Copyright 0 1988 by the Genetics Society of America
An Efficient Selection Producing Structural Gene Mutants of Yeast Alcohol
Dehydrogenase Resistant to Pyrazole
Christopher
Wills
and Doreen HornDepartment of Biology, University of Cal$ornia at San Diego, La Jolla, Calqornia 92093
Manuscript received February 26, 1988 Accepted April 9, 1988
ABSTRACT
Selection for resistance to allyl alcohol in respiration-incompetent Saccharomyces cerevisiae produces a high proportion of mutants that can be localized within the ADH2 structural gene and that still,
because of the type of selection employed, retain enzyme activity. We show here that a similar type of selection produces a similarly high proportion of mutants resistant to the competitive inhibitor pyrazole. The first four mutants examined, picked at random from a collection of spontaneous pyrazole-resistant mutants, show altered-usually increased-KM values for ethanol and NAD+, and markedly increased KI values for pyrazole, compared with the wild type. When these kinetic measures and their electrophoretic mobilities were compared, all the mutants could be clearly distinguished from each other as well as from wild type. Genetic analysis shows these mutants to be close to and probably resident in the structural gene. For a variety of reasons, these mutants are even more favorable subjects for population genetic analysis and the dissection of molecular microevolution than are allyl alcohol-resistant mutants.
C
OMPARISON of the sequences of proteins and genes between closely and distantly related or- ganisms has proved to be a powerful means of inves- tigating those results of molecular evolution that have accumulated over long periods of time [see NEI (1 987) for a recent review of the data obtained from these studies and the analyses possible with them]. These studies reveal the long-term interplay of mutation pressure, chance and selection in shaping protein mol- ecules-although so far the question of the relative contributions of chance and selection to this process remains mired in controversy.A related but somewhat different set of phenomena can be investigated through an examination of molec- ular microevolution. We will define this as the set of mutations of a particular gene that form thejirst response to a speczfic selective pressure. These are clearly distin- guishable from long-term evolutionary changes, the causes of which are difficult to determine, and the mutational load at the locus, which consists of harmful mutations that are constantly arising over the course of time.
A few point mutations, existing as genetic polymor- phisms, d o not fall into either of these latter cate- gories. Among the human hemoglobin mutations,
p-
hemoglobins S, C and E are single-step alterations, sometimes with multiple points of origin (NAGEL and LABIE 1985; KULOZIK et al. 1986), that under some circumstances confer protection against the conse- quences of malaria infection [see WILLS (1981) for review]. It is certain that these represent true molec- ular microevolutionary changes, since the environ-Genetics 119: 791-795 (August, 1988)
mental pressure that increased their frequency in var- ious populations is known.
Many other mutants in the hemoglobin /3 gene may appear only in one or a few families and are often detected because of their adverse clinical conse- quences. So many are discovered each year that the same substitution often turns up in obviously unre- lated families (RAHBAR, BENNETTS and ETTINCER
1986). Mutations of this type will not be affected by specific selective pressures such as those caused by particular diseases, and will almost certainly disappear rapidly from the population. They will tend to be lost either because they are neutral or because they confer a debility on the organism that is not offset by an advantage. Some few of these latter mutants might in fact be advantageous under particular environmental circumstances, and would fall into the category of true molecular microevolutionary changes. Discovery of these environmental circumstances by the investigator is, however, very much a matter of luck. Aside from these few cases, the rest of these alterations make up the load of harmful mutations and the background of neutral mutations that are carried at every locus in a large population.
792 C. Wills and D. Hom
select for mutants in which function has been dam- aged or lost. This is analogous to the screening of the human population for pathological variants at the
p-
hemoglobin locus. But it is more difficult to design a selection in which the survival of the organism de- pends on the continued yet altered function of the gene in question.Selection for allyl alcohol resistance in petite (aero- bic respiration incompetent) yeast constitutes such a
system (WILLS 1984; HALL and WILLS 1987a, b).
