1
Spatial Dynamics of Optimal Management in Bioeconomic Systems
David Aadland, Charles Sims and David Finnoff*
April 2013
Abstract. We develop a methodology to evaluate optimal management in a spatially and temporally dynamic bioeconomic system. The method involves standard techniques from the macroeconomics literature to calculate approximately optimal linear decision rules. Iterations between the decision rules and the biological system produce optimal transition paths over space and time. We then apply the methodology to forest management over a spatial grid where a pest insect (mountain pine beetles) preys on trees that provide a wide array of ecosystem services. The method is sufficiently general to be applicable to a wide range of spatially and temporally dynamic economic systems.
Keywords: dynamic systems, spatial models, bioeconomics, migration, predator-prey models JEL Codes: C63, D62, Q23, Q57
*
Aadland ([email protected]) and Finnoff ([email protected]) are associate professors in the Department of
2 1. Introduction
Spatial dynamic problems have gained increasing attention in the economics literature, particularly in the areas of international trade policy (Krugman, 1991), urban growth (Fujita, 1976), labor migration (Tabuchi and Thisse, 2002), and environmental policy (Fredriksson and Millimet, 2002). Research in the optimal management of ecosystems and natural resources has also started to integrate space into dynamic control problems. Research in this area has focused on metapopulation models with native species migration (e.g., Brown and Roughgarden, 1997; Bulte and van Kooten, 1999; Sanchirico and Wilen, 1999; Smith and Wilen, 2003; Horan, et al., 2005; Bhat and Huffaker, 2007; Costello and Polasky, 2008), invasive species (Albers, et al., 2010; Sanchirico, et al., 2010; Epanchin-Niell and Wilen, 2012), forest ecosystem management (Rosie, 1990; Swallow and Wear, 1993; Hof, et al., 1994; Albers, 1996; Swallow, et al., 1997; Konoshima, et al., 2008), and general institutional and policy design issues (Swallow, et al., 1990; Sanchirico and Wilen, 2005; Brock and Xepapadeas, 2010; Kaffine and Costello, 2011).
Combining space and time in optimal control problems creates significant degrees of
analytical and computational complexity. To facilitate a solution, researchers have restricted the dimension of the problem by limiting the degree of human behavior, choice variables, number of species, number of patches, number of time periods, migration patterns, or interdependency of spatial patches. For example, Smith, Sanchirico and Wilen (2009) state: “A key feature of virtually all resource models that incorporate both space and time is that spatial characteristics are described by state variables that are indexed by space, but not functionally interdependent over space.”
3
2002), we develop an approximately optimal linear management rule under rational expectations that fully incorporates temporal and spatial dynamics.1 The management rule is used in
conjunction with a nonlinear system of states to simulate optimal paths across space and time. This procedure allows the solution of continuous control problems that are functionally
interdependent over a large spatial grid. Another advantage of this approach is its ability to accommodate multiple state variables over a large spatial domain. This is an essential feature in many natural resource and environmental applications where migration or dispersal patterns are governed by species interactions such as invasive species, agricultural pests, fisheries, and epidemiology. To our knowledge, this is the first time optimal management over space has been considered in a system with multiple state variables.
To highlight the power of this approach, we apply it to the case of a pest species (mountain pine beetle) spreading across a heterogeneous landscape in order to prey on a valuable natural resource (trees). In addition to the methodological contribution, our research highlights several interesting features of managing pest outbreaks. First, pest migration and optimal resource harvesting result in a complex spatial configuration of pest and natural resource densities. This occurs because proximity to a habitat boundary creates differential pest risk across the landscape grid. Second, there exists a fundamental tradeoff between resource harvesting and insect
epidemics. While increased resource harvesting mitigates the severity of pest outbreaks (Sims, et al., 2010), it has the unintended effect of increasing the rate of pest spread over space
(Konoshima, et al., 2008). The intuition is straightforward. When pests disperse across the landscape in a density-dependent fashion, increased resource harvesting encourages pests to
1
4
migrate to areas with more abundant resources (prey). Third, local resource harvesting generates two opposing spatial externalities. The reproduction externality implies that local producers will under-harvest as they fail to recognize how harvesting decreases neighboring pest populations. The migration externality implies that local producers will over-harvest to encourage pest migration off their management unit. In a calibrated version of our model these two spatial externalities approximately offset.
2. Modeling Framework
Consider a spatially explicit bioeconomic system where the region is divided into a grid with columns indexed by and rows indexed by . Each cell on the grid is
represented by an pair with the neighboring cell of indexed by . Time is discrete and indexed by . The biological system is represented by a vector of state variables, . A subset of these variables is subject to dispersal over space governed by vectors and . Each element of the dispersal vector
gives the fraction of the associated stock in the neighbor of that disperses into in period , where is the set of all neighbors. Each element of the vector gives the fraction of the associated stock in that remains in . To capture density-dependent dispersal, the dispersal weights are a function of .2 The dispersal weights must also satisfy the adding-up constraint:
∑ (1)
5
for , which is consistent with a closed bioeconomic system (i.e., no dispersal enters or exits the system). State variables that do not disperse are assigned the value .
State variables provide value to the economic system in situ or from their harvest. These values are commonly referred to as ecosystem services (Millenium Ecosystem Assessment, 2005). The harvest of the state variable on cell in period is given by . State variables that yield no economic value from their harvest are assigned . Harvesting, depreciation, reproduction, and dispersal combine to characterize the dynamic process for the state variables. The state variables for follow a non-linear first-order process:
( ) ∑ (2)
where is the net growth function for the state variable. To capture unknown
environmental shocks to the biological system (e.g., changes in temperature or precipitation), may be subject to a random disturbance term .
Local planners choose harvests to maximize the stream of utility, , from the ecosystem services derived by local resources, . Local planner chooses a harvest vector in each period to maximize:
∑ ( )
where is the expectation operator conditional on time information, and is the discount factor. The maximization problem is subject to initial values, spatial boundary
6
constrained dynamic optimization problem that can be solved using well-known techniques such as the substitution or Lagrangian methods. However, when resources disperse across space, the optimization problem becomes more difficult and potentially intractable. We propose two simplifications to the optimization problem that will permit solutions for large grids and multiple state variables.
First, we limit the degree to which a local planner accounts for the impact of her current harvesting on her future utility through resource migration. With density-dependent dispersal, harvesting influences migration. These migration effects can be classified as either endogenous or exogenous. Endogenous migration captures 1) how the local planner’s current harvesting choice influences resource migration and 2) how those effects on migration impact her own future utility. Over time the impact of the local harvesting choice in period will work its way across the entire grid as the resource migrates. For tractability, we assume the local planner only accounts for how her harvesting choice affects future utility through migration to her neighbor and back. The local planner therefore ignores how current harvesting choices influence future utility through multi-cell migration (e.g., migration to a neighbor, then to the neighbor’s neighbor, and eventually back to the local cell). It is important to recognize that our
simplification only applies to future impacts caused directly by own harvesting choices. By contrast, the local planner fully considers exogenous migration effects that are outside her
7
Second, we use standard methods from the macroeconomics literature (Clarida, et al., 2000; Farmer, 2002) to develop a linear approximately optimal management rule under rational expectations that incorporates the full set of temporal and spatial dynamics across the region. Similar methods are regularly used in macroeconomics to simulate and study models of the business cycle and optimal policy. Consider a first-order Taylor series approximation of the bioeconomic system around its steady state:
[̃ ̃ ] [̃ ̃
], (4)
where ( ) is the vector of all state (choice) variables at each location on the grid in period , and is a square matrix of coefficients.3 The tilde ( ) above the variables indicates deviation from the respective steady state (e.g., ̃ ).4 As described in Blanchard and Kahn (1980), the system exhibits saddle-path stability if the number of
eigenvalues of inside the unit circle equals the number of variables in ̃ . Assuming the system satisfies the saddle-path stability condition, this implies linear decision (harvest) rules of the form:
Notice that the harvesting rule in equation (5) potentially depends on the entire set of
contemporaneous stocks across the landscape grid. Given initial values for these state variables, equation (5) then determines the approximately optimal harvest levels at every point on the grid.
