The Expression and Regulation of the Po Gene During Development o f the Rat: Regulation of Basal Level and Early Expression
by
Meng-Jen Lee
Department of Anatomy and Developmental Biology
University College London
A thesis presented for the degree of Doctor of Philosophy
ProQuest Number: U642130
All rights reserved
INFORMATION TO ALL USERS
The quality of this reproduction is dependent upon the quality of the copy submitted.
In the unlikely event that the author did not send a complete manuscript and there are missing pages, these will be noted. Also, if material had to be removed,
a note will indicate the deletion.
uest.
ProQuest U642130
Published by ProQuest LLC(2015). Copyright of the Dissertation is held by the Author.
All rights reserved.
This work is protected against unauthorized copying under Title 17, United States Code. Microform Edition © ProQuest LLC.
ProQuest LLC
789 East Eisenhower Parkway P.O. Box 1346
Acknowledgements
I wish to thank Professors Kristjan lessen and Rhona Mirsky for their guidance and enthusiasm throught the course o f this work. I also thank Dr. Myrna Dent and Dr. Eric Parmantier for their help and numerous discussions, especially in in situ hybridization technique. I am grateful to Dr. Angela Brennan who contributed to the set up o f the neural crest cultures and the study o f the regulation o f Po protein, and Dr. Ester Calle for contributing to the study o f the lineage segregation. I thank Dr Louise Morgan, Dr Helen Stewart and Dr. Ziping Dong for their discussion and help, especially in the early days. I also acknowledge Charlotte Dean, Christiane Zoidl and Mary Rahman for technical support and making working in the lab easy.
I am grateful to Dr. V. Pachnis, Dr. D. Ish-Horowilz, Dr. C. Birchmeier, Dr. C. Dickson, Dr. Wilkinson, Dr. G. Lemke, Dr. I. Griffth, Dr. J. Lewis, Dr. D. Henrique for their gift o f plasmids, Dr. B. Zalc for the PLP transgenic mice. I want to thank Dr. Jon Clark and Dr. Paul Martin for the micrscope and photography setup. Dr. Sue Manning for sharing her methods, Steve Harsum and C P Ma for their advice on computer methods.
TABLE OF CONTENTS
TITLE 1
ACKNOWLEDGEMENTS 2
TABLE OF CONTENTS 3
ABSTRACT 5
LIST OF TABLES AND ILLUSTRATIVE MATERIAL 7
ABBREVIATIONS 9
CHAPTER 1 : GENERAL INTRODUCTION 11
1.0 Table o f contents 12
1.1 PNS glia 15
1.2 The molecular phenotypes o f Schwann cells 18
1.3 Neuronal influence on Schwann cells 31
1.4 Wallerian degeneration 34
1.5 Myelination and the structure o f myelinated fibres 36
1.6 Other Schwann cell functions 37
1.7 The Po molecule 37
1.8 The neural crest 42
1.9 Gliogenesis and neurogenesis during the formation o f the dorsal root
ganglion 53
1.10 Axon/Schwann cell interactions during the formation o f nerves 55
1.11 The Schwann cell precursor 56
1.12 The CNS/PNS transitional zone 58
1.13 The gut 59
1.14 The placode 61
1.15 The ear 65
1.16 The nose 75
1.17 Illustrative material 81
CHAPTER 2: Materials and Methods 87
CHAPTER 3: The differential regulation o f Po gene in the 2 mature phenotypes
o f the Schwann cell lineage 110
3.0 Table o f contents 111
3.2 Results
3.3 Discussion 119
3.4 Ilustrative material 121
CHAPTER 4: The study o f Po gene expression in the migrating neural crest and
its derivatives 126
4.0 Table o f contents 127
4.1 Introduction 129
4.2 Results 135
4.3 Discussion 145
4.4 Illustrative material 158
CHAPTER 5: The study o f Po gene expression in the development o f the
inner ear 168
5.0 Table o f contents 169
5.1 Introduction 171
5.2 Results 175
5.3 Discussion 180
5.4 Ilustrative material 196
CHAPTER 6: General Discussion 201
Abstract
List of Tables Table 1-1 Table 1-2 Table 1-3 Table 1-4 Table 2-1 Table 5-1 Table 5-2
Neural crest derivatives 43
Derivatives o f neural crest and placodes, categorised by cell type 63 Contribution to the cranial ganglia by placodes and crest 63 Combinatory code of transcription factors in placodes 65 Restriction sites and polymerase used to generate riboprobes 109 Comparison o f neural crest and neurogenic placode 183
The earliest innervation o f inner ear 190
List of Illustrative Materials
Figure 1-1 Molecular structure o f Po protein 81
Figure 1-2 Diagrammatic representation o f Po orientation in the myelin
membrane 82
Figure 1-3 Neural crest migrating pathway in the trunk 83
Figure 1-4 Gross structure o f inner ear 84
Figure 1-5 Structure o f cochlea and Organ o f Corti 85
Figure 1-6 recycling cells 86
Figure2-1 cDNA encoding the entire Po coding sequence 108 Figure3-1 Po mRNA in the neural crest and in developing sciatic nerves 121 Figure3-2 Axon-dependent and reversible down-regulation o f Po mRNA in
adult non-myelin-forming Schwann cells 123
Figure3-3 Removal o f Schwann cells from axons leads to a fall in myelin
related Po expression, while basal levels are relatively unaffected 124 Figure3-4 Schematic representation o f Po mRNA distribution in cells o f the
Schwann cell lineage 125
RT-PCR o f Po in developing embryo 158
Po expression in ElO 160
Po expression in E l 1 ; comparison to TU J-1 160 Figure 4-1 Figure 4-2 Figure 4-3 Figure 4-4 Figure 4-5 Figure 4-6 Figure 4-7 Figure 4-8
Po expression in E l 2; comparison to TUJ-1, ErbB3 and Krox-20 160 Po expression in E l 4
Developing enteric nervous system Neural crest culture
Figure 4-9 A summary o f Po expression during the generation o f peripheral
glial phenotypes 167
ABBREVIATIONS
3 ’ UTR 3’ untranslated region ADS antibody diluting solution BDH dopamine-p-hydroxylase BMP-4 bone morphogenetic protein
cAMP cyclic AMP
CNPase 2 ’,3 ’,-cyclic nucleotide 3’-phosphodiesterase CNS central nervous system
CNTF ciliary neurotrophic factor CSX cervical sympathetic trunk
DMEM Dulbecco’s modified Eagle medium DREZ dorsal root entry zone
DRG dorsal root ganglia
E ( ) embryonic day
ENS enteric nervous system Gal-C galactocerebroside
GAP-43 growth associated protein 43
GDNF glial cell line derived neurotrophic factor GFAP glial fibrillary acidic protein
IHC inner hair cells ISH in situ hybridization
LIF leukaemia inhibitory factor M AG myelin-associated glycoprotein MBP myelin basic protein
MEM minimum essential medium NCAM neural cell adhesion molecule NGF nerve growth factor
NT-3 neurotrophin 3 NT-4 neurotrophin 4
OBECs olfactory bulb ensheathing cells OHC outer hair cells
Po protein zero P2 myelin protein 2
p75NGF-R p75 the low affinity NGF receptor PBS phosphate buffered saline
PF paraformaldehyde
PLL poly-L-lysine
PLP proteolipid protein
PMP-22 peripheral myelin protein 22 PNS peripheral nervous system PSANCAM polysialyated NCAM
RT-PCR reverse transcriptase polymerase chain reaction SCIP suppressed cAMP inducible POU protein SMP Schwann cell myelinating protein
TV tegmentum vasculosum
