[ 9 2 ]
RESPIRATION RATES OF WORKER HONEYBEES OF
DIFFERENT AGES AND AT DIFFERENT TEMPERATURES
BY M. DELIA ALLEN
Bee Research Department, North of Scotland College of Agriculture, Maruchal College, Aberdeen
(Received i August 1958)
INTRODUCTION
Since Rosch (1925) published his results on the ages of the worker honeybees {Apis
mellifera L.) performing various duties in the colony, considerable interest has been
focused on this problem. While it now seems clear that there is no rigid demarcation between the ages of adult workers engaged on different activities—for one bee may perform several different duties on the same day (Lindauer, 1952)—it nevertheless appears that age is not without influence in this respect, as for example in the feeding of the brood (Perepelova, 1928), the feeding of the queen (Allen, 1955), foraging (Ribbands, 1952) and 'shaking' (Allen, 1958). In view of the present state of some uncertainty on the extent of this influence it seemed of interest to examine the metabolic rates of workers of different ages, as reflected by their respiration rates, with the aim of discovering whether any marked differences existed which might in turn be related to the problems of behaviour.
With many animals, especially mammals, it is apparently true that in general the respiration rate falls as the animal becomes older and also as it grows larger, though this is no by means always the case, as for example in a number of invertebrates (reviews by Heilbrunn, 1952, Ch. 20; Prosser et al. 1950, Ch. 8). Also, in probably the majority of invertebrates and poikilothermic vertebrates the respiration rate increases with temperature, within the vital limits, although the value of the tem-perature coefficient does not necessarily remain constant at different temtem-peratures. Honeybees and some other winged insects have, however, the ability to raise their temperature appreciably above that of the environment (Pirsch, 1923; Himmer, 1932) and their metabolic rate is, therefore, likely to bear a complex relationship to environmental temperature. Bearing this in mind it seemed of some interest to relate the oxygen consumptions of stationary worker bees not only to age, but also to external temperature over the range between the point at which the insects were just in chill coma and the estimated upper lethal temperature (approximately
7-46° C ) .
METHODS
Respiration rates of worker honeybees 93
a separate match-box and fed with honey for 10 min., after which it was picked up by the wings and was put head-first into a copper gauze cylinder approximately J in. in diameter and 1 in. long, which had one end sealed off. With blunt forceps the bee was gently pushed down the cylinder until its head was just touching the end, and then a plug of cotton wool was inserted so that it rested lightly against the tip of the abdomen and closed the cylinder. As a result only slight movements were possible. Throughout the caging operation there was a danger that the bee might use its sting and in so doing be damaged by having it dragged out of the body, but with practice this did not occur frequently. However, when any bee was damaged in this way (or in any other) it was discarded.
It was appreciated that by feeding the bees before the experiments the respiration rates would probably be raised somewhat above those of fasting bees, but since some bees (particularly incoming foragers) would undoubtedly have had some honey or nectar in their honey stomachs when they were removed from the hive the only means of achieving any uniformity in this respect seemed to be by supplying food to all bees prior to the measurement of their respiration rates.
It was in most instances found convenient to measure the respiration rates of two age-groups simultaneously, using six (or in a few instances, five) bees of each age; immediately after the bees were caged each one was placed in a separate Warburg constant volume respirometer with a capacity of between 24 and 29 ml., containing 0 2 ml. 5% KOH solution in the centre well, and the apparatus was then allowed to equilibrate at the experimental temperature for 15 min. before readings were commenced. The preparation time, by which is meant the duration of the period between the removal of the bees from the hive and the end of equilibration, was in all cases approximately 1 hr. All the experiments were carried out between 5 April and 12 September, when the colonies were in the active summer condition.
The respirometers were not shaken during the experiments because normal gaseous diffusion was sufficient to ensure a constant supply of oxygen to the bees. The volumes both of the bee (approximately 100 /A.) and of the copper and cotton wool of the cage (approximately 100-160 fil.) were not taken into account in the calculations of the flask constants, in view of the fact that they were relatively constant and in any case would have caused a reduction of less than 1 % in the final result, although their accurate determination would have added considerably to the labour involved.
thorax and abdomen (minus the last abdominal segment) were then weighed. It was not practicable to determine the weights of the cleaned alimentary canals of each bee used in the experiments, but an independent investigation showed that for a bee weighing 70 mg. (approximately the mean weight) after removal of the alimentary canal, its average weight was 9 mg. Accordingly, an addition of 13% was made to the weights of all the bees used.