3 Linearizing the Euler equations from (3) and biological constraints is equivalent to approximating the original management problem as a linear-quadratic (LQ) problem (Cooley and Prescott, 1995; Brock and Xepapadeas, 2008).
8
Once these values are known, the laws of motion for the state variables in equation (2) give next period’s state variables, next period’s state variables then give next period’s harvest levels, next period’s harvests then imply next period’s state variables, and so on. This back and forth recursion between the nonlinear biological system and linear harvest rules generates the transition path to the steady state for any initial disturbance.
The management problem described above can be thought of as a non-cooperative (discrete-time) differential game. To make an optimal harvesting decision, each local planner must consider the actions of other local planners on the landscape grid. When making an optimal harvesting decision, local planner is aware of the optimal decision rule of all other local planners on the landscape grid and factors that into her optimal decision. Because the matrix system of equations in (4) include the first-order conditions of all local planners and all the relevant biological equations, the optimal harvesting rule in (5) defines a Markov (closed-loop) Nash equilibrium strategy (Dockner, et al., 2012). Calculating this dynamic equilibrium on a large grid with multiple species interactions would likely be intractable without the linear approximation, which allows one to write the system in matrix form and apply the Blanchard-Kahn solution technique.
9
3. Application to Forest Management and Mountain Pine Beetle Epidemics
Timber and nontimber ecosystem services in the western United States and Canada have recently experienced a spike in the population of a tree-killing insect (mountain pine beetle, MPB) native to western forests.5 The economic impacts of the current outbreak have been substantial resulting in billions of dollars in manufacturing losses and thousands unemployed (Patriquin, et al., 2007; Phillips, et al., 2007; Abbott, et al., 2009). While the cause of the
outbreak has recently been linked to historic patterns of forest management (Sims, et al., 2012) it remains unclear how the provision of ecosystem services may have contributed to the spread of the outbreak.
To investigate this phenomenon, we build an integrated bioeconomic landscape grid model with each cell representing a local or regional forest. See Figure 1 for a stylized representation of a 6 × 6 landscape forest grid. Unlike the metapopulation literature where a single species gravitates to patches with lower populations, our model allows density-dependent dispersal where predators (MPB) spread to patches where there is a greater density of prey (trees). While many of the local forests in MPB habitat are broadly managed under the direction of federal agencies like the Forest Service, specific management prescriptions such as when and where to harvest are left to the discretion of local forest managers.6 Since larger pine trees are necessary for MPB reproduction, harvesting these trees will necessarily reduce local MPB populations. However, the ability to lower local MPB populations also depends on the strategies employed by managers on neighboring forests since MPB may travel several kilometers and attack susceptible
5
The current epidemic covers over 43 million acres in British Columbia and another 9 million acres in the western U.S. (www.for.gov.bc.ca/hfp/mountain_pine_beetle/facts; Volz, 2011). The outbreak is also spreading into new habitats with unknown ecological consequences (Logan, et al., 2003) and is contributing to global warming as vast tracts of forest have been converted from a carbon sink to a carbon source (Kurz, et al., 2008).
6 A review of over 50 Forest Service timber harvesting projects throughout the range of MPB over the last seven
10
host trees (Aukema, et al., 2008). This implies that the spatiotemporal predator-prey relationship between MPB and pine trees is key to understanding the decision-making of local forest
managers, the dynamics of MPB outbreaks, and the supply of forest ecosystem services.
3.1 Biological System: Forest Dynamics
The biological component of the model involves a dynamic predator-prey relationship between MPB and the forest with time set in annual increments to match the MPB lifecycle (Samman and Logan, 2000).7 Individual forests are indexed by the pair on the landscape grid, where and . Each forest is divided into three size classes: seed base
, young trees , and adult trees (Heavilin and Powell, 2008). Young trees have a diameter at breast height (dbh) less than 8 inches. Although young trees have less defensive mechanisms and could provide enough nutrients for the larvae to develop, they seldom provide enough clearance in the inner bark for larval development. Adult trees are characterized by a dbh 8 inches and larger. While adults have the strongest defenses against MPB attack, they are also large enough to house egg galleries and act as an ample nutrient source. Each size class is measured in trees or seeds per acre. The laws of motion for the beginning-of-period density in each size class are given by:
( )
⏟
7 The bioeconomic model is similar to the one presented in two previous MPB papers: Sims et al. (2010), which
11
Each year, a proportion ( and ) of the seed base and young trees mature to the successive size class. Contributions to the seed base are made by the young and adult size classes at rates
and . Only adult trees are considered viable for commercial harvest and susceptible to natural mortality (at rate ) or MPB-induced mortality (at rate ). In addition, growth and mortality are assumed to occur prior to timber harvesting, differentiating the harvestable stock from the stock at the beginning of the next period .
Successful MPB attacks have two major consequences. First, females lay as many as 100 eggs in a single gallery. Larvae emerge from hatched eggs and spend the majority of their lifecycle inside the tree. The larvae construct feeding galleries in the phloem or inner bark of the tree. These galleries eventually girdle the tree by cutting off nutrient exchange between the roots and the tree (Samman and Logan, 2000). Second, MPB also carry blue stain fungi which
interupt water translocation, lower wood moisture content, and weaken defense mechanisms. Effects of the fungi coupled with damage to the inner bark eventually lead to tree death.
The rate of pine tree death from MPB is determined by the interaction between the number of MPB attacking the tree and the level of tree resistance (Berryman, et al., 1985). Following Heavilin and Powell (2008), we define the rate of MPB-induced mortality as
( )
( )
12
functional response in predator-prey interactions (Holling, 1959) and captures threshold dynamics characteristic of MPB (Berryman, et al., 1985).
3.2 Biological System: MPB Reproduction and Spatial Dynamics
Next, we describe the reproductive and migration processes for MPB. The relationship between MPB populations and the forest stock involves a one-year lag as adult MPBs typically emerge from the tree a year after initial infestation (Samman and Logan, 2000). Pre-migration MPB density in period t, , is a function of the density of successfully attacked trees at time
and a disturbance (with mean ) meant to capture random shocks in MPB reproduction:
. (10)
The parameter measures average MPB fecundity (the number of newly emerged beetles per successfully attacked tree). These shocks in MPB reproduction can be thought of as seasonal climatic variations in precipitation or temperature, which in turn affect how many MPB survive and emerge from a dead tree. Equation (10) describes the dynamic reproductive process of MPB within local forest .
13
( )
, (11)
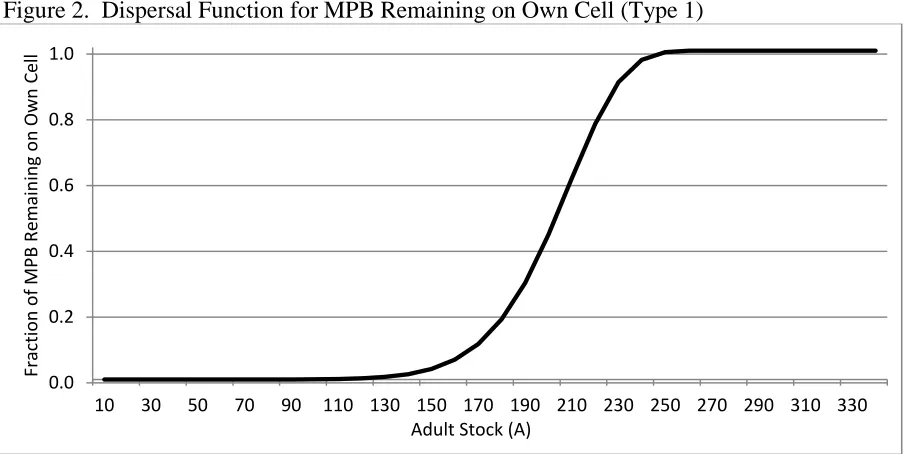
where and are fixed parameters controlling the propensity of newly emerged MPB to remain on forest . The dispersal weight, , is bounded between zero and one. If forest contains a dense supply of adult trees with a large , will approach one and all MPB remain on forest . If all the adult trees in have been killed or
harvested, , and all MPB disperse to neighboring forests. Figure 2 shows an example of the dispersal weight, , as a function of the density of adult trees.