1.0 Table o f contents 12
1.1 PNS glia 15
1.2 The molecular phenotypes o f Schwann cells 18
1.2.1 Appearance and regulation o f Schwann cell phenotypes during
development 18
1.2.2 Non-myelinating phenotypes 19
1.2.3 M yelinating phenotypes 20
1.2.4 Molecules expressed by both phenotypes 25
1.2.5 Molecules not specific to Schwann cells but important in PNS
myelination 26
1.2.6 Ion channels 29
1.3 Neuronal influence on the Schwann cells 31
1.3.1 Regulation o f the lineage choice and embryonic development o f the Schwann cell lineage; signalling by p-neuregulin binding to ErbB3
receptor 31
1.3.2 Axonal regulation o f the Schwann cell phenotypes 33
1.4 W allerian degeneration 34
1.5 The myelination and the structure o f myelinated fibres 36
1.6 Other Schwann cell functions 37
1.7 The Po molecule 37
1.8 The neural crest 42
Table 1-1 Neural crest derivatives 43
1.8.1 The experimental approaches 44
1.8.2 The migrating pathways 45
1.8.3 M olecular markers for neural crest 48
1.8.4 Control o f the migration pathway 49
1.8.5 The plasticity and the heterogeneity o f the neural crest population 50 1.8.6 The lineage relationship o f the neural crest derivatives 51 1.9 Gliogenesis and neurogenesis during the formation o f dorsal root ganglion 53 1.10 Axon/Schwann cell interaction during the formation o f nerves 55
1.11 The Schwann cell precursor 56
during early development o f PNS 57
1.12 The CNS/PNS transitional zone 58
1.13 The gut 59
1.14 The placode 62
1.14.1 Origin 62
1.14.2 Organogenesis 62
1.14.3 Contribution to cranial ganglion 62
Table 1-2 Derivatives of neural crest and placodes, categorised by cell type 63 Table 1-3 Contribution to the cranial ganglia by placodes and crest 64 1.14.4 Comparison o f the placode and the neural crest 64
1.14.5 Molecular markers to date 64
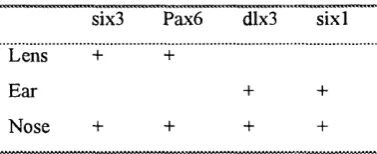
Table 1-4 Combinatory code o f transcription factors in placodes 65 1.15 The ear
1.15.1 General anatomy and corresponding functions o f the inner ear 65 1.15.2 The structure o f sensory organs and their cell types 66
1.15.3 The generation o f the inner ear 67
1.15.3.1 Early morphological changes 68
1.15.3.2 Regional specification 68
1.15.3.3 The specification o f sensory organ versus specification
o f other structures 70
1.15.4 Cell lineages o f the inner ear 71
1.15.4.1 The neurogenic lineage 71
1.15.4.2 The sensory cells: hair cells and support cells 72
1.15.4.3 The lineage o f melanocytes 72
1.15.4.4 Non-sensory, non-melanocyte epithelial cells 73 1.15.5 Factors responsible for the induction and differentiation o f the otic
vesicle 74
1.16 The nose
1.16.1 Terminology 74
1.16.2 An overview o f the olfactory system 75
1.16.3 Plasticity in the olfactory system
1.16.3.2 The ability o f olfactory axons to regenerate in the CNS 77 1.16.4 The origin and development of olfactory Schwann cells 77 1.16.5 The phenotypes o f olfactory Schwann cells; a mixture o f PNS
and CNS phenotypes 79
1.16.6 The lineage o f olfactory ensheathing cells: a mixture o f two cell lineages or two phenotypes o f a same lineage? 80 1.17 Illustrative material
Figure 1-1 M olecular structure of Po protein 81
Figure 1-2 Diagrammatic representation o f Po orientation in the myelin
membrane 82
Figurel-1 Neural crest migrating pathway in the trunk 83
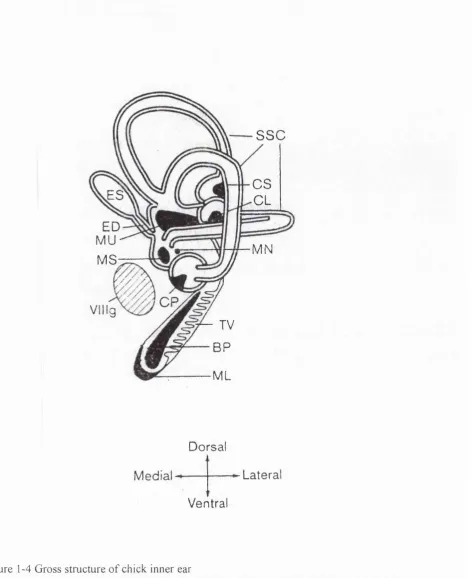
Figure 1-2 Gross structure o f inner ear 84
Figure 1-3 Structure o f cochlea and Organ o f Corti 85
Po is the major myelin protein in the peripheral nervous system (PNS) and plays an important role in myelination. It has been used as a model to study myelination and axon-to-glia effects. It was always believed that Po is expressed postnatally only in myelinating Schwann cells, and the same was assumed in several transgenic experiments designed to use the Po promoter for Schwann cell specific, postnatal expression. A trend o f discovering myelin genes during mid gestation stage (Ikenaka et al., 1992; Nakajima et al., 1993; Timsit et al., 1992; Yu et al., 1994; Zhang et al., 1995) points to a need to study Po expression at a stage when the parent cells o f Schwann cells are generated from the neural crest. In this chapter the Po molecule itself, the regulation o f molecules expressed in Schwann cells, and the development and lineage regulation o f the cell types that are relevant are described.
1.1 PNS glia
Satellite cells
The perikaryon o f a neuron in the DRG or autonomic ganglia is ensheathed by small, flat epithelium-like satellite cells that are continuous with Schwann cells enveloping the axon. Besides providing a continuous cover for neuronal bodies and their processes (Pannese, 1960; 1968), the satellite cells also secrete constituents o f basal lamina to separate the glial cells and ensheathed neurons from other elements o f the extracelluar matrix that are present in the ganglia (Pannese, 1968, Baluk et al., 1985). The position o f the satellite cells relative to the neuronal cell bodies they ensheath changes slowly, possibly associated with synaptic remodelling (Pomeroy and Purves, 1988). The satellite cells share many similarities with Schwann cells. Like Schwann cells, they respond to axotomy by proliferation, but the response o f the satellite cells is generally more rapid (Lu and Richardson, 1991). In addition, satellite cells express some molecules that are common to all peripheral glia. The molecular phenotype o f satellite cells is regulated, often negatively, by factors present in the microenvironment. Schwann cell myelinating protein (SMP) (see below), a molecule expressed by both myelinating and non-myelinating Schwann cells, is expressed in satellite cells o f embryonic but not adult DRG. When cultured in vitro and released from the tightly-packed microenvironment of ganglion, the satellite cells rapidly express SMP, suggesting that constitutive expression o f SMP is inhibited in the DRG (Cameron-Curry et al., 1993). Krox-20, a transcription factor important for myelination, is another molecule that is down-regulated by the microenvironment o f ganglion (Murphy et al., 1996). The ability o f satellite cells to re-express Krox-20 only occurs after the time in development when transition o f precursors to Schwann cells occurs, suggesting that the maturation programme in the satellite cells is on same time scale as that in Schwann cells in peripheral nerves (Murphy et al., 1996). Minor differences exist between the satellite cells in the sensory ganglion and those in the sympathetic ganglion. Satellite phenotypes were further regulated by difference in DRG and sympathetic ganglia, but again these differences are reversible when cells are cultured in vitro (Riidel and Rohrer, 1992).