After a number of preliminary experiments it was decided that there was a possi-bility of oxygen shortage influencing the results after about one-eighth of the total volume of oxygen had been consumed. In all, seventy-nine groups of bees (com-prising 463 individuals) were used, and in fifty-four of these groups this point was not reached during the first hour. Here the final readings were taken after 60 min., but in some experiments the respiration rates were more rapid and then the final readings were normally taken after 30 min. Because of this and for other reasons twenty-three groups of bees were recorded over this shorter period, and two groups over 20 min. only (Table 1).
It is evident that if the individual respiration rates were in the process of increasing or decreasing in those experiments where respiration rates were measured over less than 1 hr., the calculated volume of oxygen consumed in one hour would be res-pectively either lower or higher than the volume which would have been consumed had it been possible to continue the experiments for the full hour; nevertheless, after an examination of the individual respiration rate curves it was considered preferable to use the values based on the greater numbers of readings rather than be limited to the results obtained after 30 min. in all cases, both because of the greater accuracy of the final result (particularly at low rates of respiration) and also in view of the fact that the causes of the increasing and decreasing respiration rates were not sufficiently well known.
RESULTS
Using groups of bees ranging from o days old to 22-33 ^ Y8 ° ^ the relationships between the ages of the bees and their oxygen consumption per mg. live weight per hour (QOl) were determined at 7, 12, 17, 22, 27, 32, 37, 40, 42 and 46°C.
Figs. 1 and 2 are given as examples of the results obtained.
The values of QOf (see above) of individual bees at each particular age and
tem-perature usually had a markedly skew distribution, that is, far more readings fell on the lower side of the mean than on the higher. In order to work with data having more nearly a normal distribution it was found convenient to use a logarithmic transformation when comparing age and temperature effects, and in Figs. 1 and 2 it is the logarithms (to base 10) of the individual Qot values and the means of these
Respiration rates of worker honeybees
95
Table i. Details of ages of bees used in the experiments
(Figures in brackets indicate replication of groups.)
Temperature (*C.) 7 12 17 2 2 27 37 4° 42 46
Length of time over which respiration rates were determined
60 min.
Newly emerged bees ( x j ) 18 hr. old bees
2-day-old bees 12-day-old bees 19-day-old bees 3 3-day-old bees Newly emerged bees
18 hr. old bees 2-day-old bees 6-day-old bees 15-day-old bees 20-day-old bees 32-day-old bees Newly emerged bee»
18 hr. old bees 2-day-old bees Newly emerged bees
18 hr. old bees 7-day-old bees Newly emerged bees ( x 4)
18 hr. old bees ( x i ) 2-day-old bees ( x 2) 4-day-old bees 6-day-old bees 10-day-old bees ( x 2) 17-day-old bees 19-day-old bees 26-day-old bees 33-day-old bees Newly emerged bees
7-day-old bees 14-day-old bees 20-day-old bees 23-day-old bees 30-day-old bees Newly emerged bees ( x 3)
5-day-old bees 8-day-old bees 13-day-old bees Newly emerged bees
8-day-old bees 15-day-old bees 22-day-old bees Newly emerged bees 17-day-old bees — 30 min. — 9-day-old bees 17-day-old bees 2-day-old bees 18-day-old bees 32-day-old bees —
18 hr. old bees
20 min. — 21-day-old bees 27-day-old bees — — —
2-day-old bees '•
18 hr. old bees 2-day-old bees
—
22-day-old bees 29-day-old bees
— ' —
18 hr. old bees — 2-day-old bees
4-day-old bees 8-day-old bees 10-day-old bees 31-day-old bees Newly emerged bees 18 hr. old bees
—
The chief interest of the respiration rates given here lies not so much in their actual numerical values as in the relative differences at different ages and tem-peratures, for it is not claimed that they are a measure of basal metabolic rates of the bees concerned but simply that they reflect the metabolic rates under the experi-mental conditions. Consequently, the results have been replotted in Fig. 3 to show
1-6 1-4 1-2 1-0 0-8 0-6 =£. 0-4
r
(? 0-2 | 0- 0 - 2
- 0 - 4
- 0 - 6
- 0 - 8 <
( -o -O/
7
I
-1-Og-1-2 C)
o / o / / / ' 0 o 1 1 5 10 o o // o o 1 15 Age 0 8o ^ ^
o ^ ^
^s^ 0
o
o o
1 1 1
20 25 30 35 (days) JZ M "a.