Stage 2 represents the dispersal to neighboring forests (see figure 1 for a stylized representation of MPB dispersal to neighboring cells). Dispersal follows queen contiguity (Pfeiffer, et al., 2008) and is a function of the relative density of susceptible trees in neighboring forests. MPB dispersal to neighboring forests can represent intentional shorter-distance search for host trees or longer-distance, wind-aided travel whereby reproductive success will be greater in high-density stands of trees.8 We assume beetles do not successfully migrate to the non-forested region, implying that the landscape boundary is characterized by a reflecting barrier. The fraction of MPB migrating away from forest is given by
( )
∑ (12)
8
The specific mechanism underlying MPB dispersal is unresolved (for a review see Safranyik and Carroll (2006)). Some research has suggested that search for host trees covers relatively short distances (e.g., 250 meters or less) and longer distance dispersal (e.g., 3-5 kilometers) occurs via wind-aided travel above the tree canopy. We abstract
from the insect-level specifics and focus on the population-level outcomes by assuming MPB act as if they disperse
14
where the dispersal weights satisfy the adding-up constraint in equation (1).
The post-migration beetle population in forest is given by the sum of beetles that remains on plus those that migrate into from neighboring forests:
(
∑ ) (13)
The parameter captures the effect of resource competition on reproduction (Berryman, et al., 1985). Beetles that remain on cell and those that migrate to must compete for available space to create egg galleries given a limited stock of susceptible trees. Together, equations (11) through (13) completely characterize the spatial and temporal dynamics of the MPB population.
3.3Forest Management
Forest managers are assumed to maximize the welfare of their stakeholders: residents and users of each local forest. Utility for the representative stakeholder of each forest depends on timber products and non-timber services such as amenity values, wildlife habitat, and biodiversity. Non-timber ecosystem services depend on the quality of the forest resource, which is proxied by the stock of living adult trees, . The instantaneous utility function is therefore given by ( ), where is the relative weight stakeholders place on non-timber ecosystem services in relation to timber products.9
9
Stakeholder preferences for timber products are normalized to 1. A forest where stakeholders value non-timber
15
As discussed in Section 2, local forest managers perceive that the endogenous MPB migration effect only applies to their own and neighboring forests, while the exogenous MPB migration effect applies to the entire landscape grid. It is important to recognize that limiting the harvesting effect to neighboring forests does not imply that managers are myopic. On the
contrary, managers account for spatial MPB dynamics across the entire landscape grid and plan into the indefinite future. For example, future MPB risk resulting from an exogenous outbreak on the opposite side of the grid is considered when making the current harvesting decision.
3.3.1 Local Forest Management
Each local forest manager chooses harvests to solve the following problem:
∑
( )
where is the expectation operator conditional on information available at time . The objective function in (14) is solved subject to the forest and MPB dynamics, non-negativity constraints, and initial conditions for tree and MPB stocks. The solution for the optimal program is found through a series of substitutions that incorporate all applicable dynamics, changing the choice variable from harvest to stock of adult trees (Azariadis, 1993).
16
⏟
{ (
⏟
⏟
)
⏟
}
where
(
)
is the marginal utility of an adult tree at time t. The marginal utility is the sum of a non-timber and non-timber benefit. The left side of (15) is the immediate marginal benefit of harvesting a tree. The right side of (15) includes three future effects. The first is the marginal benefit of not harvesting: with probability a tree will be available in the following year to provide timber and non-timber benefits. The second effect is a migration cost of not harvesting: more adult trees imply more MPB will remain on the forest and more will be drawn in from neighboring forests, increasing MPB risk in . The third effect is a MPB reproduction effect associated with not harvesting: the additional MPB that remain on the forest and are drawn in
from neighbors in reproduce and increase MPB risk on the manager’s own forest in . The complete bioeconomic forest system is comprised of equations (6–13), equation (15),
non-negativity constraints and initial conditions. In theory, this system of 9 equations, I × J local forests, and time periods could be solved jointly for the beetle, tree and harvest time paths. In practice, however, there are simply too many jointly determined equations to make this
17
approximation technique presented in Section 2. For all our calibrations, the system satisfies the saddle-path stability condition and implies linear harvest rules of the form:
(16)
where is the vector of all tree classes and MPB for all cells on the landscape grid.
3.3.2 Centralized Forest Management
In the centralized approach, the forest manager maximizes the utility society receives from the landscape forest:
∑
{∑ ∑ ( )
}
The difference between the harvest choices of the local and central planners is that the latter recognizes the landscape spatial MPB externality. That is, harvest decisions in a local forest have a welfare impact on neighboring cells. While local forest managers look to neighboring cells to determine the full effect of their harvest decisions on local welfare, they ignore how their harvest decision affects welfare on neighboring cells. The central manager internalizes this spatial externality by appending the following expression to the right side of the optimal harvesting equation on cell :
∑ {
}
18
externality: harvesting an additional tree in cell causes MPB to migrate, impacting welfare on neighboring cells. The second term is the MPB reproductive externality: harvesting an additional tree in cell implies fewer MPB in neighboring cells due to less reproduction on cell . These two effects work in opposite directions; the first encourages less harvesting by the central planner while the second encourages more harvesting.
3.4Simulation Results and Discussion
In this section we present the results from a hypothetical local MPB outbreak. The outbreak involves a one-time positive perturbation to MPB fecundity in the (1,1) cell (i.e., northwest corner) of a 6 × 6 landscape forest grid.10 The MPB then spread over the grid, moving to adjacent cells according to the dispersal process. Optimal local and central forest management responds to the MPB outbreak in cell according to the harvest rule for which is a function of the 4 × 36 = 144 contemporaneous stock variables in each cell. Optimal management response is simulated over a 200 year planning horizon (T = 200) and characterized by an
increase in harvest rates and reduction in the stock of adult host trees on all local forests. The parameters and their associated definitions are shown in Table 1. A discussion of the selection process for the parameter values is presented in the appendix.
19
3.4.1 Steady-State Values: Spatial Heterogeneity and Externalities
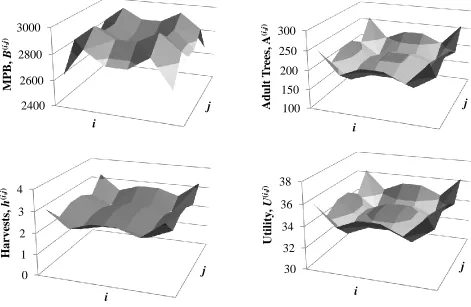
The long-run values of MPB, tree stocks, harvesting, and utility that are generated in the absence of any MPB outbreaks are shown in Table 2. Given the inherent symmetry in the landscape forest grid, we have six distinct steady states or forest types (see figure 1).11 We highlight several interesting features from Table 2. First, MPB populations are more than twice as large under a harvesting moratorium as compared to either local or central harvesting. Given our predator-prey setting, the absence of harvesting implies more host trees for MPB to
reproduce, leading to a higher MPB population. However, the higher MPB population in turn kills more trees causing the long-run equilibrium adult tree stock to be similar to either of the harvesting scenarios. MPB replace harvesting as the primary mechanism for limiting forest density. Second, steady-state MPB populations are equated across forest types when there is no harvesting. This reflects the tendency of MPB to migrate in a way that equalizes the rate of reproductive success in each cell. Yet, local managers have varying incentives to push MPB off their cell by harvesting adult trees. This gives rise to an unequal distribution of MPB across space, with intermediate forests (types 2 and 3) attracting MPB and boundary forests (types 4 and 6) repelling MPB (figure 3 presents the local harvesting case). Third, harvests and tree stocks vary significantly based on forest type. Position on the grid creates differential MPB risk even for forests that are otherwise identical. This differential MPB risk leads to a complex spatial configuration of tree stocks, MPB and harvests (figure 3). Types 4 and 6 (boundary forests) have the densest forests, the most harvesting and the lowest MPB stocks. Types 2 and 3 (intermediate forests) have the sparsest forests, the least harvesting and the highest MPB stocks. As a result, stakeholders in type 4 and 6 forests receive the highest utility while type 2 and 3 stakeholders receive the lowest utility. Fourth, stocks, harvests and utility levels are similar for
20
local and central harvesting. This occurs because the migration externality (which results in over-harvesting of adult trees as local managers encourage migration off their forest) largely offsets the reproduction externality (which results in under-harvesting as local managers ignore the costs to neighboring forests of a new MPB cohort). As a result, there is a reduced need for central management to intervene and dictate harvesting quotas on local forests. In section 3.4.4, we investigate the sensitivity of this result to variation in the fundamental parameters.