Petite cells require the activity of a cytoplasmically located alcohol dehydrogenase (ADH) in order to survive (WILLS and PHELPS 1975). This is because the reducing equivalents generated by glycolysis cannot be utilized to generate ATP by the electron transport system in such cells, but must be drained off through the production of ethanol, an energy-rich intermedi- ate. If this pathway is also blocked, as it is in cells without a cytoplasmic ADH, the cell cannot survive. Yeast ADH oxidizes allyl alcohol to the poisonous aldehyde acrolein, high concentrations of which will kill the cell. Allyl alcohol resistant mutants in grande (aerobic respiration competent) cells have lost the activity of one or more of the alcohol dehydrogenases (MEGNET 1967; WILLS and PHELPS 1975; CIRIACY 1975). In petite cells subject to allyl alcohol selection, however, a large proportion of mutants are located in the structural gene for alcohol dehydrogenase I
( A D H l ) and produce altered rather than diminished enzyme activity. T h e selection is remarkably specific, and produces mutants that alter the redox balance in the cell, shifting the equilibrium concentrations of allyl alcohol and acrolein away from the poisonous
aldehyde (WILLS 1976; WILLS and MARTIN 1980).
When five of these mutants were examined at the molecular level, amino acid substitutions that could produce this kinetic shift were found scattered
through the substrate-binding domain (WILLS and
JORNVALL 1979; WILLS, KRATOFIL and MARTIN
1982). Analyses of more of these mutants at the DNA level are currently under way.
It appears from our preliminary results that allyl alcohol resistance coupled with a retention of func- tionality can be produced by many different amino acid substitutions. T o enhance the usefulness of this system for the investigation of molecular microevo- lution, it would be useful to compare the spectrum of adaptive mutants obtained using this selective pres- sure with those obtained using other, equally specific selective pressures. T h e fact that cytoplasmic alcohol dehydrogenase enzyme function is essential to the survival of the petite cell opens up the possibility of finding such selections.
We report here on a very different type of selection, that for pyrazole resistance, in which once again a large proportion of the mutants obtained are localized
in A D H l . T h e nature of this resistance allows different mutants to be distinguished clearly using electropho- retic and kinetic criteria. T h e kinetic alterations of the mutant enzymes are pronounced, and allow the mutants to be characterized kinetically with much less labor than is involved in characterizing the allyl alco- hol resistant mutants.
MATERIALS AND METHODS
Selection for pyrazole resistance was carried out using strain X2180, MATa Suc2 mall gal2 CUP1 from the Yeast Genetics Stock Center, Berkeley. Approximately lo' cells per plate were plated without mutagenesis on the following medium: 0.75% Yeast Nitrogen Base without amino acids (Difco), 2% dextrose (w/v). T o this medium was added, after autoclaving, 1 ppm antimycin A, prepared as a 1 mg/ml solution in 95% ethanol and filter sterilized. This inhibitor, by interfering with electron transport at the level of cyto- chrome b, converts cells reversibly to the petite phenotype. ADHII is not synthesized in wild-type petite (or petite phenocopy) cells, although there are mutants in which it is synthesized (CIRIACY 1979). ADHI, however, is synthesized at normal levels. The final addition to the medium was 50 mM pyrazole (Sigma), added as a sterile aqueous solution. The pyrazole gradually breaks down in solution, but plates kept at 4" can be used for two or three weeks after prepa- ration. The plates were sealed with parafilm and incubated at 30" for 1 week.
Strains picked for visualization of ADH on activity gels were grown for 4 days at 30" in 250-ml Erlenmeyer flasks containing 100 ml of complete medium ( 1 % yeast extract, 2% Bacto-Peptone, 2% dextrose, all w/v, all from Difco) in New Brunswick air-bath shakers set at 250 rpm.
Crude extracts were prepared as follows. Cells were har- vested for 5 min at 5000 rpm in a Sorvall GSA rotor, washed once in 50 mM K-PO4 buffer, pH 7.0, then centrifuged again. The washed pellet was resuspended in an equal volume of 100 mM K-P04 buffer, pH 7.0, with 10 mM b-
mercaptoethanol. The suspension was chilled on ice and an equal volume of 0.3-mm glass beads added. The cells were disrupted by four l-min periods of agitation on a vortex mixer at the highest speed, separated by one-minute periods of cooling on ice.
Aliquots of the liquid fraction were transferred to micro- fuge tubes and centrifuged for five minutes in the cold room in a Fisher 235A microcentrifuge. The supernatant was collected in new microfuge tubes and frozen at -70".
