J. Exp. Biol. (1964), 41, 113-118 I 1 3 With 2 text-figures
Printed in Great Britain
FROG FAST MUSCLE
II. A METHOD OF MEASURING INTERNAL SERIES COMPLIANCE
BY C. J. PENNYCUICK
Department of Zoology, University of Cambridge
{Received 15 June 1963)
INTRODUCTION
The idea of a series compliance as an integral part of a muscle, following Gasser & Hill (1924), formed an important part of the analysis of Hill (1938). It is regarded as functionally independent of the contractile element, and hence the repository of mechanical energy supplied by that element or by the external load when tension increases. In Hill's (1938) model the speed of shortening, and hence the mechanical power output, of the (fully active) contractile element was held to depend solely on the tension, irrespective of whether this was changing.
In Goodall's (1957) analysis a series compliance is not included as such, the variable extensibility of the entire muscle being regarded as a symptom of activity, without analysing it into components due to active and passive elements. The results of Hill (1953) and Wilkie (1956), however, seem to establish a valid case for the existence of a passive series-elastic element into which energy is fed and from which it can be recovered simultaneously with changes of tension, and there seems little to be gained by abandoning the concept. Clearly those muscles which have tendinous endings must have some series compliance.
Various methods have been used for the estimation of series compliance, in particular the following four:
(1) Comparison of isotonic and isometric tetani (Hill, 1938). A force-velocity relationship is found from isotonic tetani, and it is assumed that at each point on an isometric tetanus the speed of shortening of the contractile component may be deduced from the tension alone. The total (internal + external) series compliance may thus be deduced from the rate of rise of tension. The assumption that the speed of shortening of the contractile component depends only on tension, irrespective of whether the tension is changing, has been queried in the previous paper, and will be shown to be untenable in a subsequent one, which invalidates this method.
(2) Controlled release (Hill, 1950). The muscle is allowed to build up tension in a tetanus isometrically, and is then released at a known speed of shortening, greater than the maximum speed of which the contractile element is capable. The time course of the declining tension is observed, and the shortening of the contractile component allowed for as in (1) above. This method is not essentially different from (1), and suffers from the same objections.
(3) Added external compliance (Macpherson, 1953). A spring of known compliance is connected in series with the muscle, and the time course of tension rise in a tetanus is compared with that obtained without the spring. Once again it is assumed that the
speed of shortening of the contractile component depends only on the tension, when rate of rise of tension is made to vary, and the same objection applies as to (1).
(4) Quick-release method (Wilkie, 1956). The muscle is allowed to build up tension isometrically, and is then released to a lower isotonic tension. There is an initial rapid shortening due to the release of energy stored in the series compliance, followed by much slower shortening at the speed appropriate to the tension. The initial displace-ment step divided by the tension step gives the total series compliance. This method involves no assumptions about the behaviour of the contractile component, except that its speed of shortening shall remain constant from the time the tension reaches the isotonic value.
Wilkie's method has been criticized by Podolsky (i960) on the ground that the con-tractile component takes time to adjust its speed after a change of tension, so causing inaccuracy in estimating the length step following release. This argument is based on an experiment purporting to show that the velocity oscillation following release is generated by the contractile component, and is not due to inertia in the recording system. Length and tension were measured simultaneously during a quick release. When the muscle was replaced by a spring of compliance about equal to that of the series-elastic component, it was found that oscillations of both length and tension occurred, in phase. When the muscle was used there were oscillations of length at a lower frequency than with the spring, but only very small oscillations of tension. Hence (it was argued) the velocity oscillations were not due to resonance caused by the inertia of the lever and the series compliance.
Podolsky does not say whether the spring in the 'substitute' experiment was im-mersed in Ringer's solution, and he does not describe his apparatus in enough detail to enable one to tell whether the mass of the Ringer's solution in the muscle bath loaded the force transducer, a circumstance which is difficult to avoid in systems where length and tension are measured at opposite ends of the muscle. It is thus possible that the force transducer was loaded by a much greater mass when the muscle was tested than with the spring, which might account for the much smaller deflexion during the transient oscillations.