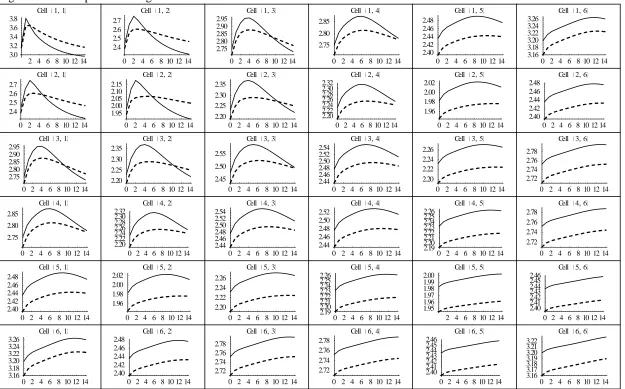
3.4.2 Transition Dynamics after a MPB Outbreak
Figures 4 and 5 contrast the impacts of optimal and no harvesting on the spatiotemporal dynamics of beetles and adult trees after a MPB outbreak. As MPB are native and endemic, there are no long-run equilibrium effects of the outbreak. There are however significantly different transition effects depending on the management regime. Figure 6 displays optimal local and central harvest paths to highlight the differences in management responses after the outbreak. All three figures show the dynamics over 15 years following the initial outbreak across the entire landscape grid.
The MPB populations are displayed in figure 4 with thick, solid and dashed lines
21
the MPB to disperse to neighboring cells. They then attack trees in the neighboring cells and start the reproduction process over again. The migration and reproduction process continues over space and time until the MPB have dispersed over the entire grid and the outbreak has dissipated through time. Figure 5 shows that adult tree densities across the grid are lower after the outbreak, but gradually regenerate as younger, immune trees mature to the adult category. MPB also spread to neighboring cells at a greater rate under optimal harvesting than under no harvesting. We discuss this effect further in the next section.
Figure 6 displays the optimal local and central harvesting strategies. Steady-state, initial harvest levels are similar for centralized planning (dashed lines) and local planning (solid lines). After the outbreak, harvest levels immediately increase across the grid. In cell (1,1), the local forest manager responds immediately to the local outbreak by harvesting additional adult trees because the expected future value of a tree declines. In the neighboring forests, cells (1,2), (2,2) & (2,1), managers also increase their harvesting in , as they anticipate the effects of the outbreak spreading to their forest in . Immediate harvesting has the benefit of providing additional timber products but also reduces the likelihood that MPB from the epidemic disperse to your local forest. The increased harvesting on neighboring cells causes additional beetles to leave these cells in . Most of these beetles will disperse to cells (1,3), (2,3), (3,3), (3,2), and (3,1) in since there are fewer susceptible host trees in cell (1,1) due to the outbreak. Anticipating this influx of MPB in , managers two cells away from the outbreak
22
of local forest managers. Even though the effect of the outbreak will not reach other forests for at least two periods, the additional harvesting on neighboring cells in creates a ripple effect that encourages all managers to immediately increase harvest levels. As the stock of adult trees decline, harvest levels gradually drop and fall below steady-state levels due to the increasing marginal non-timber value of each adult tree. With central planning, there is a similar but smoother pattern, with the initial increase in harvesting being less steep and the decline more gradual.
3.4.3 Contrasting the Degree of Spread: Optimal versus No Harvesting
A comparison of the transition paths over management regimes reveals a potential tradeoff caused by a MPB outbreak. Outbreaks under no harvesting are severe with high aggregate MPB abundances and high degrees of MPB induced tree mortality, but lower rates of dispersal across the landscape. In contrast, outbreaks with optimal harvesting see a reduction in severity, yet the MPB disperse across the landscape at a higher rate. Optimal harvesting allows humans to anticipate the MPB-induced mortality, harvest the trees before they are attacked, capture the value of the trees from the market, and moderate the severity of the outbreak. However, there is an ecological feedback response. MPB disperse quicker in their search for prey, which in turn moderates the incentive to harvest. The combined outcome sees a less severe outbreak but more spread over the landscape. These interdependencies are not present in aspatial models (or spatial models of a single species) and provide a clear contribution of this work.
23
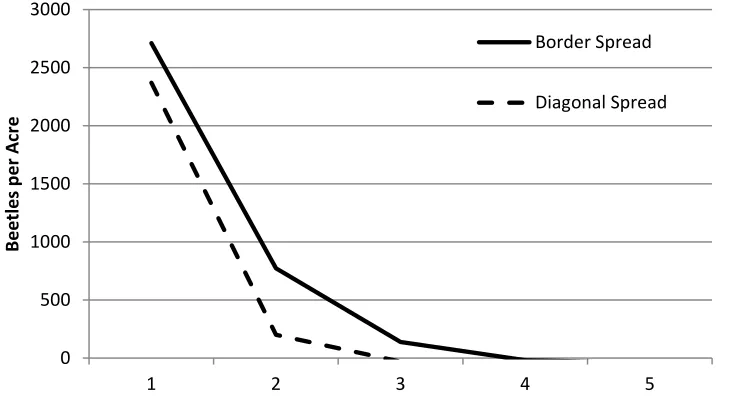
To measure the degree of MPB spread, we calculate the deviation of MPB from their endemic, steady-state level, ̃ , at a point in time after the outbreak. This measure captures pure MPB migration but also includes other factors that influence MPB population such as reproductive efficiency. Since there are more trees and fewer beetles on the borders, we contrast MPB spread in two directions – along the top border ̃ ̃ ̃ ̃ ̃ and down the
diagonal ̃ ̃ ̃ ̃ ̃ .12 Figure 7 graphs the difference in ̃ under local and no harvesting. The graph captures the influence of harvesting on MPB spread with positive values indicating that harvesting leads to more MPB spread.13 The figure highlights the tradeoff between the severity and extent of MPB epidemics: harvesting effectively limits the severity of MPB outbreaks but encourages MPB migration and increases the spread rate for MPB
epidemics. The nature of this tradeoff will differ over space given the differences in spread along the border and diagonal.
3.4.4 Sensitivity Analysis to Parameter Changes
Table 3 shows the steady-state values for a change in several fundamental parameters. Each parameter is individually decreased and increased from its value in Table 1. Drought and winter temperatures can cause changes in the number of MPB offspring from an infested tree ( ) and tree defenses against MPB attack ( ) (Heavilin and Powell, 2008). An increase in leads to more killed trees and fewer MPB. Even though more MPB emerge from an infested tree, there are fewer host trees so the MPB population falls. Likewise, a decrease in implies that a tree’s
12
Due to the symmetry of the landscape grid, horizontal and vertical spread originating from the (1,1) cell will be equal.
13 The graphs are qualitatively similar for the comparison between central management and no harvesting. Results
24
defenses have weakened and fewer MPB are needed to successfully attack a tree. This results in fewer trees and fewer MPB. In both cases, the reproduction externality increases relative to the migration externality because failure to harvest a tree results in more successful MPB
reproduction which, in turn, increases MPB migration to neighboring forests. An increase in can be interpreted as a less healthy forest with a higher natural death rate. This leads to less harvesting, fewer trees and fewer MPB. The reproduction externality now exceeds the migration externality because the migration externality is smaller due to the reduction in the MPB
population. This implies that recent changes in climate and forest health will provide an
incentive for local managers to under-harvest as a result of the spatial externality. In contrast, an increase in implies a greater preference for non-timber forest benefits and therefore less harvesting, more trees, more MPB, and a relatively greater migration externality. Surprisingly, historic efforts to place a greater weight on non-timber benefits of public forests will create an incentive for local managers to over-harvest as a result of the spatial externality. Finally,
determines how many MPB migrate off the local forest in steady state. We fix this value at 0.5 for each local forest. Although changes in have little effect on the steady-state values, we note that it has a significant effect on the spread of a MPB outbreak over the forest landscape.