Perisynaptic Schwann cells are specialised non-myelinating Schwann cells that cover a single large terminal axon and its branches at the highly specialised synaptic zone. The outline o f the elliptical soma o f the perisynaptic Schwann cells can be visualised within the endplate when the nerve terminals are labelled with a fluorescent agent that has high affinity for the axolemma, for example, fluorescent peanut agglutinin (PNA) (Ko, 1987). This type o f Schwann cell is different from other Schwann cells and behaves in some way more like astrocytes. Unlike non-myelinating Schwann cells, they normally do not express growth associated protein 43 (GAP-43) (Curtis et al., 1992; W oolf et al., 1992). After nerve transection, perisynaptic Schwann cells respond to denervation by up-regulating GAP-43 in 18 hours, while for myelinating Schwann cells it often takes several weeks (Tetzleff et al, 1989; Curtis et al., 1992). Glial fibrillary acidic protein (GFAP) level is also up-regulated in perisynaptic Schwann cells during denervation, utilizing a mechanism that involves blockage o f neurotransmitter rather than the loss o f axonal contact in the myelinating Schwann cells (Georgiou et al., 1994; lessen et al., 1990). Both myelinating and non myelinating Schwann cells proliferate after denervation while perisynaptic Schwann cells do not divide (Clemence et al., 1989; Connor and McMahan, 1987). They form long processes that extend ahead o f presynaptic axon terminals but retract on reinnervation (W oolf et al., 1992). In frog the perisynaptic Schwann cells respond to neurotransmitters with a release o f intracellular Ca^ (Jahromi et al., 1992). These differences suggest that perisynaptic Schwann cells are specialised to respond to events occurring at synapses.
The Schwann cells
1.2 The molecular phenotypes of Schwann cells
The differentiation o f myelinating Schwann ceils is marked by dramatic morphological differences that are involved in myelin formation. The myelinating cells express a whole set o f highly regulated molecules to meet this task. These includes protein zero (Po), myelin basic protein (MBP), myelin-associated glycoprotein (MAG), proteolipid protein (PLP), PMP-22 (peripheral myelin protein 22), myelin protein 2 (P2), 2’,3’,-cyclic nucleotide 3’-phosphodiesterase (CNPase) (references see bellow). Another set o f molecules are specifically expressed in mature non-myelinating Schwann cells. This group o f molecules includes p75, the low affinity NGF receptor (p75NGF-R), neural cell adhesion molecule (NCAM), Ll/N gC A M (neural-glia cell adhesion molecule), GFAP, two proteins recognised by rat-specific monoclonal antibodies, A5E3 and Ran-2, and GAP-43 (references see follows). Although again specifically regulated, the functions o f this set o f molecules, some o f which are shared by CNS glia, are mostly unknown, apart from N-CAM and L I (see follows). Among molecules expressed by the two phenotypes o f Schwann cells, some are not specific to the Schwann cell lineage but their expression marks important stages o f the development o f the Schwann cell lineage. These include S I00, vimentin, laminin, and galactocerebroside (Gal-C) (references see bellow). Some are expressed by both phenotypes and are restricted in the Schwann cell lineage. This type o f molecules includes seminolipid 0 4 , 0 8 and 0 9 (references see below), the function o f which are unknown. Some transcription factors and other proteins that are crucial in myelination are described below.
1.2.1 Appearance and regulation of Schwann cell phenotypes during development
The transcription factor Krox-20 is expressed in the entire length o f spinal nerves o f E l 5 mouse embryo (equivalent o f E l 7 rat), after the transition from precursor Schwann cell, soon after S I00 expression in the Schwann cells (Topilko et al., 1994; Murphy et al., 1996). By E l 8 most Schwann cells also express the lipid antigen 0 4 , which first appears in the S I00 positive cell population at E l 6 (Mirsky et al., 1990). 0 4 is believed to be the sulphated form o f Gal-C that appears two days later in development, but it is also possible that antibody to 0 4 also detects an antigen other than sulphated Gal-C (Mirsky et al., 1990). By E20 95% o f the S I00 positive Schwann cells are also 0 4 positive (Mirsky et al., 1990). The expression o f Gal-C is first detected around birth in the myelinating Schwann cells and is only seen in the non-myelinating Schwann cell after the maturation of this cell type, which occurs around the 3rd week after birth (lessen et al., 1985). Most o f the phenotypes found in non-myelinating but not myelinating Schwann cells are also expressed by the Schwann cell precursor at E l 4-15, except for GFAP and S-100, which is expressed in all Schwann cells (lessen et al., 1990). GFAP expression is first detected during development at E l 8 in cells that become myelinating Schwann cells as well as those that will become non-myelinating (Jessen et al., 1990). After birth the phenotypes o f the presumably homogenous immature Schwann cells undergo dramatic changes depending on the type o f axons they associate with. The development o f myelinating Schwann cells in the nerve involves an initial up-regulation o f the myelin proteins, for example, the induction o f Po, MAG, MBP, and P2. This is followed by the down- regulation o f molecules associated with the non-myelinating phenotype, including GFAP, N-CAM, A5E3 and p75NGF-R (Jessen et al., 1990; Jessen et al., 1987b). In the adult rat, all these down-regulated proteins are essentially undetectable by immunohistochemical methods in myelinating Schwann cells. The phenotypes o f non-myelinating Schwann cells remain mostly similar to the more immature phenotypes, except for expressing Gal-C 3 weeks after birth (Jessen et a., 1985; Eccleston et al., 1987b).
1.2.2 Non-myelinating phenotypes
two proteins recognised by rat-specific monoclonal antibodies, A5E3 and Ran-2, and GAP-43 (Yen and Field, 1981; Jessen and Mirsky, 1984; Jessen et al., 1984; Ni eke and Schachner, 1985; Mirsky et al., 1986; Jessen et al., 1990; Stewart et al., 1992). This group o f molecules tends to be regulated in the opposite direction to the way the myelinating phenotype is regulated. These markers are down-regulated in myelinating Schwann cells after the expression o f myelin proteins and remain unchanged in the non-myelinating cells. p75NGF-R are down-regulated by treatments elevating the intracellular cAMP level (Mokuno et al., 1988; Lemke and Chao, 1988 Morgan et al.,
1991). p75NGF-R, NCAM, L I, GAP-43, GFAP and A5E3, are up-regulated during denervation (Curtis et al., 1992; Daniloff et al., 1986; Jessen et al., 1987b; Martini and Schachner, 1988; Taniuchi et al., 1986, 1988). The maintenance or maturation o f the non-myelinating phenotypes is believed to be by a default pathway, but recent results suggest that additional signals, probably regulated by TGF-(3, may be needed for the expression o f the full non-myelinating phenotype. In vitro TGF-P is shown to up-regulate the expression o f LI and NCAM, besides blocking cAMP-induced expression o f Po and the lipid antigens Gal-C and 04 (Mews and Meyer, 1993; M organ et al., 1994; Stewart et al., 1995). Much less known is about the significance o f the phenotype o f non-myelinating Schwann cells than about the myelin proteins. Being adhesion molecules, NCAM and LI may be involved in axon Schwann cell interactions at the premyelination stage, as well as in mature unmyelinated fibres (Seilheimer and Schachner, 1988; Martini and Schachner, 1986; Wood et al., 1990). p75NGF-R probably plays a role in grabbing the nerve growth factor (NGF) on the Schwann cell surface, making it available for uptake and transport by the axons during axonal growth (Johnson et al., 1988).
1.2.3 M yelinating phenotypes Po (see below)
MBP
1985; Newman et al., 1987). They are a common participant in all myelin sheaths, even in the most primitive vertebrates (Saavedra et al., 1989). Different upstream regulatory sequences of MBP in the CNS and PNS have been identified (Gow et al.,
1992). Both oligodendrocytes and myelinating Schwann cells target MBP proteins to myelin via translocation o f MBP mRNA to the myelin sheath (Colman et al., 1982; Trapp et al., 1987; Ainger et al., 1993). A role for MBP in the structure and compaction o f the major dense line of the myelin sheath was shown by the phenotypes o f two dysmyelinating mice lines with a mutated MBP gene (Privât et al., 1979; Popko et al, 1988; Mol ineaux et al., 1986). In the shiverer and shiverer myelin- deficient mice, myelin is scarce in the CNS, the residual sheaths are not compacted at the major dense line, although PNS myelin appears almost normal, probably compensated for by Po (Martini et al., 1995a). When MBP is introduced by transgenic techniques into a shiverer mouse, a normal sheath is restored (Readhead et al., 1987; Kimura et al., 1989). A regulatory role in myelination for MBP is also proposed. It is known that exon II containing MBP (MBPexII) isoforms distributes throughout cytoplasm and often in the nucleus. It is now known that MBPexII is actively targeted to the nucleus and its peptide segment provides a nuclear localisation signal and probably also regulates the segregation o f MBP mRNA to the cell processes (Pedraza et al., 1997).