Podolsky's second experiment showed that the period of the length oscillation became shorter as the temperature was increased, the isotonic tension being the same. On increasing the temperature from o to 70 C. the period of the oscillation decreased by a factor of about 3, but this was accompanied by an increase of about four times in the speed of shortening on which the oscillation was superimposed. As only about half a cycle of oscillation is clearly visible, and it is by no means clear where this ends, the validity of this result may be doubted.
On the basis of Podolsky's published evidence his criticism of the validity of Wilkie's method would not therefore appear to be conclusive.
A new method
Frog fast muscle. II 115
change of tension—there is no sudden change either of tension or of speed of shortening of the contractile component.
It is assumed that the mechanical power output of the contractile component is the same just before and just after the transition. Let the isotonic shortening speed be v, the tension F, the power output P, and the total series compliance in the isometric phase C. Then in isotonic contraction
P = Fv, (1) and in isometric contraction
P = CF{dFjdt). (2) If P is the same just before and just after the transition, then
C = vUdF/dt), (3) where v is measured just before the transition and dFjdt just after, or vice versa.
MATERIAL AND METHODS
The material was the fast component of the iliofibularis of the frog, set up as described in the previous paper.
As long as the contractile component is producing a reasonable amount of power it does not matter at what stage of contraction the observation is made, since the only assumption inherent in the method is that v and dFjdt are observed so close together in time that the power output will be, for practical purposes, the same. The method of stimulation chosen was a tetanus at 40/sec. applied to the nerve, ensuring that the power would be large.
Length and tension were recorded, and the records were afterwards differentiated by drawing tangents at the transitions. If transitions in both directions could be recorded on the same trace estimates of compliance were obtained at two different lengths at the same tension (the isotonic tension). Examples of recordings are shown
n Fig. 1.
The dangers of determining the slopes of curves by drawing tangents have been referred to in the previous paper. In the present case the observations were made during a tetanus, where the slopes were not changing rapidly, the most favourable conditions for the procedure.
RESULTS
The method allows the compliance to be estimated at two lengths for each observa-tion, and this can be repeated at a series of different tensions. The compliance observed, after allowing for the compliance of the transducer and connexions, is of the same general order as that found in previous determinations, viz. about 3-5 % of the muscle length for the maximum isometric tension which the muscle can develop.
Variation with length and tension
If Cx denotes the compliance determined at the transition from isometric to isotonic contraction, C2 that found from the reverse transition, and if the muscle is allowed to
shorten 4-6 % of its length in between, then Cx is found to be much more strongly
dependent on tension than Cs. In this case Cx was determined at the length set by 5
g.wt., so that C2 was measured at a length shorter than that corresponding to zero
passive tension.
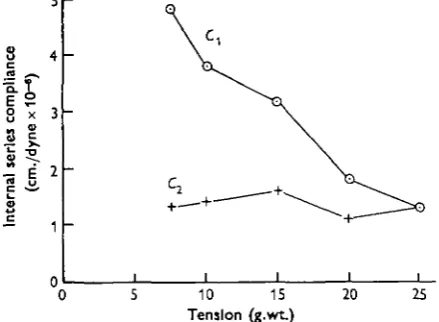
Fig. 2 shows the averages of two experiments in which Cx and C2 were estimated at
5 tensions between 7-5 and 25 g.wt. It can be seen that whereas C1 decreases from nearly 5 x icr6 cm./dyne at 7-5 g.wt. to about 1-3 x icr6 cm./dyne at 25 g.wt., Ct is nearly constant in the region of the latter figure throughout the tension range.
