m h o f f , A. S c h n e i d e r & L. P o d g o r s e k
for t h e o x i d a t i o n of s u l p h i d e a n d o t h e r r e d u c e d s u l p h u r c o m p o u n d s i n t h e i n v e s t i g a t e d s e d i m e n t s , a s is d e m o n s t r a t e d b y t h e i r h i g h n u m b e r s i n t h e t o p s e d i m e n t l a y e r s of b o t h s a m p l i n g sites. T h e a c t i v e r o l e of t h e p u r p l e s u l p h u r b a c t e r i a i n t h e o x i d a t i o n of r e d u c e d s u l p h u r c o m p o u n d s w i t h i n t h e s e d i m e n t is e v i d e n t f r o m t h e f a c t t h a t m o s t of t h e i r c e l l s w e r e f i l l e d u p w i t h s t o r e d e l e m e n t a l s u l p h u r - a n i n t e r m e d i a t e i n t h e o x i d a t i o n of s u l p h i d e to s u l p h a t e . A d d i t i o n a l l y , i n s e d i m e n t c o r e s w h i c h s t r o n g l y r e l e a s e d s u l p h i d e i n t o t h e s u p e r n a t a n t w a t e r w h e n o x y g e n s u p p l y w a s i n h i b i t e d for 12 h o u r s i n t h e d a r k (13.5 ,~M s u l p h i d e ) , t h e m a j o r p a r t of t h i s s u l p h i d e w a s o x i d i z e d w h e n o x y g e n w a s a l l o w e d to d i f f u s e i n t o w a t e r a n d w h e n t h e s e d i m e n t w a s i l l u m i n a t e d . O n l y 1 to 1.5 ~ M s u l p h i d e w e r e r e l e a s e d u n d e r t h e s e c o n d i t i o n s d u r i n g a 12 h p e r i o d . B l a c k b u r n e t al. (1975) d e m o n s t r a t e d t h e p r e d o m i n a n t r o l e of p h o t o t r o p h i c b a c t e r i a b y s i m i l i a r e x p e r i - m e n t s i n s e d i m e n t c o r e s , too. P h o t o t r o p h i c s u l p h u r b a c t e r i a a n d a e r o b i c c h e m o t r o p h i c s u l p h u r b a c t e r i a c e r t a i n l y a r e c o m p e t i n g s u l p h u r - o x i d i z e r s i n t h e t o p l a y e r s of t h e i n v e s t i g a t e d s e d i m e n t s , i n p a r t i c u l a r b e c a u s e s o m e of t h e p h o t o t r o p h i c p u r p l e b a c t e r i a c a n u s e o x y g e n a s a n e l e c t r o n a c c e p t o r d u r i n g c h e m o t r o p h i c g r o w t h ( K ~ m p f & P f e n n i g , 1980). W e c o n c l u d e t h a t t h e c o m b i n e d a c t i o n of b o t h g r o u p s of b a c t e r i a m o s t e f f i c i e n t l y o x i d i z e s r e d u c e d s u l p h u r c o m p o u n d s i n t h e t o p l a y e r s of t h e s e d i m e n t s . N i t r a t e m a y r e p l a c e o x y g e n as f i n a l e l e c t r o n a c c e p t o r a n d will s u p p o r t o x i d a t i o n of s u l p h i d e ~ i n p a r t i c u l a r w h e n o x y g e n a n d l i g h t a r e l i m i t i n g .
Acknowledgements. This work was s u p p o r t e d by the grant DYSMON A2 of the B u n d e s m i n i s t e r ffir Forschung u n d Technologie of Germany.
L I T E R A T U R E C I T E D
Blackburn. T. H.. Kleiber. P & Fenchel T., 1975. Photosynthetic sulphide oxidation in m a r i n e sediments. - Oikos 26. 103-108.
F e h e n F. & Laue. W. 1956. Beitr~ge zur C h e m i e des Schwefels. 29. Ober die D a r s t e l l u n g von R o h s u l f a n e n . - Z. anorg, allg. Chem. 288. 103-112.
Imhoff. J. F.. 1988. A n o x y g e n i c phototrophic bacteria. In: M e t h o d s in aquatm bacteriology. Ed. by B. Austin. Wiley, Chichester. 207-240.
J 6 r g e n s e n . B. B., 1982. Mineralization of organic m a t t e r in t h e sea b e d - the role of sulfate reduction. - Nature. Lond. 296. 643-645.
J 6 r g e n s e n . B. B. & Bak. F.. 1991. P a t h w a y s and microbiology of thiosulfate t r a n s f o r m a t i o n s and sulfate reduction in a m a n n e s e d i m e n t - Appl. e n v i r o n Microbiol. 57. 847-856.
K~impf, C. & Pfennig, N., 1980. Capacity of C h r o m a t i a c e a e for chemotrophic g r o w t h - Arch. Microbiol. 127, 125-135.
Lorant, I. S., 1929. Uber eine n e u e colorimetrische M e t h o d e zur B e s t i m m u n g d e s Schwefels in Sulfiden. Sulfaten usw, - Hoppe-Seyler's Z. physiol. Chem. 185. 245-266.
Pachmayr, F.. 1960. V o r k o m m e n u n d B e s t i m m u n g yon S c h w e f e l v e r b i n d u n q e n in Mineralwasser. Diss.. Univ. Mtinchen, 48 pp.
Pfennig, N. & TKiper, H. G. 1992. T h e family Chromatiaceae. In: The Prokaryotes. Ed. by A. Balows. H. G. Trtiper, M. Dworkin, W. H a r d e r & K. H. Sehleifer. Springer. N e w York, Vol. 4. 3200-3221. Suckow. R,. 1966. S c h w e f e l m i k r o b e n g e s e l l s c h a f t e n der See- u n d Boddengew~isser y o n H i d d e n s e e .
- Z. allg. Mikrobiol. 6, 309-315.
Widdel. F., 1988. Microbiology a n d ecology of sulfate- a n d sulfur-reducing bacteria. In: Biology of a n a e r o b i c microorganisms. Ed. b y A. J, B. Z e h n d e r . Wiley, New York. 469-585.