MAG
have roles in initiating enwrapment and promoting neurite extension in early myelination (Johnson et al., 1989). The presence o f 72 IcDa form o f MAG in the large multivesicular bodies suggests another possible function for MAG in retrograde transport o f membrane components from the periaxonal space to the oligodendrocyte cell body (reviewed by Hudson 1990). No natural mutant for MAG has been described to date, although abnormal distribution o f MAG in quaking mice, which is not mutated in the MAG gene, has been reported (Trapp et al., 1988). MAG inhibits neurite extension from older neurites via interaction with a sialoglycoprotein in culture, but not for neurites from newborn DRG neurons (DeBellard et al., 1996; Tang et al., 1997). A study of MAG knockout mice showed that regeneration is better in mice lacking MAG, supporting an inhibitory role for MAG in vivo (Filbin, 1995).
PLP
dysmyelination o f CNS axons in white matter tracts (3) abnormal myelin compaction (4) premature death o f oligodendrocytes (summarised in Klugmann et al., 1997). In the PNS the PLP/DM-20 is a minor component, constituting less than 1% o f total myelin protein (Lemke, 1993). PLP has recently been identified in PNS compact myelin, and the new discovery o f a null allele o f PLP with PNS demylination demonstrates its role in proper myelin function in the PNS as well as in the CNS (Garbern et al., 1997).
PMP-22/ gas/CD25/SR13/PASII
slowly progressive distal muscle atrophy and weakness starting from second or third decade, heterozygous deletion of the same chromosomal region leads to HNPP (Chance et al., 1993) that is characterized by repeated weakness which is caused by pressure or trauma to affected nerves.
Genes encoding putative myelin assembly proteins; P2, CNPase
P2, a 14.8 kDa basic protein, is a minor component o f peripheral myelin, accounting
for only 0.05% -l% o f total myelin protein. It is only present in compact myelin and not in the Schmidt-Lantermann incisures (Hahn et al., 1987). It is speculated that this protein is involved in the synthesis and transport of long chain fatty acids (reviewed by Hudson 1990). Another protein thought to be involved in the assembly o f myelin is CNPase (2 ’,3’-cyclic nucleotide 3 ’-phosphodiesterase). CNPase is a cytoplasmic protein expressed in glial cells and some non-neuronal tissues as well. The 2 isoforms are 46 and 48 kDa. Developmental expression o f the CNPase gene during peripheral nerve development, unlike in the CNS, does not correlate with myelin formation (Stahl et al., 1990). CNPase in the CNS may take part in intercellular transportation o f mRNA, guiding specific proteins to their site o f myelin assembly (Hudson, 1990)
Periaxin
is expressed diffusely in the cytoplasm (Dytrych et al., 1998). In rat the expression o f periaxin starts at birth and peak at 8 to 20 days after biith, which is a period o f active myelination. Like MAG, in compacted myelin it is only expressed in the periaxonal sheath, and is believed to be involved in initial enwrapment o f the axon and in the periaxonal region o f the Schwann cell membrane (Gillespie et al., 1994).
1.2.4 Molecules expressed by both phenotypes SlOO
Calcium-binding protein SlOO is expressed by both non-myelin and myelinating cells from several different species in vivo and in vitro. Beside the nervous system, it is also present in melanocytes, some CNS glial cells and some neurons. Its prevailing expression in all Schwann cells in mature nerves, in culture, in denervated nerve trunks and by all embryonic Schwann cells except for the Schwann cell precursor, but not in fibroblast or perineurial cells, makes it a good operational marker for cells o f Schwann cell lineage (reviewed by Mirsky and lessen, 1990). The expression o f SlOO in cells o f Schwann cell lineage marks the transition from precursor cells to the Schwann cells, which in rat happens in the sciatic nerves between E l 5 and E17 (lessen et al., 1994). The induction o f SlOO, unlike many other Schwann cell phenotypes, is irreversible once triggered.
Gal-C
detected around birth in the myelinating Schwann cells, and in the non-myelinating cells it is only detected after the axons were separately located within different troughs o f the Schwann cell cytoplasm, the final stage o f morphologenesis o f the unmyelinated fibre (Jessen et al., 1985; Eccleston et al., 1987b).
04
The antigen which 0 4 monoclonal antibody recognizes is the sulphated form o f Gal- C and perhaps some other unidentified lipids as well (Bansal and Pfeiffer, 1987). The 04 epitope is expressed two days earlier than Gal-C during development o f both oligodendrocytes and Schwann cells, while sulfatide is synthesised from Gal-C using the enzyme galactosyl sulfotransferase (Sommer and Schachner, 1981; Wolswijk and Noble, 1989; Mirsky et al., 1990). It is possible that 0 4 recognises epitopes other than sulfated Gal-C in Schwann cells (Mirsky et al., 1990). Nonetheless, the antigen which 0 4 antibody recognizes is regulated by axonal signals and elevation o f cAMP in a way similar to that of Gal-C (Mirsky et al., 1990). During development, the expression o f 0 4 antigen is first detected in rat El 6 - 17, and by E l 8 most o f the cells dissociated from the sciatic nerves are 0 4 positive, a stage coinciding with the peak proliferation o f embryonic Schwann cells (Mirsky et al., 1990). Sulfatides were shown to bind selectively to Schwann cell basal lamina (Roberts et al., 1985). This up-regulation o f 04 may be related to basal lamina production (Mirsky et al., 1990).
1.2.5 Molecules not specific to Schwann cells but important in PNS myelination SCIP (O ct-6/Tst-l)
specific populations o f neurons during development (Lemke, 1993) and in neonatal testes and epidermis (He et al., 1989). In the Schwann cell lineage, the earliest expression reported is in Schwann cell precursors (Blanchard et al., 1996) and it peaks at one day after birth (Monuki, 1989). Purified Schwann cells in vitro do not express SCIP, but mRNA can be induced by elevation o f cytoplasmic cAMP (Monuki et al., 1989).
Like the myelin proteins, SCIP expression is closely regulated by axonal contact. A high level o f SCIP mRNA was detected during development and regeneration, and axotomy results in rapid down-regulation o f this gene (Scherer et al., 1994). That SCIP expression is well in advance o f myelination, and in transient assays the SCIP protein binds to the promoter o f the gene encoding Po to repress its transcription (He et al., 1991), suggests a role for SCIP during myelinalion. Indeed the importance o f SCIP in the enwrapping o f the axon is demonstrated by several mutations in which SCIP function is defective. The implication from observation o f these mutations, however, is not consistent. Weinstein et al. (1995) designed transgenic mice in which a dominant negative antagonist o f SCIP prevents normal SCIP DNA binding. This antagonist was expressed under a Po promoter to be specific to the Schwann cells. This results in premature myelination, over expression o f myelin specific gene products, and hypermyelination. On the other hand, null mutation o f SCIP results in a constraint or delay of myelination (Bermingham et al., 1996; Jaegle et al., 1996), suggesting that SCIP is important in initiating the myelination process.