Horizontal starch gel electrophoresis of native enzymes and visualization of enzyme activity were carried out using the procedures of WILLS and PHELPS (1 975).
Genetic crosses and tetrad dissection followed the proce- dures of FINK (1 970).
Determinations of KM and K I were carried out on crude extracts by measuring the K M for ethanol at saturating NAD+ concentrations (7.75 mM), and the KM for NAD+ at saturating ethanol concentrations (100 mM). Because the mutant enzymes usually differed greatly from the wild type, a range of concentrations for the nonsaturating substrate was used initially. Five concentrations that spanned the K M
Pyrazole-Resistant Yeast Mutants 793
separated lines on Lineweaver-Burk plots. All points were measured in duplicate. Data were collected using a PDP-11 microcomputer interfaced to a Gilford 2400 spectropho- tometer.
Pyrazole is a competitive inhibitor of alcohol dehydrogen- ase. If b is the slope of the Eadie-Hofstee plot of the inhibited reaction, K M is obtained from the uninhibited reaction, and
[q
is the concentration of inhibitor, thenK , = - b [ l ] / K ~
+
[ I ] .Errors of K M estimates, for both ethanol and NAD+, are
given as the 95% confidence intervals of slopes of the lines from the Eadie-Hofstee plots. Three estimates of K , for each enzyme were obtained at different inhibitor concentrations. These data are presented as the means of these sets of estimates plus or minus twice the standard errors of the means.
RESULTS
On MVD with
50
mM pyrazole, wild-type cells grow for a few cell divisions and then stop. At the end of 1 week of incubation at 30°, 20-30 large mutant colo- nies per plate had appeared on a background of very small colonies.Twenty spontaneous pyrazole-resistant mutants
were picked from these plates and the ADHI of these mutants examined by activity-gel electrophoresis. Four mutants grew slowly and showed low levels of ADHI. These were discarded from further analysis. Of the remainder, eight migrated to the same approx- imate position as the wild type ADHI, three migrated more slowly and five more rapidly. This represented a higher proportion of mobility mutants than is nor- mally obtained with allyl alcohol selection (HALL and
WILLS
1987a).
Four of these mutants were picked at random for further genetic and kinetic analysis, the only criterion being that they produce substantial amounts of ADHI.
SP
1 1 andSP 15
show the same electrophoretic mobil-ity as the wild
type
within the limits of resolution of the method, whileSP16
migrates more rapidly andSP 17
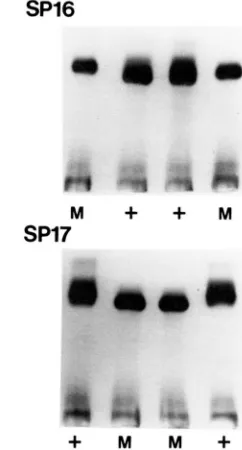
more slowly.All four of the mutants were crossed to wild type. Of the seven tetrads dissected from each cross, all segregated 2:2 for resistance to pyrazole. T h e electro- phoretic patterns of three tetrads from the
SP16
andSP17
crosses were examined. These were found to correlate in each case with the pyrazole resistance.T h e electrophoretic patterns exhibited by two of these tetrads, one from
SP16
and one fromSPl7,
are shown in Figure 1. Cells were grown in the presence of antimycin A, to convert them to petite phenocopies in which only ADHI and the mitochondrial ADH are expressed. T h e mobilities match the growth pheno- type on pyrazole, with the resistant mutant in theSP
16
cross migrating more rapidly than the wild type and the resistant mutant in theSP17
cross migrating more slowly.Crosses of
SPl
1 with all three of the other mutantsSP16
-
0 . 0
r 9
‘1 ‘ 1-
M + + M
SP17
7
+ M M +
FIGURE 1 .-Tetrads of SP l6 and SPI 7, showing 2:2 segregation for electrophoretic mobility. The symbols
+
and M refer to the inability and the ability to grow on pyrazole medium, respectively, which in the six tetrads analyzed electrophoretically co-segregated with the mobility alteration.were carried out to determine the allelism of the resistance mutants. Thirteen tetrads from the cross with
SP15, 14
from the cross withSP16
and15
from the cross withSP17
were plated on pyrazole and antimycin A medium. All segregants of all the tetrads grew. No wild-type segregants were observed, makingit highly likely that all four of the mutants are alleles. None of these crosses rules out the possibility that the four pyrazole resistant mutants are all due to modifications of the same gene, separate from the
ADH2 structural gene, that posttranscriptionally mod- ifies the ADHII product in various ways in the various mutants.
