Copyright © 2004, American Society for Microbiology. All Rights Reserved.
Preventive Effects of
Escherichia coli
Strain Nissle 1917 on Acute and
Chronic Intestinal Inflammation in Two Different Murine Models
of Colitis
Michael Schultz,
1* Ulrike G. Strauch,
1Hans-Jo¨rg Linde,
2Sonja Watzl,
1Florian Obermeier,
1Claudia Go¨ttl,
1Nadja Dunger,
1Nicole Grunwald,
1Ju¨rgen Scho¨lmerich,
1and Heiko C. Rath
1Departments of Internal Medicine I1and Medical Microbiology and Hygiene,2University of Regensburg,
Regensburg 93042, Germany
Received 16 July 2003/Accepted 11 December 2003
Escherichia colistrain Nissle 1917 (EcN) is as effective in maintaining remission in ulcerative colitis as is
treatment with mesalazine. This study aims to evaluate murine models of acute and chronic intestinal inflammation to study the antiinflammatory effect of EcN in vivo. Acute colitis was induced in mice with 2% dextran-sodium sulfate (DSS) in drinking water. EcN was administered from dayⴚ2 to dayⴙ7. Chronic colitis was induced by transfer of CD4ⴙCD62LⴙT lymphocytes from BALB/c mice in SCID mice. EcN was
admin-istered three times/week from week 1 to week 8 after cell transfer. Mesenteric lymph node (MLN) cytokine secretion (of gamma interferon [IFN-␥], interleukin 5 [IL-5], IL-6, and IL-10) was measured by enzyme-linked immunosorbent assay. Histologic sections of the colon were analyzed by using a score system ranging from 0 to 4. Intestinal contents and homogenized MLN were cultured, and the number ofE. coli-like colonies was determined. EcN was identified by repetitive extragenic palindromic (REP) PCR. EcN administration to DSS-treated mice reduced the secretion of proinflammatory cytokines (IFN-␥, 32,477ⴞ6,377 versus 9,734ⴞ
1,717 [Pⴝ0.004]; IL-6, 231ⴞ35 versus 121ⴞ17 [Pⴝ0.02]) but had no effect on the mucosal inflammation. In the chronic experimental colitis of the transfer model, EcN ameliorated the intestinal inflammation (histology score, 2.7ⴞ0.2 versus 1.9ⴞ0.3 [Pⴝ0.02]) and reduced the secretion of proinflammatory cytokines. Translocation of EcN and residentE. coliinto MLN was observed in the chronic colitis model but not in healthy controls. Administration of EcN ameliorated acute and chronic experimental colitis by modifying proinflam-matory cytokine secretion but had no influence on the acute DSS-induced colitis. In this model, preexisting colitis was necessary for translocation of EcN and residentE. coliinto MLN.
Probiotic microorganisms are defined as viable nutritional agents conferring benefits to the health of the human host (21). In human disease, the beneficial effects of various probiotics were demonstrated for the treatment of a variety of disorders such as infectious rotavirus-induced diarrhea in infants (15), amelioration of the side effects of antibiotic therapy (11), and prevention of atopic disease (6). Clinical trials have also sug-gested a potential role of different probiotic preparations in the treatment of inflammatory bowel disease (IBD) (36). The most striking data exist forEscherichia colistrain Nissle 1917 (EcN) in the maintenance of remission of ulcerative colitis (17, 18, 33). Treatment with EcN demonstrated beneficial effects equal to those of mesalamine (17). However, the mechanisms by which EcN mediates its effects are not fully understood. Alteration of the intestinal microflora (40), and other effects, including local and systemic immunomodulation (20, 25, 35) and barrier enhancement (22, 23), have been described previ-ously. In vitro studies that used epithelial cell lines have dem-onstrated that EcN effectively inhibits the invasion of entero-invasiveE. coliandSalmonella entericaserovar Typhimurium (8, 28).
Several animal models for acute or chronic intestinal inflam-mation have been established, and most of them are essentially associated with the commensal intestinal flora, as demon-strated by the lack of intestinal inflammation in germfree an-imals, while other rodents of the same genetic background, raised in an conventional setting, developed severe colitis (34). The majority of probiotic preparations have been already tested in gut flora-associated animal models of colitis (12, 23, 24, 37). However, no murine model of colitis has been em-ployed so far to study the antiinflammatory mechanism of EcN in detail.
This study aims to evaluate the antiinflammatory effect of EcN in murine models of acute and chronic intestinal inflam-mation.
MATERIALS AND METHODS
Animals.BALB/c and immunodeficient C.B.-17 SCID mice were purchased from Charles River Laboratories (Sulzfeld, Germany). All animals were held in standard caging conditions and received standard rodent chow and drinking water ad libitum. All animal experiments were approved by the local institutional review board.