3
1-6 1-4 1-2 1-0 08 06 0-4 0-2 ( 0<- 0 - 2
- 0 - 4
- 0 - 6
- 0 - 8
- 1 0
1-2 -o %
-3 5 1 0
8
o
8 8
o
1 1 1
15 20 25 Age (d»y$)
4
o
i
30 3
Fig. i. Oxygen uptake of adult workers at 120 C. Fig. 2. Oxygen uptake of adult workers at 32° C.
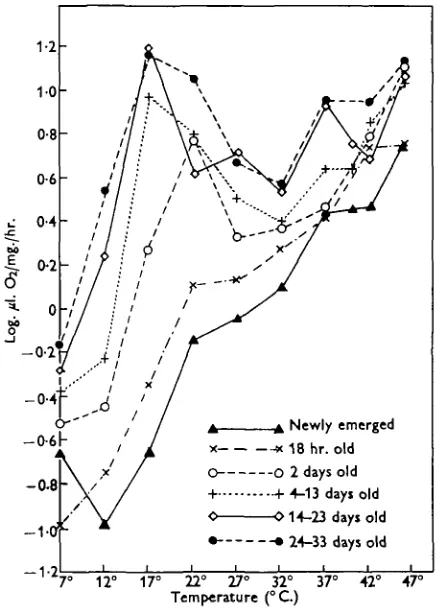
the influence of both temperature and age on the respiration rates. The age groups here considered are as follows: newly emerged, 18 hr., 2 days, 4-13 days, 14-23 days, and 24-33 days. The means used (as in Figs. 1 and 2) were calculated from the log10 respiratory values of all the individuals falling into each of the above
categories.
Respiration rates of worker honeybees 97
Secondly, the individual curves do not all follow identical paths. The newly emerged and 18 hr. bees have basically a linear increase in respiration rate with temperature when plotted in this way (apart from a few fluctuations), as might be expected in a poikilothermic animal. At 2 days old, however, the position is less straightforward, since there is a peak at 220 C. in addition to the peak at 460 C. This earlier peak is even more pronounced in the three oldest groups, where it occurs at i 7 ° C , reaching a height approximately the same as that at 46°C. Between the two peaks is a trough with a minimum value at about 32°C.
x x 18 hr. old
O O 2 days old + •+ 4-13 dayj old
14-23 days old
• • 24-33 days old
[image:6.451.117.338.185.491.2]7° 12° 17° 22° 27° 32° 37° 42° 47° Temperature ( ° C )
Fig. 3. Effect of age and temperature on oxygen uptake of adult workers (means).
The main trends shown in Fig. 3 are very clear, but it is perhaps informative also to consider the individual results briefly. Usually the readings for each bee recorded at 10 min. intervals lay closely on a straight line and sudden changes in respiration rate, which would have indicated fluctuations in activity, occurred only in six instances; of these, four were found at a temperature of 460 C , which was lethal after about 1 hr. Sometimes, however, a tendency for respiration rates either to increase or decrease with time was apparent; the details of these changes have been discussed by Allen (1958), but broadly speaking they amounted to a decrease with time at the lowest temperatures (7 and 120 C ; possibly at 170 C ) , and an increase
at temperatures approaching the upper lethal point (42 and 460 C), although not all bees at these temperatures showed any alteration in rate over the period studied. At 22, 27, 32, 37 and 400 C. there was no consistent indication of a curved relation-ship with time.
It has been stated already that there was little evidence of fluctuating muscular activity during the experiments, but no visual estimate could be made of the level of activity while the respiration rates were being measured because the bees were not clearly visible while in the gauze cages. However, in order to make some assessment of the probable activity, observations were made on groups often foraging and ten newly emerged bees confined in glass tubes by pads of cotton wool, so placed that the bees could make only limited movements. These were maintained at temperatures of 7, 12, 17, 27, 37 and 460 C. respectively. Both age groups were completely motionless at 7° C. but the foraging bees made slight movements at 120 C , whereas the young bees were in a state of chill coma as before. At 170 C. the foragers were apparently capable of making normal movements, but the young bees reached the corresponding state only at 22° C. At 27 and 370 C. both groups moved readily if at all disturbed, but if left undisturbed both tended to remain stationary. At 46° C. both for a time showed marked respiratory movements of the abdomen and made rather spasmodic movements of the legs, though later the level of activity gradually decreased, until after 60 min. one newly emerged bee and six foragers were found to be dead. It thus appears that the foragers may be more sus-ceptible than the young bees to high temperatures, whereas the young bees chill at low temperatures more readily than the old bees.