4. Conclusions
25
outbreaks. The harvest rules are a function of the current tree and MPB stocks from every cell on the grid. Methodologically, the most important feature of the optimal harvest rules is that they only depend on current stocks. Therefore, rather than needing to collectively solve the entire set of first-order conditions and biological equations, the system collapses to a recursive set of simpler two-period tasks. For problems of local optimal control, this procedure allows for a much larger grid even in the presence of spatial interdependencies. The method is sufficiently general to be applicable to a wide range of spatially and temporally dynamic economic systems.
26
encourages local over-harvesting so more MPB migrate to neighboring forests, while the reproduction externality encourages under-harvesting as local planners fail to recognize that dense forests lead to more MPB and elevated MPB risk on neighboring forests. The sensitivity analysis shows that although these two externalities will always have opposite sign, they are not always of equal magnitude. This implies that the role for central management will vary across areas with differing rates of resource growth and pest susceptibility.
27 References
Abbott, B., Stennes, B., van Kooten, G. C., 2009. Mountain pine beetle, global markets, and the British Columbia forest economy. Canadian Journal of Forest Research 39, 1313-1321. Albers, H. J., 1996. Modeling Ecological Constraints on Tropical Forest Management: Spatial
Interdependence, Irreversibility, and Uncertainty. Journal of Environmental Economics and Management 30, 73-94.
Albers, H. J., Fischer, C., Sanchirico, J. N., 2010. Invasive species management in a spatially heterogeneous world: Effects of uniform policies. Resource and Energy Economics 32, 483-499.
Aukema, B. H., Carroll, A. L., Zheng, Y., Zhu, J., Raffa, K. F., Moore, R. D., Stahl, K., Taylor, S. W., 2008. Movement of outbreak populations of mountain pine beetle: influences of spatiotemporal patterns and climate. Ecography 31, 348-358.
Azariadis, C., 1993. Intertemporal Macroeconomics. Blackwell Publishers, Cambridge, MA. Bentz, B., 2006. Mountain pine beetle population sampling: inferences from Lindgren
pheromone traps and emergence cages. Canadian Journal of Forest Research 36, 351-360. Berryman, A. A., Dennis, B., Raffa, K. F., Stenseth, N. C., 1985. Evolution of optimal group
attack, with particular reference to bark beetles (Coleoptera: Scolytidae). Ecology 66, 898-903.
Bhat, M. G., Huffaker, R. G., 2007. Management of a transboundary wildlife population: A self-enforcing cooperative agreement with renegotiation and variable transfer payments. Journal of Environmental Economics and Management 53, 54-67.
Blanchard, O. J., Kahn, C. M., 1980. The Solution of Linear Difference Models under Rational Expectations. Econometrica 48, 1305-1311.
Brock, W., Xepapadeas, A., 2008. Diffusion-induced instability and pattern formation in infinite horizon recursive optimal control. Journal of Economic Dynamics and Control 32, 2745-2787.
Brock, W., Xepapadeas, A., 2010. Pattern formation, spatial externalities and regulation in coupled economic-ecological systems. Journal of Environmental Economics and Management 59, 149-164.
Brown, G., Roughgarden, J., 1997. A metapopulation model with private property and a common pool. Ecological Economics 22, 65-71.
Bulte, E. H., van Kooten, G. C., 1999. Metapopulation dynamics and stochastic bioeconomic modeling. Ecological Economics 30, 293-299.
Clarida, R., Galí, J., Gertler, M., 2000. Monetary Policy Rules and Macroeconomic Stability: Evidence and Some Theory. Quarterly Journal of Economics 115, 147-180.
Cole, W. E., 1983. Interaction between Mountain Pine Beetle and Dynamics of Lodgepole Pine Stands. United States Department of Agriculture, Forest Service Res. Note INT-170. Cooley, T. F., Prescott, E. C., 1995. Economic Growth and Business Cycles. In: Cooley, T. F.,
(Ed., Frontiers of Business Cycle Research, Princeton, NJ.
Costello, C., Polasky, S., 2008. Optimal harvesting of stochastic spatial resources. Journal of Environmental Economics and Management 56, 1-18.
Dockner, E. J., Jorgensen, S., Van Long, N., Sorger, G., 2012. Differential games in economics and management science. Cambridge University Press.
28
Farmer, R. E. A., 2002. Macroeconomics of Self-fulfilling Prophecies. The MIT Press, Cambridge, MA.
Fredriksson, P. G., Millimet, D. L., 2002. Strategic Interaction and the Determination of Environmental Policy across U.S. States. Journal of Urban Economics 51, 101-122. Fujita, M., 1976. Spatial patterns of urban growth: Optimum and market. Journal of Urban
Economics 3, 209-241.
Heavilin, J., Powell, J., 2008. A novel method of fitting spatio-temporal models to data, with applications to the dynamics of mountain pine beetles. Natural Resource Modeling 21, 489-524.
Hof, J. G., Bevers, M., Joyce, L. A., Kent, B., 1994. An Integer Programming Approach for Spatially and Temporally Optimizing Wildlife Populations. Forest Science 40, 177-191. Holling, C. S., 1959. The components of predation as revealed by a study of small predation of
the European pine sawfly. Canadian Entomologist 91, 30.
Horan, R. D., Wolf, C. A., Fenichel, E. P., Matthews Jr., K. H., 2005. Spatial Management of Wildlife Disease. Review of Agricultural Economics 27, 483-490.
Kaffine, D. T., Costello, C., 2011. Unitization of Spatilly Connected Renewable Resources. The B.E. Journal of Economics Analysis and Policy 11, Article 15.
Koch, P., 1996. Lodgepole Pine in North America. Forest Products Society.
Konoshima, M., Montgomery, C. A., Albers, H. J., Arthur, J. L., 2008. Spatial-endogenous fire risk and efficient fuel management and timber harvest. Land Economics 84, 449-468. Krugman, P., 1991. Geography and Trade. MIT Press, Cambridge, MA.
Kurz, W. A., Dymond, C. C., Stinson, G., Rampley, G. J., Neilson, E. T., Carroll, A., Ebata, T., Safranyik, L., 2008. Mountain pine beetle and forest carbon feedback to climate change. Nature 452, 987-990.
Logan, J. A., Regniere, J., Powell, J. A., 2003. Assessing the impacts of global warming on forest pest dynamics. Frontiers in Ecology and the Environment 1, 130-137.
Lotan, J. E., Critchfield, W. B., 1990. Pinus contorta Dougl. ex Loud. In: Burns, R. M., Honkala, B. H., (Eds.), Silvics of North America Vol. 1 confiers. Department of Agriculture, Washington, D.C., pp. 302-315.
Millennium Ecosystem Assessment, 2005. Ecosystems and Human Well-Being: Synthesis, Island Press, Washington, DC
Patriquin, M. N., Wellstead, A. M., White, W. A., 2007. Beetles, trees, and people: Regional economic impact sensitivity and policy considerations related to the mountain pine beetle infestation in British Columbia, Canada. Forest Policy and Economics 9, 938-946. Pfeiffer, D., Robinson, T., Stevenson, M., Stevens, K. B., Rogers, D. J., Clements, A. C. A.,
2008. Spatial analysis in epidemiology. Oxford University Press, New York.
Phillips, B. R., Breck, J. A., Nickel, T. J., 2007. Managing the economic impacts of mountain pine beetle outbreaks in Alberta, Information Bulletin. Western Centre for Economic Research, Edmonton, Alberta.
Rosie, J. P., 1990. Multicriteria Nonlinear Programming for Optimal Spatial Allocation of Stands. Forest Science 36, 487-501.
Row, C., Kaiser, H. F., Sessions, J., 1981. Discount rate for long-term Forest Service investments. Journal of Forestry 79, 367-369.