Krox-20
Krox-20, originally isolated as an immediate-early seium response gene, encodes a transcription factor with a DNA binding domain consisting o f three C2H2-type zinc
crest-derived boundary cap cells and glial cells o f cranial and spinal nerve and ganglion (Wilkinson et al., 1989; Herdegen et al., 1993; Topilko et al., 1994). The expression o f Krox-20 is important in maintenance o f rhombomere 3 and 5. The hind brain o f mice homozygous for the Krox-20 mutation are disrupted in that rhombomeres 3 and 5 are completely eliminated (Schneider-Maunoury et al., 1993, 1997). Furthermore, Krox-20 is suggested to be a key regulator o f rhombomere- specific gene expression in the developing hindbrain (Sham et al., 1993; Nonchev et al., 1996; Seitanidon et a l, 1997). It can interact with multiple binding sites in the promoter region o f several Hox genes and can induce ectopic expression o f the Hox genes in vivo (Sham et a l, 1993; Nonchev et a l, 1996; Seitanidon et a l, 1997). Krox-20 is also an important regulator during the maturation o f peripheral glia and in the process o f myelination (Topiko et a l, 1994; Murphy et al., 1996). The expression o f Krox-20 in PNS coincides with the acquisition o f maturation, demarcated by the immunoreactivity o f SlOO, after the transition from the Schwann cell precursor (Murphy et a l, 1996). The expression o f Krox-20 in the Schwann cell lineage is regulated by axonal contact and cAMP elevation in a way similar to that o f the myelinating proteins but it is also up-regulated by neuregulins (Murphy et al., 1996). The observation that myelination is prevented and myelin proteins Po and MBP are absent from myelin in Krox-20 knock out mice demonstrates the importance o f Krox- 20 in regulating the expression and assembly o f proteins necessary for myelination (Topilko et al., 1994).
QKI proteins
correlation o f the QKI expression and qk^ mutants also suggests that the QKI proteins are regulators o f myelination (Hardy et al., 1996).The three mRNAs transcribed from this gene, 5, 6 , and 7 kb in length respectively, differ only in their 3 ’ end and 3’
untranslated regions. The predicted amino acid sequences o f this mRNA contain a KH domain, which is believed to be associated with regulation o f cellular RNA metabolism (Musco et al., 1996). The QKI proteins are not restricted to the nervous system. They are also expressed in heart, lung, testis, and during development in the neuroectodermal cells of the neural tube (Ebersole et al., 1996). In the nervous system, they are normally expressed in both the glial cells o f CNS and PNS. The distribution o f QKI 6 and 7 are identical. The QKI proteins are present in the
myelinating cells o f CNS and PNS, with QKI 5 mostly in the nucleus and QKI 6 and
7 in the cytoplasm. They are also expressed in astrocytes and non-myelinating Schwann cells (Hardy et al., 1996).
Connexin 32
Connexin 32 was originally found as a liver gap junction protein (Paul, 1986), and was only recently identified as a myelin-related protein in the PNS and CNS (Scherer et al., 1995). In the peripheral nerve, it is believed to form gap junctions that shorten the communication pathway between the periaxonal cytoplasm and the peripheral cytoplasm in which the cell nucleus is located (Scherer et al., 1995). Molecular genetic studies have shown that gene mutation o f connexin 32 (Cx32), a gap junction protein, results in the PNS abnormality, X-linked Charot-Marie-Tooth disease (CMTX) (Bergoffen et al., 1993; Bone et al, 1995; Bruzzone et al., 1994; Fairweather et al., 1994; lonasescu et al., 1994). Although it is expressed in many tissues, tissues other than the PNS appear to be normal (Bergoffen et al., 1993). The relationship between loss o f connexin 32 function and CMTX disease is still unclear. Reduced junction permeability caused by decrease in either pore size or open channel probability, resulting in reduced permeation o f second messengers such as cAMP through gap junctions, probably causes the demyelination and axonal degeneration (Oh et al., 1997).
Mammalian Schwann cells in culture express at least six kinds o f voltage-dependent ion channels, and others may not have been detected yet. Both voltage-dependent sodium and potassium currents have been recorded using whole-cell patch clamp technique (Chiu et al., 1984). Three other voltage-dependent channels were detected when measuring the outward current: a chloride channel and two types o f potassium current (Howe and Ritchie, 1988). A calcium-dependent cation-selective channel and anion selective channels have also been detected in Schwann cells (Bevan et al., 1984; Gray et al., 1984). Glial ion channels are generally similar to their neuronal counterparts, except for some sodium channels. (Mi et al., 1995; Barres et al., 1990). Using whole-cell patch-clamp recording, Schwann cells in culture also express voltage-gated potassium and sodium currents. These currents occurs at lower density than in many neurons, but are qualitatively similar to neuronal currents (Shrager et al., 1985). The expression o f sodium and potassium channels are different in Schwann cells o f two phenotypes, in vitro and when Schwann cells were acutely dissociated. Outward sodium channels and potassium currents have been recorded in non-myelinating Schwann cells but not in myelinating cells. However, inwardly rectifying potassium currents were found in both phenotypes (Chiu 1987, 1988). Transection o f nerves results in the appearance of sodium channels in myelinating Schwann cells but down regulated the sodium channels in the non-myelinating cells, showing another example o f axonal regulation o f Schwann cell phenotype (Chiu,
1988).
It has been suggested that glial ion channels play a role in regulating the extracellular potassium. A rapid release o f accumulated potassium from neuron to extracellular space happens whenever there is neuronal activity. It has been demonstrated that elevation o f intracellular potassium in glial cells and depression in neurons occurs during neuronal firing. It has also been shown that these potassium homeostatic mechanisms involve passive fluxes o f ions through channels instead o f active transport o f ions by pumps or extracellular diffusion (reviewed by Barres et al.,
compensatory chloride channel (Coles et a l, 1989; Barres et a l, 1988; Ballanyi et a l, 1987).
1.3 Neuronal influences on Schwann cells
Different aspects o f glial cell differentiation have been shown to be modulated by interactions with neuronal element, i.e., the neuronal body and the axons (for a review see Reynolds and Woolf, 1993; Mirsky and Jessen, 1998). Broadly speaking, neurons exert four classes o f action on Schwann cells: survival, stimulation o f differentiation, inducing or repressing proliferation; and modifying the migration/growth o f the Schwann cells. Neuronal factors exert an important influence on Schwann cells during several developmental stages. During early development o f the lineage, Schwann cell precursors (see 1.1 1) survive if cultured in the presence o f neuron-
derived factors, which in term promote the maturation and proliferation o f precursor to Schwann cells (Jessen et al., 1994; Dong et al., 1995). The dependence on axons for survival gradually becomes smaller during development, and adult Schwann cells are independent o f neuronal factor for survival, but low levels o f neuron secreted factors are needed to prevent immature Schwann cells and teloglia undergoing apoptotic death (Grinspan et al., 1996; Syroid et al., 1996). Later in postnatal development two types o f axons direct the differentiation o f immature Schwann cells into two phenotypes in a highly ordered fashion (Jessen et al., 1990; see following section). Upon denervation, the Schwann cells respond to the mitogenic axolemma and proliferate (Salzer et al., 1980; Ratner et al., 1988). Several o f the neuronal related actions on Schwann cells are regulated by neuregulin and are discussed below.
1.3.1 Regulation of the lineage choice and embryonic development of the Schwann cell lineage: signaling by (3-neuregulin
muscle and epithelial cells (for reviews see Peles and Yarden, 1993; Carraway and Bruden, 1995; Marchionni, 1995; Lemke, 1996). Neuregulins modulate the cell response via binding to tyrosine kinase receptors o f the erbB family. Neuregulin binds to erbB3 and erbB4 with high affinity, but not the erbB2 (HER2) and erbBl (EGF) receptors (Peles et al., 1992, 1993; Plowman et al., 1993; Carraway et al.,
1994). However, erbB2 is capable o f forming heterodimers with erbB3 or erbB4 and by receptor cross-phosphorylation initiates downstream signalling within the cell (Holmes et al., 1992; Wen et al., 1992; Carraway and Cantley, 1994; Beerli et al.,
1995).
this survival and maturation o f Schwann cell precursors is shown by experiments in which soluble hybrid protein that contains the extracellular domain o f the erbB4 receptor, a highly specific receptor for neuregulin, prevents the axonal rescue and subsequent maturation o f Schwann cell precursors (Dong et al., 1995). Several observations in vivo support the idea that P neuregulin is the neuronal derived factor that directs the development o f the Schwann cell precursors. Very high level o f expression is seen in both DRG and ventral horn motor neurons at E l 4 and later ages in rat (Meyer and Birchmeier, 1994; Bermingham-McDonagh et al., 1997; Marchionni et al., 1993) Very few Schwann cell precursors were seen in the ventral root and growing emhi yonic nerves in mice deficient of neuregulin or its receptor, ErbB3, again suggesting that neuregulin plays an important role in the survival o f Schwann cell precursors (Meyer and Birchmeier, 1995; Riethmacher et al., 1997). Furthermore, neuregulin stimulates postnatal Schwann cell proliferation and inhibits apopototic Schwann cell death. The axonal signal which is mitogenic to Schwann cells is probably regulated by membrane-bound neuregulin and Schwann cell associated ErbB2 an i ErbB3 (Morrissey et al., 1995; Reithmacher et al., 1997). Neuregulin rescues Schwann cells from developmentally regulated programmed cell death (Syroid et al., f^96). P neuregulin was also reported to promote Schwann cell migration and neuritc outgrowth (Mahanthappa et al., 1996; Calle, Mirsky, Jessen, unpublished), which might be important during the initial interaction between cells o f the Schwann cell lineage and the growing neurites.