DISCUSSION
It has been shown that the respiration rates of the two youngest groups of bees studied (0-18 hr. old) had a simple relationship with temperature. All the groups of older bees, however, had two peaks, one in the region of the upper lethal tem-perature and the other at a clearly suboptimal temtem-perature. The lowest point of the trough lay between at about 320 C , which is the temperature found towards the centre of the brood area in a healthy colony. It is difficult or impossible to define the optimum temperature for adult bees, but at 32° C. the workers carry out many of their normal activities under natural conditions, and for this reason it may be assumed that at this temperature the respiration rates are those corresponding to the rates occurring in the brood area of the hive. For convenience, temperatures in the region of 32° C. will be referred to subsequently as 'optimal temperatures'. From this assumption and from the evidence supplied by the two youngest groups in Fig. 3, it would seem that the first of the two peaks in the older age groups can be regarded as an elevation of the ' expected' rate, due to factors not normally operating in truly poikilothermic animals. It is suggested that the cause is, in fact, that the bees are exerting some degree of temperature regulation by which the metabolic processes are speeded up when the environmental temperature is too low and the insects are consequently in danger of being rendered inactive.
notice-Respiration rates of worker honeybees 99
ably higher in the newly emerged bees than in the foragers, and it is noteworthy that in the three oldest groups studied (4-33 days) the first peak is found at a temperature thought to be the lowest at which normal movements are possible. Furthermore, the 2-day-old bees, which are likely to chill at a temperature inter-mediate between that causing chilling in the youngest and in the older bees, showed this first peak at 22° C. instead of 170 C , presumably again indicating that it occurred as soon as the temperature was high enough to permit normal movement. The results also suggest that the movements (if, indeed, the increased respiration were due to greater muscular activity) decreased in vigour when the optimal temperatures were reached, since the oxygen consumption at these temperatures was lower than pre-viously ; whereas if the same degree of activity had been maintained throughout, oxygen consumption should have risen with increasing temperature.
It has been found previously by a number of workers that individual bees, as opposed to a group, can raise their internal temperature above that of the environ-ment when it i3 at suboptimal levels and lower it at high temperatures as, for example, Pirsch (1923) has shown. In addition, Himmer (1932) reported that this temperature regulation in the honeybee was influenced by age. He demonstrated that 1-2-day-old bees had only a very limited ability to raise their temperature above that of the environment, while 'house' bees of 3-15 days were much better developed in this respect, and guard bees and foragers of more than 20 days old could raise their temperature even higher above that of the surrounding air. The results shown in Fig. 3 are in accordance with these findings, since the highest values for oxygen consumption at suboptimal temperatures were recorded in the two oldest groups of bees (14-33 days), with a steadily declining rate as age de-creased. Also in agreement with Himmer's work are the lack of evidence of tem-perature regulation in the two youngest groups (0-18 hr.), the peak at 220 C. in the 2-day-old group, and the earlier, higher peaks in the 4-33 day groups at 170 C.; all of which very probably indicate higher internal temperatures with increasing age at these suboptimal environmental temperatures.
Expressed in another way, this early peak and subsequent depression in the respiration rates of what would be the majority of the bees in a colony is almost certainly related to the heat regulation of the cluster. At temperatures below the optimal cluster temperature some means of heat production is necessary and, whatever the exact mechanism, this can be achieved in practical terms only by increased consumption of oxygen. At or about the optimal temperature, however, no such increased rate of consumption is required. The quieter behaviour of the bees observed in the glass tubes when at 27 and 370 C. than when at 170 C. is also broadly in keeping with this view.
It is difficult to compare the numerical values for oxygen consumption obtained in these experiments with those found by the few other workers who have made similar measurements on bees at various temperatures (Parhon, 1909; Steidle, quoted by Zander, 1921; Kosmin, Alpatov and Resnitschenko, 1932) because they have all used different methods of calculating the final QOi values, and this materially
affects the results. Furthermore, Parhon used relatively large groups of bees free
to move about, whereas Kosmin et al. (1932) worked with individual bees and noted the amount of movement. Steidle also used individual bees but did not disclose their degree of activity. The results of these workers have been further discussed elsewhere (Allen, 1958), but here it is sufficient to mention that there is some indication that the older bees in the present experiments had somewhat similar respiration rates to the foraging bees of Kosmin et al. (1932) which were described as 'moving slowly on floor of vessel'.
