Cover Crop Biomass Production Is More Important than Diversity for Weed Suppression
Full text
Figure

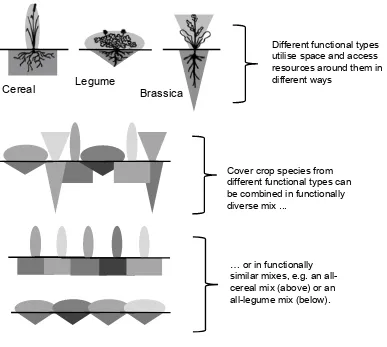
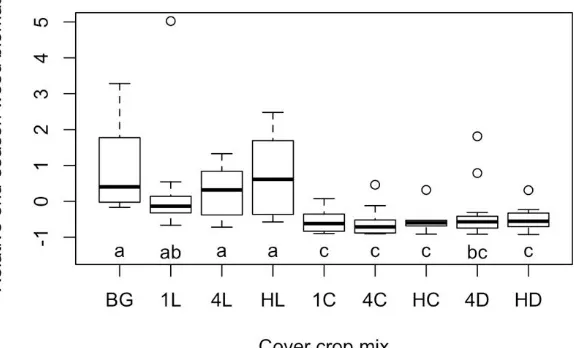
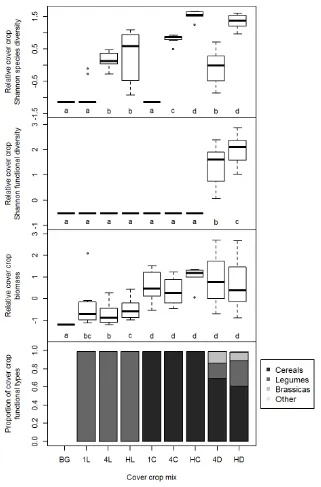


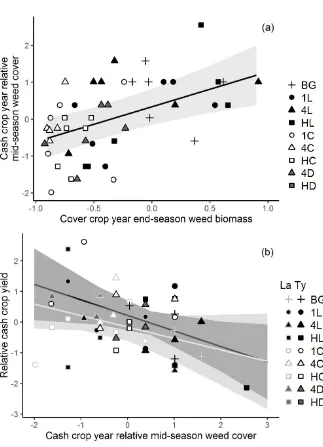
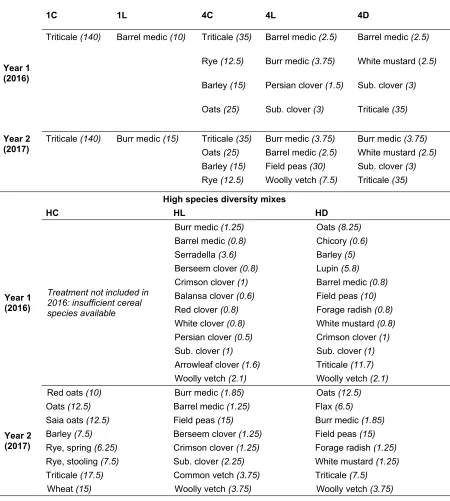
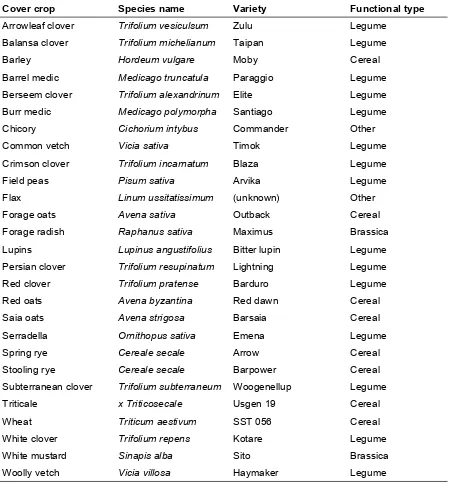
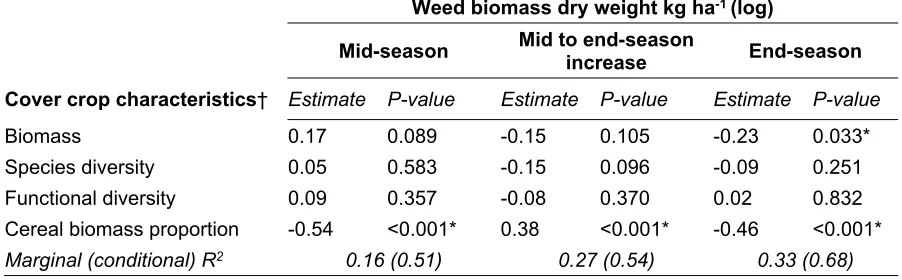
Outline
Related documents
The purpose of this causal comparative quantitative study is to determine the impact that differences in family structure and student attendance, including absence type, had
In case of the two layered concrete beams, the real influence of lightweight concrete (LWC) type in the layered reinforced concrete is observed significantly after increasing
We apply our approach to a decision problem concerning the recommendation of post-operative breast cancer treatment; specifically, whether or not to administer chemotherapy based
Car entrance to the parking facility Public Works Department customer service, Pohjoinen Makasiinikatu 9 Police, Punanotkonkatu 2 Helsinki City Tourist and Convention
The granger causality test examines the causal relationship between pollutant emissions; energy consumption and economic.. growth within a multivariate Johansen’s
[r]
(2002); 3) Assessment of supply chain performance using analysis of the Supply Chain Operation Model (SCOR) approach with performance attributes measured