29
Synthesis of Its Biology, Management and Impacts on Lodgepole Pine, Victoria, British Columbia, pp. 3-66.
Safranyik, L., Shrimpton, D. M., Whitney, H., 1974. Management of Lodgepole Pine to Reduce Losses from the Mountain Pine Beetle. Canadian Forest Service, Pacific Forest Research Station, Forest Technical Report 1:24.
Samman, S., Logan, J., 2000. Assessment and response to bark beetle outbreaks in the Rocky Mountain area. In: Service, U. F., (Ed., RMRS-GTR-62. USDA Forest Service. Sanchirico, J., Wilen, J. E., Coleman, C., 2010. Optimal Rebuilding of a Metapopulation.
American Journal of Agricultural Economics 92, 1087-1102.
Sanchirico, J. N., Wilen, J. E., 1999. Bioeconomics of Spatial Exploitation in a Patchy Environment. Journal of Environmental Economics and Management 37, 129-150. Sanchirico, J. N., Wilen, J. E., 2005. Optimal spatial management of renewable resources: matching policy scope to ecosystem scale. Journal of Environmental Economics and Management 50, 23-46.
Sims, C., Aadland, D., Finnoff, D., 2010. A Dynamic Bioeconomic Analysis of Mountain Pine Beetle Epidemics. Journal of Economic Dynamics and Control 34, 2407-2419.
Sims, C., Aadland, D., Finnoff, D., Powell, J. A., 2012. How ecosystem service provision can increase forest mortality from insect outbreaks. Land Economics forthcoming.
Smith, M. D., Sanchirico, J. N., Wilen, J. E., 2009. The economics of spatial-dynamic processes: Applications to renewable resources. Journal of Environmental Economics and
Management 57, 104-121.
Smith, M. D., Wilen, J. E., 2003. Economic impacts of marine reserves: the importance of spatial behavior. Journal of Environmental Economics and Management 46, 183-206.
Smith, W. B., Miles, P. D., Perry, C. H., Pugh, S. A., 2009. Forest Resources of the United States, 2007. USDA Forest Service, Washington, DC, pp. 336.
Swallow, S. K., Parks, P. J., Wear, D. N., 1990. Policy-relevant nonconvexities in the production of multiple forest benefits. Journal of Environmental Economics and Management 19, 264-280.
Swallow, S. K., Talukdar, P., Wear, D. N., 1997. Spatial and Temporal Specialization in Forest Ecosystem Management under Sole Ownership. American Journal of Agricultural Economics 79, 311-326.
Swallow, S. K., Wear, D. N., 1993. Spatial Interactions in Multiple-Use Forestry and Substitution and Wealth Effects for the Single Stand. Journal of Environmental Economics and Management 25, 103-120.
Tabuchi, T., Thisse, J.-F., 2002. Taste heterogeneity, labor mobility and economic geography. Journal of Development Economics 69, 155-177.
Volz, M., 2011. Knowledge gaps abound in beetle-kill forest fires, The Register-Guard, Eugene, Oregon.
www.for.gov.bc.ca/hfp/mountain_pine_beetle/facts, Facts About B.C.'s Mountain Pine Beetle, British Columbia Ministry of Forests, Lands and Natural Resource Operations.
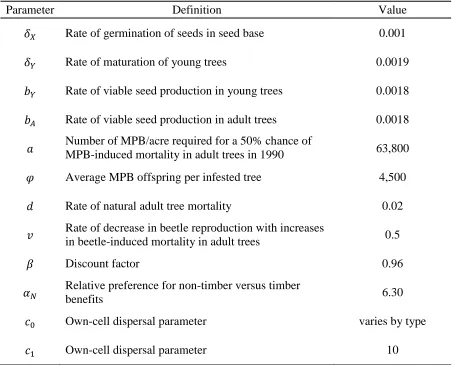
30 Table 1. Model parameters
Parameter Definition Value
Rate of germination of seeds in seed base 0.001
Rate of maturation of young trees 0.0019
Rate of viable seed production in young trees 0.0018 Rate of viable seed production in adult trees 0.0018 Number of MPB/acre required for a 50% chance of
MPB-induced mortality in adult trees in 1990 63,800
Average MPB offspring per infested tree 4,500
Rate of natural adult tree mortality 0.02
Rate of decrease in beetle reproduction with increases
in beetle-induced mortality in adult trees 0.5
Discount factor 0.96
Relative preference for non-timber versus timber
benefits 6.30
Own-cell dispersal parameter varies by type
Own-cell dispersal parameter 10
31
Table 2. Steady-State Values for No, Local, and Central Harvesting Variable
(per acre)
Type
1 2 3 4 5 6
No H
arv
esti
ng
MPB, 7657.21 7657.21 7657.21 7657.21 7657.21 7657.21
Adult Trees, 203.04 161.87 183.02 224.28 198.60 258.45
Harvests, 0 0 0 0 0 0
Loc
al Ha
rve
sti
ng
MPB, 2820.08 2981.08 2897.16 2758.05 2862.11 2651.45
Adult Trees, 201.12 163.92 182.77 222.03 198.66 255.36
Harvests, 2.46 1.97 2.22 2.74 2.42 3.19
Utility, 34.40 32.88 33.69 35.13 34.30 36.16
C
entra
l Ha
rv
esti
ng
MPB, 2975.60 3183.13 3074.04 2839.19 3032.07 2759.78
Adult Trees, 201.64 165.25 183.74 221.61 199.53 255.28
Harvests, 2.43 1.94 2.18 2.71 2.38 3.15
32 Table 3. Sensitivity Analysis to Parameter Changes
Parameter % Change
Variable (under Optimal Central Planning)
net externality (migration + reproduction)
-5%† 2.40 227.41 3883.63 -0.0022
0% 2.45 203.55 2978.79 -0.0013
+5%† 2.48 184.46 1758.47 -0.0005
-10% 2.41 171.56 772.44 +0.0001
0% 2.45 203.55 2978.79 -0.0013
+10% 2.40 246.61 4715.70 -0.0026
-10% 2.58 203.51 2508.47 -0.0009
0% 2.45 203.55 2978.79 -0.0013
+10% 2.34 203.58 3347.59 -0.0017
-10% 2.41 203.91 4245.80 -0.0025
0% 2.45 203.55 2978.79 -0.0013
+10% 2.32 201.26 1630.26 +0.0001
-10% 2.46 203.96 2964.37 -0.0009
0% 2.45 203.55 2978.79 -0.0013
+10% 2.44 203.17 2992.17 -0.0015
Notes. The variables are measured as the average steady-state values across the 36 cells on the landscape forest grid. The net spatial externality is given by equation (18). A negative number indicates that the migration externality (first term in equation (18)) dominates the reproduction externality (second term in equation (18)). A positive value indicates that the reproduction externality dominates the migration externality. (†) Steady-states
33
Figure 1. Stylized Representation of the Forest Landscape and Beetle Dispersal
Type 6 (1,1) Type 5 (1,2) Type 4 (1,3) Type 4 (1,4) Type 5 (1,5) Type 6 (1,6) Type 5 (2,1) Type 3 (2,3) Type 3 (2,4) Type 2 (2,5) Type 5 (2,6) Type 4 (3,1) Type 3 (3,2) Type 1 (3,3) Type 1 (3,4) Type 3 (3,5) Type 4 (3,6) Type 4 (4,1) Type 3 (4,2) Type 1 (4,3) Type 1 (4,4) Type 3 (4,5) Type 4 (4,6) Type 5 (5,1) Type 2 (5,2) Type 3 (5,3) Type 3 (5,4) Type 2 (5,5) Type 5 (5,6) Type 6 (6,1) Type 5 (6,2) Type 4 (6,3) Type 4 (6,4) Type 5 (6,5) Type 6 (6,6)
Notes. The green shaded area represents the forest landscape while the brown shaded area is the non-forested perimeter. There are six types of cells based on symmetric grid positions. Each cell contains an index representing row and column . The figure in cell (2,2) represents beetle dispersal to neighboring cells based on a queen contiguity pattern.