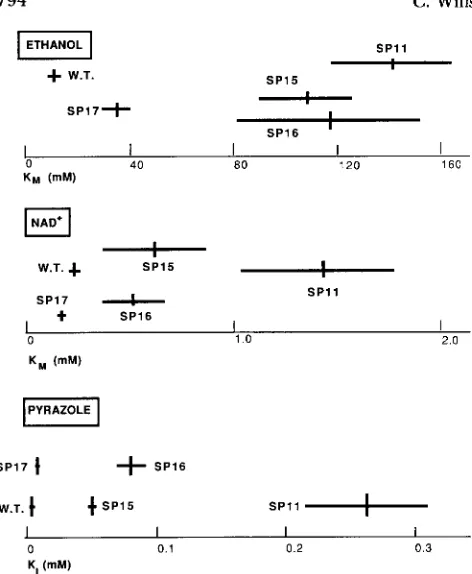
Figure
2
shows the results of the kinetic measure- ments for the wild type and the four mutant enzymes. For the ethanol and NAD+K M
values the vertical lines show the best estimates from the Eadie-Hofstee plots (in which the slope of the regression line yields the negative of the K M value). T h e horizontal bars showthe
95%
confidence intervals for the slope. For the pyrazole inhibition study, the vertical bars are the average of determinations at three levels of pyrazole, and the horizontal bars are plus or minus twice the standard error of the mean.In almost all cases,
K M
and K I values are larger and in some cases substantially larger in the mutants than in the wild type. T h e most altered mutant isSP11,
with
K M
values for ethanol and NAD+ roughly ten times those of the wild type, and a K I70
times greater. T h e wild type can be distinguished from the mutant enzymes by all three criteria. T h e only criterion by794 C. Wills and D. Horn
SP11
I
+
W.T. SP15S P I 7 + __c_ I
SP16
'
I I I I I
40 80 120 160
KM (mM)
p
J
+
W.T.
+
SP15I
SP17 1 S P I 1
t SP16
I I
0 2 0 1 0
K, (mM)
J I I I
0 0.1 0.2 0.3
K, (mM)
FIGURE 2.-KM values and K , values for the wild type and the four mutant ADHs, with the associated errors of their estimates.
The errors appear to grow larger with increasing K value, but this is an artifact of the linear scale employed; the coefficients of
variation are all quite similar. See text for details. W.T. = wild type;
SP = pyrazole-resistant mutant.
other and the wild type, however, is inhibition by pyrazole. SP17, the mutant with the smallest differ- ence from wild type in its KI, is also unusual in that it has a significantly smaller KM than wild type for NAD+. In spite of the differences in their K , values, all the mutants grow readily at 50 mM pyrazole, in contrast to the wild type which stops after a few cell divisions.
DISCUSSION
Pyrazole and its less toxic 4-substituted derivatives have been the subject of much investigation in animals because of their use in inhibiting human alcohol de- hydrogenase activity and reducing the effects of al- cohol toxicity-a toxicity which is primarily mediated not by ethanol but by acetaldehyde (DAHLBOM et al.
1974; DEIS and LESTER 1979). Pyrazole binds the enzyme less tightly than its derivatives. T h e com- pound is a heterocyclic five-membered ring, with two adjacent nitrogens:
v
N HT h e two nitrogens are involved in the formation of
a ternary complex among pyrazole, the catalytic site of the enzyme, and the cofactor NAD+. One nitrogen binds to the catalytic zinc of the enzyme itself, the other to the C-4 of the nicotinamide moiety of NAD+
(THEORELL and YONETANI 1963; BRAND~N et al.
1975). This has the effect of wedging the pyrazole molecule into the cleft leading to the active site, retarding the entry of ethanol.