1.3.2 Axonal r e g u l a t i o n of Schw ann cell phenotypes
promptly undergo numerous and radical changes in morphology and gene expression which result in a developmentally regressed phenotype. For the myelinating Schwann cells, the loss o f axonnI contact results in loss o f myelin markers and an expression o f an immature phenotype that is very similar to the mature non-myelinating phenotype. This reversion o f the phenotypes can be seen when cells are dissociated from the adult nerve and are plater! in culture without neurons, or transplanted to a nerve environment which will induce myelination in Schwann cells (Aguayo et al., 1972; Weinberg and Spencer, 1976; Trapp et al., 1988; Mirsky et al., 1980; Lemke and Chao, 1988; Jessen & Mirsky 1991), Apart from Schwann cells, sub-types o f PNS glia are also known lo depend on continuous axonal contact to maintain their phenotypes. For example, the phenotype o f teloglia is regulated by neurotransmitter released by presynaptic axons (Jahomi et al., 1992), and some myelin proteins are actively down-regulaieci in enteric glia and satellite cells (Dulac et al., 1990; Cameron-Curry et al.. 1993). In contrast to the molecules specific to the myelinating Schwann cells, the expression o f Gal-C, 0 4 antigen, the lipid antigens 0 8 and 0 9 , are up-regulated in both 'Ahwann cell pathways during development (Eccleston et al., 1987a). The expression of these lipids, like the proteins, is axon-dependent and is rapidly down-regulated when Schwann cells are withdrawn from axonal contact (Jessen et al., 1987a). These axonal signals, although crucial in the differentiation o f Schwann cells, are still not clear. Treatments that produce elevation o f intracellular cyclic AMP levels have widespread effects on the phenotypes o f the Schwann cells (Mirsky et al., 1990; Morgan et al., 1991; Sobue et al., 1986). Besides synergising with growth factors to promote Schwann cell proliferation, they induce expression o f 0 4 and Gal-C and up-iegulate the expression o f myelin genes such as Po, MBP, and CNPase (Mirsky and .lessen, 1990; Lemke and Chao, 1988). The state o f Schwann cells in the cell cycle, as well as the extent o f maturation o f the cells, seems to contribute to the choic e of programs following elevation o f intracellular cAMP levels (Mirsky et al., 1990; Morgan et al., 1991; Mirsky and Jessen, 1990).
1.4 The Wallerian degeneration system
degeneration. The reaction at the site o f the injury differs depending on whether the sheath o f the injured nerve fibre is severed. The scale o f macrophage participation is larger, accompanied by an early inflammatory reaction, when the sheath is ruptured (Sunderland, 1978). The time course and extent of degeneration also differs depending on the type o f insult, distance from the neuronal body, diameter o f axon, age o f animal, as well as species. In general, the axon loses action potential, fast or slow transport and neuronal transmitter secretion immediately after axotomy (Weller and Carros-Navarro, 1977; Sunderland, 1978).The onset o f degeneration o f the axon was reported from within 1 0 minutes in finer axons after mild crush, or in some cases
in 48 hours (Sunderland, 1978). This is followed by a down-regulation of the myelin genes (see previous section). The first week sees breakdown o f axonal component and release o f axonal cytoplasm, and the myelin sheaths break up into ovoids. Over the next few weeks further degeneration includes breakdown o f axonal fragments and myelin by Schwann cells and macrophages, and lysosomal enzymes digest the lipids (Sunderland, 1978). The basal lamina that surrounds each Schwann cell in the distal nerve stump remains intact and the Schwann cells remain within their basal laminae while they proliferate after axotomy (reviewed by Scherer and Salzer, 1996). This Schwann cell multiplication results in the formation o f bands o f Bungner.
The fragmented axonal membrane, and myelin debris were thought to contribute to the mitogenic effect on Schwann cells seen in Wallerian degeneration (Salzer et al., 1977; Salzer and Bunge 1980; Sobue and Pleasure, 1985; Bigbee et al., 1987; Clemence et al., 1989). The axonal signal might involve neuregulin signaling via its receptor ErbB2 (Morrisey et al., 1995). It has also been suggested that macrophages that have phagocytosed myelin produce a conditioned medium that is mitogenic for the Schwann cells (Baichwal et al., 1988), and indeed the peak number o f macrophages present in the distal stump coincides with proliferation o f non myelinating cells and precedes that o f myelinating cells by one day (Clemence et al.,
the chamber did not proliferate, and the myelin was discarded but not phagocytosed (Beuche and Freide, 1984; Scheldt and Friede 1987). Other roles o f macrophages in the degenerating distal stump including recycling lipids from decomposed myelin by producing apolipoprotein E (Snipes et al., 1986). Furthermore, macrophages are known to produce cytokines such as interlukin-1 and interferon a and (3 (Klein,
1990).The roles o f macrophages in the degenerating distal stump are still not fully understood, but is likely to involve clearing of debris and stimulation o f proliferation in Schwann cells.
1.5 Myelination and the structure of myelinated fibers
paranodal area. The periaxonal space is reduced to 3 nm and crossed by bands or septa (reviewed by Bray et al., 1981). The functional significance o f this contact might be to restrict sodium channels to the nodal region (Black et al., 1990; Salzer, 1997), to do with ion coupling between axons and Schwann cells (Barres, 1990), or to ensure electrical isolation o f the nodal and internodal extracellular spaces (Livingston et al., 1973). PNS myelin differs from that o f the CNS in that its inner and outer myelin lamellae are not compacted but contain thin layers o f cytoplasm (Mugnaini et al., 1977). The cell cytoplasm is connected by channels called 'Schimidt-Lanterm ann’ incisures. This continuity o f Schwann cell cytoplasm within the myelin sheath may serve as a channel to transfer proteins from the Schwann cell body, or transport phospholipids from axon to myelin (reviewed by Bray et al, 1981).
1.6 Other function o f the Schwann cells
Schwann cells were originally believed to be structural cells and act as an adhesion and nurturing component for the axons, but modern research suggests that they are playing active roles in the nervous system, like the neuronal component. The Schwann cells promote neuronal survival and differentiation by a series o f actions (reviewed by Reynolds and Woolf, 1993). Growth factors produced by Schwann cells are known to have an important role in promoting neuronal survival and differentiation (reviewed by Scherer and Salzer, 1996; Mirsky and Jessen, 1998). The restriction o f the ion channels o f myelinated axons to the paranodal regions is regulated by Schwann cells. Following demyelination, sodium channel expression in the axonal membrane becomes diffuse and is restored to clusters in new nodes o f Ranvier by the presence o f Schwann cells during re-myelination (Dugandzija- Novakovic et al., 1995). Schwann cells also express different types o f ion channels to maintain the ionic balance o f axons (Beven, 1990; Barres et al., 1990). During increased nerve terminal activity, additional transmitter release could trigger changes at peri-synaptic Schwann cells that would stabilize the synapse or enhance synaptic efficacy (Georgiou et al., 1994).