34
Figure 2. Dispersal Function for MPB Remaining on Own Cell (Type 1)
Notes. The own-dispersal weight is ( ( ) ), where and 0.0
0.2 0.4 0.6 0.8 1.0
10 30 50 70 90 110 130 150 170 190 210 230 250 270 290 310 330
Fract
ion
o
f M
PB
Re
m
ain
in
g
o
n
Ow
n
Ce
ll
35
Figure 3. Steady-State Values for the Local Forest Manager across a 6 × 6 Forest Grid
2400 2600 2800 3000
j
M
P
B
,
B
(
i
,
j
)
i
100 150 200 250 300
j
Adu
lt
T
re
es,
A
(
i
,
j
)
i
0 1 2 3 4
j
Harve
sts,
h
(
i
,
j
)
i
30 32 34 36 38
j
Ut
il
ity
,
U
(
i
,
j
)
36
Figure 4. Landscape Mountain Pine Beetle (MPB) Dispersal after Local Outbreak
Notes. The thick solid line represents MPB dynamics under no harvesting. The solid (dashed) lines represent MPB dynamics under optimal local (central) planning. The vertical axis measures number of MPB per acre on each local forest; the horizontal axis measures time in years. A MPB outbreak occurs in cell (1,1) at time . The initial outbreak is ten times the steady-state MPB population in cell (1,1). The initial outbreak is restricted to be equal under the three cases.
0 2 4 6 8 10 12 14 0 2000 4000 6000 8000 10000 12000
Cell 1, 1
0 2 4 6 8 10 12 14 0
2000 4000 6000 8000
Cell 1, 2
0 2 4 6 8 10 12 14 0
2000 4000 6000 8000
Cell 1, 3
0 2 4 6 8 10 12 14 0
2000 4000 6000 8000
Cell 1, 4
0 2 4 6 8 10 12 14 0
2000 4000 6000 8000
Cell 1, 5
0 2 4 6 8 10 12 14 0
2000 4000 6000 8000
Cell 1, 6
0 2 4 6 8 10 12 14 0
2000 4000 6000 8000
Cell 2, 1
0 2 4 6 8 10 12 14 0
2000 4000 6000 8000
Cell 2, 2
0 2 4 6 8 10 12 14 0
2000 4000 6000 8000
Cell 2, 3
0 2 4 6 8 10 12 14 0
2000 4000 6000 8000
Cell 2, 4
0 2 4 6 8 10 12 14 0
2000 4000 6000 8000
Cell 2, 5
0 2 4 6 8 10 12 14 0
2000 4000 6000 8000
Cell 2, 6
0 2 4 6 8 10 12 14 0
2000 4000 6000 8000
Cell 3, 1
0 2 4 6 8 10 12 14 0
2000 4000 6000 8000
Cell 3, 2
0 2 4 6 8 10 12 14 0
2000 4000 6000 8000
Cell 3, 3
0 2 4 6 8 10 12 14 0
2000 4000 6000 8000
Cell 3, 4
0 2 4 6 8 10 12 14 0
2000 4000 6000 8000
Cell 3, 5
0 2 4 6 8 10 12 14 0
2000 4000 6000 8000
Cell 3, 6
0 2 4 6 8 10 12 14 0
2000 4000 6000 8000
Cell 4, 1
0 2 4 6 8 10 12 14 0
2000 4000 6000 8000
Cell 4, 2
0 2 4 6 8 10 12 14 0
2000 4000 6000 8000
Cell 4, 3
0 2 4 6 8 10 12 14 0
2000 4000 6000 8000
Cell 4, 4
0 2 4 6 8 10 12 14 0
2000 4000 6000 8000
Cell 4, 5
0 2 4 6 8 10 12 14 0
2000 4000 6000
Cell 4, 6
0 2 4 6 8 10 12 14 0
2000 4000 6000 8000
Cell 5, 1
0 2 4 6 8 10 12 14 0
2000 4000 6000 8000
Cell 5, 2
0 2 4 6 8 10 12 14 0
2000 4000 6000 8000
Cell 5, 3
0 2 4 6 8 10 12 14 0
2000 4000 6000 8000
Cell 5, 4
0 2 4 6 8 10 12 14 0
2000 4000 6000
Cell 5, 5
0 2 4 6 8 10 12 14 0
2000 4000 6000
Cell 5, 6
0 2 4 6 8 10 12 14 0
2000 4000 6000 8000
Cell 6, 1
0 2 4 6 8 10 12 14 0
2000 4000 6000 8000
Cell 6, 2
0 2 4 6 8 10 12 14 0
2000 4000 6000 8000
Cell 6, 3
0 2 4 6 8 10 12 14 0
2000 4000 6000
Cell 6, 4
0 2 4 6 8 10 12 14 0
2000 4000 6000
Cell 6, 5
0 2 4 6 8 10 12 14 0
2000 4000 6000
37 Figure 5. Adult Trees Stock after Local MPB Outbreak
Notes. The thick solid line represents adult tree stocks under no harvesting. The solid (dashed) lines represent adult tree stocks under optimal local (central) planning. The vertical axis measures the number of adult trees per acre on each local forest. The horizontal axis measures time in years. A MPB outbreak occurs in cell (1,1) at time .
0 2 4 6 8 10 12 14 235
240 245 250 255
Cell 1, 1
2 4 6 8 10 12 14 188 190 192 194 196 198
Cell 1, 2
0 2 4 6 8 10 12 14 216
218 220 222 224
Cell 1, 3
0 2 4 6 8 10 12 14 218 219 220 221 222 223 224
Cell 1, 4
0 2 4 6 8 10 12 14 197.5
198.0 198.5 199.0
Cell 1, 5
0 2 4 6 8 10 12 14 254
255 256 257 258
Cell 1, 6
2 4 6 8 10 12 14 188 190 192 194 196 198
Cell 2, 1
2 4 6 8 10 12 14 156
158 160 162 164
Cell 2, 2
0 2 4 6 8 10 12 14 177 178 179 180 181 182 183
Cell 2, 3
0 2 4 6 8 10 12 14 180.0 180.5 181.0 181.5 182.0 182.5 183.0
Cell 2, 4
0 2 4 6 8 10 12 14 161.5 162.0 162.5 163.0 163.5 164.0 164.5 165.0
Cell 2, 5
0 2 4 6 8 10 12 14 198.0 198.2 198.4 198.6 198.8 199.0 199.2
Cell 2, 6
0 2 4 6 8 10 12 14 216
218 220 222 224
Cell 3, 1
0 2 4 6 8 10 12 14 177 178 179 180 181 182 183
Cell 3, 2
0 2 4 6 8 10 12 14 198 199 200 201 202 203
Cell 3, 3
2 4 6 8 10 12 14 199
200 201 202 203
Cell 3, 4
2 4 6 8 10 12 14 182.0 182.2 182.4 182.6 182.8 183.0 183.2
Cell 3, 5
0 2 4 6 8 10 12 14 221.5 222.0 222.5 223.0 223.5 224.0
Cell 3, 6
0 2 4 6 8 10 12 14 218 219 220 221 222 223 224
Cell 4, 1
0 2 4 6 8 10 12 14 180.0 180.5 181.0 181.5 182.0 182.5 183.0
Cell 4, 2
2 4 6 8 10 12 14 199
200 201 202 203
Cell 4, 3
0 2 4 6 8 10 12 14 200.0 200.5 201.0 201.5 202.0 202.5 203.0
Cell 4, 4
2 4 6 8 10 12 14 182.4
182.6 182.8 183.0 183.2
Cell 4, 5
2 4 6 8 10 12 14 221.5 222.0 222.5 223.0 223.5 224.0
Cell 4, 6
0 2 4 6 8 10 12 14 197.5
198.0 198.5 199.0
Cell 5, 1
0 2 4 6 8 10 12 14 161.5 162.0 162.5 163.0 163.5 164.0 164.5 165.0
Cell 5, 2
2 4 6 8 10 12 14 182.0 182.2 182.4 182.6 182.8 183.0 183.2
Cell 5, 3
2 4 6 8 10 12 14 182.4
182.6 182.8 183.0 183.2
Cell 5, 4
0 2 4 6 8 10 12 14 162.0 162.5 163.0 163.5 164.0 164.5 165.0
Cell 5, 5
0 2 4 6 8 10 12 14 198.6
198.8 199.0 199.2
Cell 5, 6
0 2 4 6 8 10 12 14 254
255 256 257 258
Cell 6, 1
0 2 4 6 8 10 12 14 198.0 198.2 198.4 198.6 198.8 199.0 199.2
Cell 6, 2
0 2 4 6 8 10 12 14 221.5 222.0 222.5 223.0 223.5 224.0
Cell 6, 3
2 4 6 8 10 12 14 221.5 222.0 222.5 223.0 223.5 224.0
Cell 6, 4
0 2 4 6 8 10 12 14 198.6
198.8 199.0 199.2
Cell 6, 5
0 2 4 6 8 10 12 14 255.0 255.5 256.0 256.5 257.0 257.5 258.0 258.5
38 Figure 6. Landscape Harvesting after Local MPB Outbreak
Notes. The solid (dashed) lines represent adult tree harvesting under optimal local (central) planning. The vertical axis measures the number of adult trees harvested per acre on each local forest. The horizontal axis measures time in years. A MPB outbreak occurs in cell (1,1) at time .