Although no attempt was made at the outset to isolate completely independent pyrazole-resistant mu- tants, all four mutants are different. It therefore seems likely that many different single amino acid substitu- tions can increase the KI and protect the cell against pyrazole inhibition. Two probable classes of substitu- tion are those restricting the size of the cleft and those altering the active site.
Mutants in which the cleft size is reduced should have a clear-cut phenotype. Ethanol is a substantially smaller molecule than pyrazole, and might be able to reach the active site even if its access were somewhat restricted. This would have the effect of increasing the K M for ethanol while leaving that for NAD+ rela- tively unchanged. Possible candidate mutants for such a change are SP15 and SPl6, in which the ethanol KM
values have changed about tenfold while the NAD+
K M values have only doubled.
Because the mutant enzymes retain substantial ac- tivity, the configuration of the active site itself is unlikely to be changed greatly in any of the mutants. It may be, however, that the interatomic distance between the zinc and the nicotinamide C-4 can be altered enough to reduce the probability that the pyrazole bridge will form without completely destroy- ing the enzyme's catalytic activity. Mutant S P l l is a particularly likely candidate for such an alteration, since the KM values for both substrate and cofactor are substantially increased over the wild type.
T h e mutant that does not conform to either of these expectations is SP17, which shows the smallest shift in KM values and K,. It has a significantly lower
KM for NAD+ than the wild type. This may, under physiological conditions, so facilitate the conversion of acetaldehyde to ethanol that the cell is able to survive the presence of high levels of pyrazole.
Selection for pyrazole resistance in petite pheno- copies results in functional mutants localizable to the structural gene, just as does allyl alcohol resistance under similar circumstances. But the interpretation of the kinetic shifts seen with pyrazole resistance is much easier than it is with those seen in allyl alcohol-resistant mutants (WILLS 1984). These latter mutants owe their resistance to a much subtler kinetic mechanism, which is both harder to measure and more difficult to inter- pret.
Pyrazole-Resistant Yeast Mutants 795
which results in a shift in the equilibrium concentra- tions of allyl alcohol and its poisonous aldehyde in the direction of the harmless alcohol. This can be brought about by a great variety of alterations in the K M values for substrate or cofactor in both the forward and back reactions, and in the ratios of Vf to V b . In many of the cases that have been investigated, it has not been possible to relate these alterations to the observed shifts in the redox balance.
T h e kinetic alterations leading to pyrazole resist- ance, on the other hand, are for the most part straight- forward. T h e kinetic picture is simplified further by the fact that pyrazole will not form a ternary complex with the enzyme and NADH, only with NAD+.
Pyrazole selection provides a valuable new dimen- sion to the utility of this system for investigating molecular microevolution. It will be most instructive to compare the spectrum and distribution of muta- tions obtained with this selection to that obtained with allyl alcohol. We hope using these systems to answer the vexing question of how much of a protein mole- cule is “seen” by selective pressures. It is often stated (most recently by HARTL 1987) that parts of a mole- cule not directly involved in its function are observed to evolve more rapidly because they are more likely to accumulate neutral mutations. It can with equal facility be argued that parts of a molecule distant from critical regions can more easily accumulate molecular microevolutionary changes because these changes can alter and perhaps improve the function of the mole- cule without destroying it. We would in the latter case predict that molecular microevolutionary changes that are known to be the result of selection should be scattered widely throughout the molecule, a predic- tion that has so far been borne out for the case of allyl alcohol selection.
Experiments are under way to test this prediction further by sequencing a variety o f A D H 1 mutant genes obtained by both allyl alcohol and pyrazole selection. To facilitate this, the wild-type gene has been inserted into a shuttle plasmid derived from pEMBL18, and selected in a strain with a disrupted genomic A D H l
gene.
This work was supported by grants from the National Institutes of Health and the Universitywide Energy Research Group (Univer- sity of California).
LITERATURE CITED
BRAND~N, C.-I., H. JORNVALL, H. EKLUND and B. FURUGREN, 1975 Alcohol dehydrogenases. pp. 104-190. In: The Enzymes,
Vol. XI, Edited by P. D. BOYER. Academic Press, New York. CIRIACY, M., 1975 Genetics of alcohol dehydrogenase in Saccha-
romyces cerevisiae. I. Isolation and analysis of adh mutants. Mutat. Res. 29: 315-326.