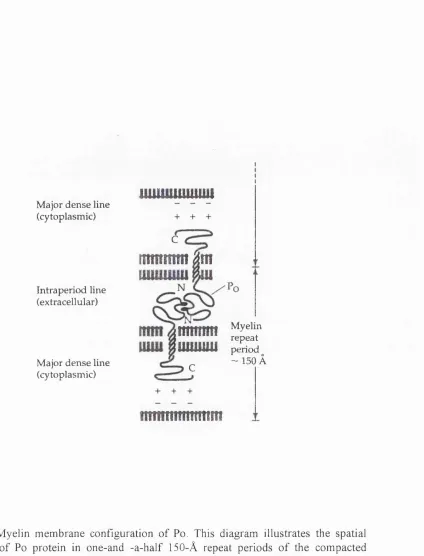
Po is an integral membrane glycoprotein o f molecular weight 28 kDa. The human Po gene, MPZ, was shown to map to chromosome Iq22-q23 (Hayasaka et at., 1993) and the rat Po gene has been isolated, sequenced and analyzed (Lemke and Axel, 1985; Lemke et al., 1988). In rats and mice, this gene consists o f six exons distributed over 7kb o f DNA. The topographic arrangement o f these exons in the genome is consistent with the functional segregation o f the Po protein into extracellular, membrane- spanning, and cytoplasmic domains (Lemke et al., 1988). The primary amino acid struture o f Po has been deduced from cloned cDNAs (Lemke and Axel, 1985) and bovine Po has been directly determined by protein sequencing (Sakamoto et al., 1987). It consists o f a single membrane spanning region, a large extracellular hydrophobic region and small basic cytoplasmic region (Figure 1-1). Being a primitive member o f the immunoblobulin-related protein family, the extracellular domain o f Po is highly glycosylated (see below) and is responsible for the homophilic adhesion properties o f Po and the formation o f the intraperiod line o f compacted myelin (Figure 1-2). In vitro the recombinant extracellular domain is capable o f forming tetramers and dimers, and the same interaction is believed to play an essential role in forming a network to secure the adhesion between apposing Schwann cell membranes (Shapiro et al., 1996). Po molecules are believed to interact via electrostatic charges with the negatively charged lipid bilayer o f the opposing memebrane via its basic intracellular domain in order to form the major dense line in myelin (Ding and Bruden, 1994) (Figure 1-2). Apart from the secondary structure, various post-translational modifications are also believed to contribute to Po function. The Po molecule undergoes many post-translational modifications, including glycosylation (Kitamura et al., 1976; Matthieu et al., 1975; Quarles, 1980), phosphorylation (Brunden and Poduslo, 1987b; Singh and Spritz 1976), sulphation (Matthieu et al., 1975), and acylation (Agrawal et al., 1983). Among these modifications, HNK-1-reacting glycans and mannose-rich N-glycans are potentially involved in cell adhesion (Bollensen et al., 1987; Griffith et al., 1992; Brunden, 1992).
in which Po is neutralized by antisense mRNA (Owens and Boyd, 1991; Giese et al., 1992; Martini et al., I995a,b). Defects in the human myelin protein zero gene are linked to several peripheral neuropathies including Charcot-Marie-Tooth disease type IB, Dejerrine-Sottas disease, and congenital hypomyelination (Hayasaka et al., 1993; Kulkens et al., 1993; Warner et al., 1996). The adhesion properties o f the extracellular domain o f Po have been suggested to play a role in the initial contact o f glia to the axons and in the compaction of myelin. Po is capable o f homophilic binding when ectopically expressed in transfected cells (Filbin et al., 1990; D ’Urso et al., 1990). It is also reported that when ectopically expressed in carcinoma cells, the host cells could revert to an epithelial phenotype (Doyle et al., 1995). When coated on plastic, Po extracellular domain promotes neurite outgrowth o f DRG neurons (Schneider-Schaulies et al., 1990). Nonetheless, it is still not quite clear how the molecule achieves its adhesive ability and compaction. It is suggested that L2/HNK-1 carbohydrate, as well as protein-protein interactions, mediate homophilic binding (Griffith et al., 1992; Filbin and Tennekoon, 1991, 1993). However, the crystal structure o f Po suggests that Po molecules could form a lattice and hold two opposing membranes together, even without glycosylation (Shapiro et al., 1996). No matter whether it is associated with the polypeptide chains or with the glycans, the adhesion ability o f Po provides an essential force in forming and maintaining the myelin sheaths.
1989). Both Po protein synthesis and mRNA levels rise about 30- to 40 fold from birth to their peak in actively myelinating nerves during the second week and fall to lower levels in adult nerves (Lemke and Axel, 1985; Trapp et al., 1988; Stahl et al., 1990; Baron et al., 1994). Schwann cells require continual positive signals from axons in order to synthesize high levels o f Po (Weinberg and Spencer, 1976, Trapp et al. 1988, Mirsky et al., 1980; Lemke and Chao, 1988; Jessen & Mirsky 1991, Fernandez-Valle et al., 1993, Scherer et al., 1994, Gupta et al., 1993). In other words, the presence or absence o f competent axons markedly influences the expression o f the Po gene (and other myelin protein genes) in Schwann cells. Axotomy is followed by a rapid and profound down-regulation (Le Blanc et al., 1987; Gupta et al., 1988, Trapp et al., 1988). This loss o f phenotype is flexible in that reinnervation is associated with re-expression (Le Blanc et al., 1987; Mitchell et al., 1990). When associated with neurites that are usually not myelinated, the Schwann cells express very low levels o f Po protein (Aguayo et al., 1972; Brunden et al., 1992). Although the Po is largely down regulated when deprived o f signal from axons larger than 1 |j.M in diameter, a basal level o f Po mRNA in this condition has been reported by several groups (Poduslo et al, 1985; LeBlanc et al., 1987; Brunden and Poduslo, 1987; Brunden et al., 1990; Brunden, 1992; Kreider et al., 1988; Burro ni et al., 1988; LeBlanc and Poduslo et al., 1990). Very low levels o f protein are detected in denervated nerves, in Schwann cells in culture, or Schwann cell co-cultured with SCG neurites (Mirsky et al., 1980; Poduslo et al., 1985; Morrison et al., 1991; Brunden et al., 1992), coincident with the presence o f basal levels o f Po mRNA.
Meyer, 1993; Einheber, et al., 1995; Cheng and Mudge, 1996), and the effect is more profound when the cell is in the cell cycle (Morgan et al., 1994). DNA synthesis, when induced by serum or growth factors including PDGF, bFGF, TGF-p at the presence o f cAMP, inhibits the cAMP induced Po level (Morgan, et al, 1991, 1994). A study o f transcriptional elements o f the Po gene shows that cis-acting elements reside within a 500 bp region o f DNA upstream o f the coding sequences region (Lemke et al., 1988; Brown and Lemke, 1997). The promoter fragment does contain a known cAMP response element (Brown and Lemke, 1997), and it alone can drive Schwann cell specific expression o f heterogeneic genes (Messing, 1992, 1994). In vitro, this promoter site can be specifically bound and repressed by Tst-1, a member o f the POU domain gene family (He et al., 1991)
The product o f Po gene is regulated mostly at transcriptional level. Po mRNA and protein expression show a similar pattern during developement, and in denervation the down-regulation o f the protein is accompanied by a down-regulation o f the mRNA (Lemke and Axel, 1985; Stahl et al., 1990). Basal Po mRNA could be expressed when no protein is expressed, during denervation or when the cells are cultured without serum or ascorbic acid. (Poduslo et al, 1985; LeBlanc et al., 1987; Brunden et al., 1990; Kreider et al., 1988; Burroni et al., 1988; LeBlanc and Poduslo et al., 1990, Morrison, 1991). In other word, a translational regulation o f Po gene product exists for the low level expression o f mRNA, when there is minimal axonal contact. Post-translational regulation by catabolism o f Po is also reported (Brunden et al., 1990). When deprived o f axonal contact by denervation, posttranslational oligosaccharide processing o f Po is altered, and the glycoprotein is degraded in lysosomes soon after its formation (Poduslo, 1985; Brunden and Poduslo, 1987).