2 4 6 8 10 12 14 3.0
3.2 3.4 3.6 3.8
Cell 1, 1
0 2 4 6 8 10 12 14 2.4
2.5 2.6 2.7
Cell 1, 2
0 2 4 6 8 10 12 14 2.75
2.80 2.85 2.90 2.95
Cell 1, 3
0 2 4 6 8 10 12 14 2.75
2.80 2.85
Cell 1, 4
0 2 4 6 8 10 12 14 2.40
2.42 2.44 2.46 2.48
Cell 1, 5
0 2 4 6 8 10 12 14 3.16 3.18 3.20 3.22 3.24 3.26
Cell 1, 6
0 2 4 6 8 10 12 14 2.4
2.5 2.6 2.7
Cell 2, 1
0 2 4 6 8 10 12 14 1.95
2.00 2.05 2.10 2.15
Cell 2, 2
0 2 4 6 8 10 12 14 2.20
2.25 2.30 2.35
Cell 2, 3
0 2 4 6 8 10 12 14 2.20 2.22 2.24 2.26 2.28 2.30 2.32
Cell 2, 4
0 2 4 6 8 10 12 14 1.96
1.98 2.00 2.02
Cell 2, 5
0 2 4 6 8 10 12 14 2.40
2.42 2.44 2.46 2.48
Cell 2, 6
0 2 4 6 8 10 12 14 2.75
2.80 2.85 2.90 2.95
Cell 3, 1
0 2 4 6 8 10 12 14 2.20
2.25 2.30 2.35
Cell 3, 2
0 2 4 6 8 10 12 14 2.45
2.50 2.55
Cell 3, 3
0 2 4 6 8 10 12 14 2.44 2.46 2.48 2.50 2.52 2.54
Cell 3, 4
0 2 4 6 8 10 12 14 2.20
2.22 2.24 2.26
Cell 3, 5
0 2 4 6 8 10 12 14 2.72
2.74 2.76 2.78
Cell 3, 6
0 2 4 6 8 10 12 14 2.75
2.80 2.85
Cell 4, 1
0 2 4 6 8 10 12 14 2.20 2.22 2.24 2.26 2.28 2.30 2.32
Cell 4, 2
0 2 4 6 8 10 12 14 2.44 2.46 2.48 2.50 2.52 2.54
Cell 4, 3
0 2 4 6 8 10 12 14 2.44
2.46 2.48 2.50 2.52
Cell 4, 4
0 2 4 6 8 10 12 14 2.19 2.20 2.21 2.22 2.23 2.24 2.25 2.26
Cell 4, 5
0 2 4 6 8 10 12 14 2.72
2.74 2.76 2.78
Cell 4, 6
0 2 4 6 8 10 12 14 2.40
2.42 2.44 2.46 2.48
Cell 5, 1
0 2 4 6 8 10 12 14 1.96
1.98 2.00 2.02
Cell 5, 2
0 2 4 6 8 10 12 14 2.20
2.22 2.24 2.26
Cell 5, 3
0 2 4 6 8 10 12 14 2.19 2.20 2.21 2.22 2.23 2.24 2.25 2.26
Cell 5, 4
2 4 6 8 10 12 14 1.95 1.96 1.97 1.98 1.99 2.00
Cell 5, 5
0 2 4 6 8 10 12 14 2.40 2.41 2.42 2.43 2.44 2.45 2.46
Cell 5, 6
0 2 4 6 8 10 12 14 3.16 3.18 3.20 3.22 3.24 3.26
Cell 6, 1
0 2 4 6 8 10 12 14 2.40
2.42 2.44 2.46 2.48
Cell 6, 2
0 2 4 6 8 10 12 14 2.72
2.74 2.76 2.78
Cell 6, 3
0 2 4 6 8 10 12 14 2.72
2.74 2.76 2.78
Cell 6, 4
0 2 4 6 8 10 12 14 2.40 2.41 2.42 2.43 2.44 2.45 2.46
Cell 6, 5
0 2 4 6 8 10 12 14 3.16 3.17 3.18 3.19 3.20 3.21 3.22
39
Figure 7. Relative Degree of MPB Spread: ‘Local Optimal’ less ‘No’ Harvesting
Notes. The curves are a relative measure of MPB spread. The measures are calculated as the difference between MPB populations (deviations from steady states) with local optimal versus no harvesting. A positive number indicates greater spread of MPB with (local optimal) harvesting as compared to no harvesting. The horizontal axis measures the number of periods after the initial MPB outbreak or alternatively the (diagonal or horizontal) distance from the epicenter of the outbreak in cell (1,1).
0 500 1000 1500 2000 2500 3000
1 2 3 4 5
B
e
e
tles p
e
r
A
cr
e
Time Since Outbreak & Distance from Epicenter Border Spread
40 Figure 8. Aggregate Mountain Pine Beetle Population
Notes. The thick line indicates a no harvesting policy; the solid thin line is optimal local harvesting; the dashed line is optimal central harvesting. Aggregate MPB populations are the sum over the 36 cells shown in Figure 3.
20 40 60 80 100 Years
100000 150000 200000 250000 300000
41 APPENDIX
Here, we present additional details on the calculation of the local and centralized harvesting conditions, (15) and (18). We also discuss the choice of parameter values and present a
comparison of the linear and nonlinear harvest rules.
Local first-order condition
The first-order condition for the fully forward-looking planner is
{ ( ) ∑
∑ }. (A1)
where
is the effect of harvesting working through the seed base. The harvesting condition in (15) therefore imposes two restrictions on the local manager’s choice. First, the effect of harvesting on future MPB risk is limited to and effects. This restricts the MPB migration and reproduction impacts from harvesting to neighboring forests. Second, the seed effects of harvesting, , are ignored. Due to the relatively slow growth of the forest resource, these effects are negligible even when summed over the planning horizon. For example, the seed effect in period 4, the first period a seed effect is present, is the marginal net benefit of an adult tree in period 4 multiplied by less than 1 . All
subsequent seed effects are even smaller.
42
( ) ( ) [
∑
] (A2)
where
(
) (
) (A3)
(
) . (A4)
Equation (A2) reflects the ability of the manager on local forest to harvest in period and lower MPB risk in period by encouraging migration off forest and discouraging migration to forest .
The second derivative reflects the ability of the manager to harvest in period and influence MPB risk in period :
( ) ( )
[
(
)
∑ (
)
]
(A5)
where
( ) ( ) [
∑
] (A6)
43
( )
(
)
(A8)
(A9)
( ) . (A10)
The index in (A6) runs over the set of neighbors shared by cell and cell . The effect on in (A5) occurs through both MPB reproduction and migration.
Central first-order condition
The central planner’s first-order condition includes two derivatives that account for the spatial MPB externality. The first is given in (A6) and the second is
( ) ( )
(
), (A11)
Equations (A6) and (A11) are similar to equations (A2) and (A5) except rather than harvests impacting own MPB risk, harvests impact MPB risk on neighboring forests.
Selection of the parameter values