0-1 sec
Fig. 1. (I) Uppertrace: length, shortening downwards. Lower trace: tension, increasingupwards. An estimate of total series compliance is given by the ratio of speed to rate of change of tension at the transitions from isometric to isotonic contraction and back again. Stimulus: tetanus at 40 impulses/sec, applied to nerve. Isotonic tension in g.wt.: a, 7-5 ; b, 10; c, 15 ; d, 20; e, 25. (II) (a,c)As in (I): Tensions 15, 35 g.wt. (6,d) Wilkie's method, same tension. Compliance estimated by dividing length step by tension step afer release, to peak of first overshoot in each case. Scale: *, 10 msec./div.; y, 3-7 g.wt./div.; 0-023 cm./div.
This effect was not a spurious one due to a difference in the compliance of the long and short lever stops on the apparatus. To check this the leverwas held against the long stop by the elastic, and was then gradually loaded with weights in place of the muscle, its movement being observed through a microscope fitted with a micrometer eyepiece. The compliance both just before lifting off the long stop and just after resting on the short stop was found to be less than io~7 cm./dyne.
Frog fast muscle. II 117
initial isotonic velocity would be too high, giving too high a compliance. With increasing tension Cx decreases and approaches C2, as would be expected on this explanation. C2 is therefore regarded as a more reliable measure of series compliance than Cj.
I
Q. 1
E ? 8 x 3
S •5 Eg 2
I
10 15 Tension (g.wt.)
[image:5.451.119.339.121.282.2]20
Fig. 2. Ci is compliance measured at transition from isometric to isotonic shortening, C, is compliance measured at the reverse transition. Average of two experiments on different muscles, showing d much more strongly dependent on tension than CV
This conclusion was checked by fitting a magnetic release to the lever, which was used on alternate records to lock the lever for the first 180 msec, of the tetanus and then released. The technique resembled Wilkie's (1956) method, except that, as both length and tension were recorded, the steps on each were measured to the peak of the first overshoot, instead of the backward extrapolation of the length record used by Wilkie.
Averaging the results for 10, 15, 20 and 25 g.wt., the values obtained by the two methods for the total series compliance were: C1; 2-26 x io~* cm./dyne; C2,1-15 x io~*
cm./dyne; Wilkie's method, 1-25 x io~* cm./dyne.
The shortening allowed between observations of C1 and C2 was here 0-25 cm., or
10% of the muscle length.
The value of Cs agrees well with that obtained by Wilkie's method, while that for
Cx is i-8 times as great. The method appears to be a reasonable one provided care is
taken to ensure that the tension which might be expected to remain in the parallel elastic structures at the transition is small compared to the nominal tension.
SUMMARY
1. Methods which have been used in the past for estimating internal series com-pliance are discussed.
3. The method is found to agree with Wilkie's quick-release method, provided parallel elastic structures are not carrying significant tension at the transition.
It is a great pleasure to thank my supervisor, Prof. J. W. S. Pringle, F.R.S., for his help and encouragement during this work. I am also deeply indebted to Dr K. E. Machin, in particular, for discussion and advice, and to Messrs J. A. Popple and D. M. Unwin for assistance with the design and construction of apparatus. The work was carried out under a D.S.I.R. Research Studentship, and I am most grateful to that organization for their support.
REFERENCES
GASSBR.H. S . & H I L L A.V. (1924). The dynamics of muscular contraction. Proe.Roy. Soc. 8,96,398-437. GOODALL, M. C. (1957). Kinetics of muscular contraction. 1. Yale J. Biol. Med. 30, 224-43.
HILL, A. V. (1938). The heat ofshortening and dynamic constants of muscle. Proe.Roy. Soc. 8,136,136-95. HILL, A. V. (1950). The series elastic component of muscle. Proc. Roy. Soc. B, 137, 273-80. HILL, A. V. (1953). The mechanics of active muscle. Proc. Roy. Soc. B, 141, 104-17.
MACPHERSON, L. (1953). A method of determining the force-velocity relation of muscle from two isometric contractions. J. Pkysiol. iaa, 173-7.
PODOLSKY, R. J. (i960). Kinetics of muscular contraction: the approach to the steady state. Nature,
Lond., 188, 666-8.