species o f the mammalian, avian, amphibian, bony fish, and cartilaginous fish (Lemke and Axel, 1985; Sakamoto et al., 1987; Barbu, 1990; Takei and Uyemura, 1993; Stratmann and Jeserich, 1995; Saaveder et al., 1989). The overall homology o f Po-like molecules in chick and trout to rat is 78% and 51%, respectively (Barbu,
1990; Stratmann and Jeserich, 1995). The Po gene is a major component o f the CNS myelin in the fish but during evolution it is excluded from the CNS and replaced by PLP/DM-20 in terrestrial veterbrates (amphibia, reptilia, aves, and mammalia). Two alternatively spliced forms o f trout Po mRNA species have been reported for CNS and PNS respectively, and several mRNA species for the Po gene have been reported in chick (Stratmann and Jeserich, 1995; Barbu, 1990), although other researchers suggest that the detection o f several species o f embryonic chick Po mRNA arises from cross reaction with 28S rRNA (Zhang et al., 1995). Although at the protein level various Po species are different in that a substantial number o f substitutions are found, the overall structural similarity is retained (LeBlanc and Mezei, 1986; Barbu, 1990; Stratmann and Jeserich, 1995). Post-translational modifications seem to be conserved across species (Lemieux et al., 1995).
1.8 The neural crest
The neural crest is a group o f cells which emerges as the neural plate folds to form the neural tube. Before their emigration, these cells are morphologically similar to epithelium-like neural tube cells. Evidence suggestes that neural crest cells are not segregated within the tube and are equipotent to the other cells located more ventrally in the tube (Bronner-Fraser and Fraser, 1989). The premigratory neural crest cells switch their adhesion between cells to more cell-substrate interactions, and exit from the dorsal midline. After their emergence, these cells proliferate and differentiate as they migrate along defined pathways to form seemingly unrelated types o f cells, which include; 1) the neurons and nonneuronal cells o f the sensory, sympathetic, and
T able 1-1 D erivatives o f the neural crest
Classification Cell type
PNS Neurons
Specific tissues or cells Sensory neurons of.
Spinal ganglia
Trigeminal (V) root ganglion, facial (VII) root ganglion,glossopharyngeal (IX) root ganglion, vagal (X) root (jugular ganglion)
Autonomic neurons of: Sympathetic ganglia Parasympathetic ganglia Enteric ganglia
Satellite cells in:
All sensory ganglia, including geniculate (VII), otic (VIII), petrosal (IX), and nodose (X)
Sympathetic and parasympathetic ganglia Enteric ganglia
Schwann cells o f peripheral nerves Nasal and orbitary skeleton
Palate and maxillary skeleton Trabecule
Sphenoid capsule Cranial vault:
Squamosal Frontal Otic capsule Visceral skeleton
Dermis, smooth muscle and adipose tissue of the skin in face and ventral part o f neck Ciliary muscles
Striated muscle o f face and neck
Wall o f large arteries derived from aortic arches Glial cells
Mesectodermal Skeletal tissue derivatives
Other derivatives
Endocrine and paraendocrine cells
Melanocytes
(except endothelial cells )
Tooth papillae (except endothelium o f blood vessels )
Corneal ‘endothelium’ and stromal fibroblasts Meninges in:
Prosencephalon mesecephalon Connective tissue of:
pituitary
lacrymal glands salivary glands
thyroid and parathyroid thymus
Dorsal fm mesenchyme (amphibians and fishes) Carotid body type I cells
Calcitonin-producing cells (C-cells) Adrenal medulla
(adapted from Le Douarin and Smith, 1988)
1.8.1 Experimental approaches
system (Le Douarin 1973). This method is based on the structural differences in the interphase nucleus in these 2 species. Explants could be taken from the donor and grafted into the same or different locations o f the same or different developmental stages o f the host in ovo. This is a useful tool but still was not applied to mammals. Monoclonal antibodies NC-1 and HNK-1 (Vincent et al, 1983; Abo and Balch, 1981) both recognize a carbohydrate epitope on the majority o f migrating neural crest cells (Tucker et al, 1984). By examining whole mount embryo or sections through embryos stained at different stages, pathways o f neural crest migration can be inferred, both in avian and murine species (Bronner-Frasier, 1986; Loring and Erickson, 1987; Erickson et al., 1989). However this approach has several pitfalls. Firsly the antibodies are not entirely specific to the neural crest population, and do not recognize all neural crest cells. Secondly only a static picture o f neural crest migration could be gained this way. Some time later two other methods were adapted to follow cell lineages o f single cells in vivo. One is a retrovirus-mediated gene transfer method, in which a recombinant retrovirus bearing the E. Coli P- Galactosidase (lacZ) gene infects a progenitor and is inherited by its progeny, which can then be identified by a histochemical stain for lacZ (Sanes et al., 1986; Price et al, 1987; Sanes, 1989). The other method is to inject vital dye Dil into a single cell or the lumen o f the neural tube or directly into the neural folds (Serbedzija et al., 1989, 1990, 1991, 1992; Bronner-Frasier and Frasier, 1988, 1989). The dye serves to mark descendants o f the injected cell until the dye fades or is diluted excessively because o f cell proliferation.
1.8.2 The migrating pathways Trunk
Interestingly neural crest cells which migrate ventrally move in a segmental fashion through the rostral half o f each somite, but do not move through the caudal half (Rickmann et al., 1985; Weston, 1963, Erickson et al, 1992), while those that migrate dorsally under skin do so in an unsegmented manner (Serbedzija et al., 1989, 1990, Erickson et al, 1992). This population o f cells appears to migrate much later than those that take the ventral pathway (see Erickson and Goins, 1995) to give rise to melanocytes (Dusahne, 1935, Dorris, 1939, Rawles, 1947; Mayer, 1973).
reports, however, give an idea o f how difficult it is to describe a highly mobile, three dimensional, heterogenic cell population using morphological evidence.
Head
The cranial neural crest cells differ from the trunk crest in several ways. The cranial crest contributes to the formation o f sensory and autonomic ganglia and gives rise to a range o f mesenchymal tissues produced elsewhere by the mesoderm (Le Lievre and LeDouarin, 1975; Noden, 1975). The migrating pathways o f the cranial crest are much more complex. The migrating cranial neural crest in mouse groups into three broad streams from the dorsal aspect o f the neural tube migrating ventrally from the caudal forebrain, midbrain and hindbrain, while in chick and probably also rat only the midbrain and hindbrain give rise to neural crest (Serbedzija et al., 1992). None or very few neural crest cells were found to migrate into the otic vesicle (Le Douarin, 1982; Lumsden et al., 1991; Serbedzija, 1992). Neural crest arising from the level o f forebrain migrates ventrally in a non-segmented, continuous stream through the mesenchyme between the eye and the diencephalon. The neural crest from the midbrain level migrates ventrolaterally through the mesenchyme between the surface o f mesencephalon and the ectoderm (Le Douarin, 1982; Lumsden et al., 1991; Serbedzija, 1992). Cells from mesencephalic and rhomb encephalic levels migrate into their neighboring branchial arches corresponding to the segmented disposition o f the rhomb encephalon. Branchial arches 1,2 and 3 receive crest cells migrating from rhombomeres 2, 4, and 6 respectively, where the cranial nerve entry/exit points
reside, avoiding mesenchymal space lateral to rhombomere 3 and 5 (Lumsden et al., 1991). Enhanced levels o f cell death were found in the dorsal midline o f rhombormere 3 and 5, suggesting that the lack o f migrating crest from these segments is probably due to elimination instead o f lack o f emigrating cells (Lumsden et al.,