The influence of viral RNA secondary structure on interactions with innate host cell defences
Full text
Figure
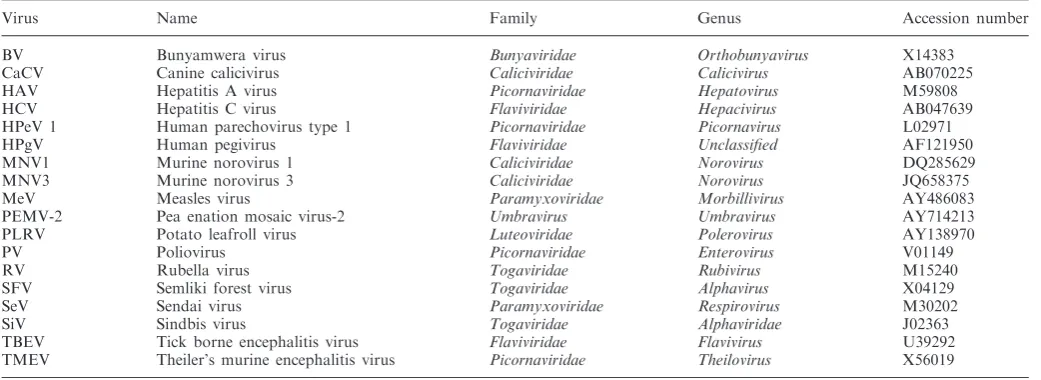

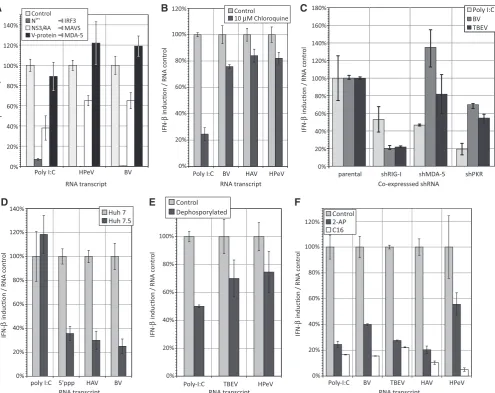

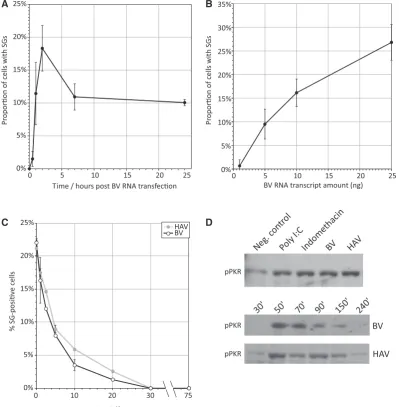



Related documents
Daubechies-10 wavelet system up to level 4 on the monthly variations in the thickness of the unfiltered water layer, and the filtering of the vertical details from
This paper proposes a simpler method and fast which is Multi Dimension of Coarse to Fine Search Method to solve the Optimal Power Flow with non-differentiable objective
Effects of intraocular lens glistenings on visual function: A prospective study and presentation of a new glistenings grading methodology.. This version of the publication may
Autotuning Control Based PID Implementation For CSTH Process- The control and simulation (while) loop used in this program is related to the practical control loop
(a) Event amplitude and energy ranges assigned to different damage mechanisms of ply fragmentation and delamination and (b) density of the AE events in the amplitude domain for
Our research aim is to offer a different perception of the medical product – and, specifically, of the prosthetic device; as a way to promote in users and external observers
The IC-WHO [33] is an adaptation of a previous question- naire used to assess quality of maternity care based on the WHO ’ s recommendations [34]. As such, in comparison to the
Two populations of' virus having subgroup-specific homogeneity (A and B) were isolated from standard avian myeloblastosis virus stocks by passage in vivo through genetically