CIRIACY, M., 1979 Isolation and characterization of cis and trans
acting elements involved in the synthesis of glucose-repressible alcohol dehydrogenase (ADHII) in Saccharomyces cereuisiae.
Mol. Gen. Genet. 176: 421-4.31.
DAHLBOM, R., B. R. TOLF, A. AKESON, G. LUNDQUIST and H. THEORELL, 1974 On the inhibitory power of some further pyrazole derivatives of horse liver alcohol dehydrogenase. Biochem. Biophys. Res. Commun. 57: 549-553.
DEIS, F. H., and D. LESTER, 1979 Biochemical pharmacology of pyrazoles. pp. 303-323. In: Biochemistry and Pharmacology of Ethanol, Vol. 2, Edited by E. MAJCHROWICZ and E. NOBLE. Plenum Press, London.
FINK, G. R., 1970 The biochemical genetics of yeast. Methods Enzymol. 17A: 59-78.
HALL, J. G., and C. WILLS, 1987a Functional alcohol dehydrogen- ase mutants of Saccharomyces cereuisiae conferring temperature- conditional allyl alcohol resistance. Genetics 115: 65-7 1 . HALL, J. G., and C. WILLS, 1987b Conditional overdominance at
an alcohol dehydrogenase locus in yeast. Genetics 117: 421- 427.
HARTL, D. L., 1987 The molecular approach to evolution (book review). Science 237: 782.
KULOZIK, A. E., J. S. WAINSCOAT, G. R. SERJEANT, B. C. KAR, B. AL-AWAMI, G. J. ESSAN, A. G . FALUSI, S. K. HAQUE, A. M. HILALI and S. KATE, 1986 Geographical survey of beta S- globin gene haplotypes: evidence for an independent Asian origin of the sickle-cell mutation. Am. J. Hum. Genet. 3 9 239- 244.
MEGNET, R., 1967 Mutants partially deficient in alcohol dehydro- genase in Schirosacchomyces pombe. Arch. Biochem. Biophys. NAGEL, R. L., and D. LABIE, 1985 The consequences and impli- cations of the multicentric origin of the Hb S gene. Prog. Clin. Biol. Res. 191: 93-103.
NEI, M., 1987 Molecular Evolutionary Genetics. Columbia Univer- sity Press, New York.
RAHBAR, S., G. A. BENNETTS and L. J. ETTINGER, 1986 Hemoglobin H Hammersmith as the cause of severe hemolytic anemia in a Chinese girl. Am. J. Pediatr. Hematol. Oncol. 8:
13-1 7.
THEORELL, H., and T. YONETANI, 1963 Crystallization of binary and ternary complexes of horse liver alcohol dehydrogenase with DPN and with DPNH and isobutyramide. Arch. Biochem. Biophys. 100: 554-558.
WILLS, C., 1976 Production of yeast alcohol dehydrogenase iso- zymes by selection. Nature 261: 26-29.
WILLS, C., 1981 Genetic Variability. Oxford University Press, Ox- ford.
WILLS, C., 1984 Structural evolution of yeast alcohol dehydrogen- ase in the laboratory. pp. 233-254. In: Microorganisms as Model Systems f o r Studying Evolution, Edited by R. P. MORTLOCK. Plenum, New York.
WILLS, C., and H. JORNVALL, 1979 Amino acid substitutions in two functional mutants of yeast alcohol dehydrogenase. Nature
WILLS, C., P. KRATOFIL and T . MARTIN, 198 1 Functional mutants of yeast alcohol dehydrogenase. pp. 305-329. In: Genetic En- gineering of Microorganismsfor Chemicals, Edited by A. HOL- LAENDER. Plenum, New York.
WILLS, C., and T . MARTIN, 1980 Alteration in the redox balance of yeast leads to allyl alcohol resistance. FEBS Lett. 119 105- 108.
WILLS, C., and J. PHELPS, 1975 A technique for the isolation of yeast alcohol dehydrogenase isozymes with altered substrate specificity. Arch. Biochem. Biophys. 236: 782-791.
121: 194-210.
279: 734-736.