Finally, returning to the question of whether behavioural changes at different ages coincide with changes in metabolic rates, it can be stated that there is some evidence that this is actually the case. For example, the present results suggest that the older bees are capable of raising their internal temperature above that of the environment when the latter is at a markedly suboptimal level, whereas the younger bees show no sign of having such an ability, and it thus appears that the responsi-bility for raising the temperature of the colony in cold weather must lie with the older bees. Similarly, an appreciable difference has been demonstrated between the rates of oxygen uptake of young bees and that of bees of foraging age, and while this could possibly have been due to a greater degree of movement by the older bees, the fact that the same difference was found at 70 C. when all ages were presumably quite stationary would indicate that more basic causes were responsible.
SUMMARY
1. The respiration rates of individual adult worker bees confined in small gauze cages were determined over a range of temperatures between 7 and 46° C. The respiration rates usually remained constant over a period of 1 hr., although at 7, 120 C , and possibly at 170 C. there were indications of a decrease with time, and at 42 and 460 C. indications of an increase with time. Fluctuations which would have reflected variable activity during any of the experiments were rare.
2. At each temperature studied the respiration rates increased progressively from the youngest adults (newly emerged) to the oldest measured (24-33 days). The increase was particularly marked during the first few days of adult life.
3. The respiration rates of newly emerged and 18 hr. old adults increased pro-gressively between 7 and 460 C, but 4-33-day-old bees showed two peaks (at 17 and 460 C.) with a depression having a minimum value at 32° C. lying between. 2-day-old bees had similar peaks at 22 and 460 C.
4. It is suggested that the first peak in the respiration rate of the older bees occurred at the lowest temperature at which free movements were possible, and that it indicated an increased body temperature maintained by the bees to combat the effects of the low environmental temperature. The lack of this first peak in the case of the youngest bees is thought to mean that they were unable to raise their temperature above that of the environment. The information obtainable from the literature supported these conclusions; their implications in relation to the regulation of temperature within the colony are discussed.
Respiration rates of worker honeybees 101
REFERENCES
ALLEN, M. DELIA (1955). Observations on honeybees attending their queen. Brit. J. Aram. Behav. 3,66-9.
ALLEN, M. DELIA (1958). A study of some activities of the honeybee. Ph.D. Thesis, University of London.
HAYDAK, M. (1934). Changes in total nitrogen content during the life of the imago of the worker honeybee. J. Agric. Res. 49, 21-8.
HEILBRUNN, L. V. (1952). An Outline of General Physiology, 3rd edition. Philadelphia: Saunders. HIMMER, A. (1932). Die Temperaturverhaltnisse bei den sozialen Hymenopteren. Biol. Rev. 7,
224-53.
KOSMIN, N. P., ALPATOV, W. W. & RESNITSCHENKO, M. S. (1932). Zur Erkenntnis des Gaswechsels und des Energieverbrauchs der Biene in Beziehung zu deren Aktivitflt. Z. vergl. Pkysiol. 17,
408-22.
LINDAUER, M. (1952). Ein Beitrag zur Frage der Arbeitsteilung im Bienenstaat, Z. vergl. Physiol. 34, 299-345. (Transl. Bee World 34, 63-73, 85-90.)
PARHON, M. (1909). Les echanges nutritifa chez les abeilles. Ann. Sci. Nat. Zool. 9, 1-58. PEREPELOVA, L. I. (1928). [The nurse bees.] Opuit. Pas. 551-7.
PIRSCH, G. B. (1923). Studies on the temperature of individual insects, with special reference to the honeybee. J. Agric. Res. 24, 275-87.
PROSSER, C. L., BISHOP, D. W., BROWN, F. A., JAHN, T. L. & WULFF, V. J. (1950). Comparative Animal Physiology. Philadelphia: Saunders.
RIBBANDS, C. R. (1952). Division of labour in the honeybee community. Proc. Roy. Soc. B, 140,
32-43-ROSCH, G. A. (1925). Untersuchungen iiber die Arbeitsteilung im Bienenstaat. I. Die TStigkeiten im normalen Beinenstaate und ihre Beziehungen zum Alter der Arbeitsbeinen. Z. vergl. Physiol. a,