Acute colitis. (i) Experimental design.BALB/c mice (body weight [BW], approximately 20 g) were divided into three groups. The treatment group (n⫽ 4) with dextran-sodium sulfate (DSS)-induced acute colitis received EcN; one control group (n⫽4) with DSS-induced acute colitis received plain water; and the other control group (n⫽5) with DSS-induced acute colitis received antibi-otic therapy with vancomycin-imipenem (50 mg/kg of BW), a treatment regimen known to effectively prevent acute DSS-induced colitis as shown previously (31). * Corresponding author. Mailing address: Department of Internal
Medicine I, University of Regensburg, Regensburg 93042, Germany. Phone: 49-941-944-7001. Fax: 49-941-944-7016. E-mail: michael [email protected].
372
on August 17, 2020 by guest
http://cvi.asm.org/
EcN (Ardeypharm, Herdecke, Germany) per ml resuspended in 200l of sterile saline by gastric gavage from day⫺2 (before the onset of colitis) until day 7, when the experiment was terminated. Antibiotic therapy with vancomycin-imi-penem (50 mg/kg of BW) was administered in drinking water from day⫺2 until day⫹7.
Chronic colitis. (i) Experimental design.C.B.-17 SCID mice (BW, approxi-mately 20 g) after adoptive T-cell transfer were divided into two groups. The treatment group (n⫽9) received EcN, and the control group (n⫽9) received plain water. Healthy BALB/c mice (n⫽3) treated with EcN served as negative controls. At the end of the experimental period, mice were sacrificed, and intestinal tissues, MLN, and cecal contents were removed aseptically for further evaluation.
(ii) Induction of chronic colitis.Splenic CD4⫹CD62L⫹T lymphocytes from BALB/c mice were isolated as described previously (26, 27) with slight modifi-cations. In brief, CD4⫹T cells were purified from spleen mononuclear cells of healthy mice by negative depletion using anti-CD8, anti-major histocompatibility complex II, anti-B220, and anti-CD11b antibodies (BD Biosciences Pharmingen, San Diego, Calif.) followed by anti-rat immunoglobulin G immunomagnetic microbeads (Miltenyi Biotech GmbH, Bergisch Gladbach, Germany). The re-sulting CD4⫹T-cell population was further separated by immunomagnetic mi-crobeads into CD62L⫹and CD62L⫺T cells. CD62L⫹cells (purity,⬎95%) showed high expression of CD45RB by fluorescence-activated cell sorter analy-sis. A total of 0.25⫻106CD4⫹CD62L⫹T cells were resuspended in 200l of sterile phosphate-buffered saline (PBS) and injected intraperitoneally (i.p.) into recipient C.B.-17 SCID mice. As a control, we used i.p. injection of 200l of PBS without cells.
(iii) Administration of EcN.EcN (5⫻1010CFU/ml in 200l of sterile saline [Ardeypharm]) was administered by gastric gavage two times/week from week 1 posttransfer to the end of the experiment at week 8.
Clinical assessment of colitis.At the end of the experiment, animals were clinically assessed for six clinical signs of intestinal inflammation, including gen-eral mobility and agility, stool texture, fur color and texture, anal prolapse, and rectal bleeding. One point was given for the presence of each sign. Points were added up, and a clinical score ranging from 0 to 6 was determined (Table 1).
Histological grading of colitis.Colonic tissues were removed and embedded into paraffin for histological analysis. The tissues were cut into 3- to 4-m sections, stained with hematoxylin and eosin, and scored by two investigators (F.O. and M.S.) who were unaware of the source of tissue. For histological evidence of colitis, we applied a scoring system previously validated in HLA-B27 transgenic rats and described in detail elsewhere (31, 38) to quantitate intestinal inflammation. Briefly, the tissue samples were assessed for edema, the influx of inflammatory cells, damage to the mucosal architecture, crypt abscesses, and ulcerations; the assessments were evaluated by using a scale ranging from 0 to 4 (Table 2).
Measurement of cytokine secretion.MLN (pooled from each group of mice) were collected under sterile conditions in cold cell culture medium (RPMI-1640 [10% fetal calf serum, 100 U of penicillin per ml, and 100g of streptomycin per ml from GIBCO-BRL, Eggenstein, Germany, and 3⫻10⫺5M -mercaptoetha-nol from Sigma, Deisenhofen, Germany]). Lymph nodes were mechanically disrupted and filtered through a cell strainer (70-m pore size). A total of 2⫻ 105cells/well were incubated in anti-CD3-coated plates in 200 ml of culture medium for 24 h. Subsequently, cytokine secretion (of gamma interferon [IFN-␥], interleukin 5 [IL-5], IL-6, and IL-10) was measured in the supernatant by enzyme-linked immunosorbent assay (Endogen; Pierce Biotechnology, Inc., Rockford, Ill.). Following the manufacturer’s protocol, four wells per condition
for 24 h in room air supplemented with 10% CO2, and CFU were counted. Typing ofE. coli-like isolates by REP-PCR.Colonies with typical appearance ofEnterobacteriaceaefrom cultured MLN were picked at random, and repetitive extragenic palindromic (REP) PCR was performed according to previous reports (43). Briefly, after adjustment of the concentration of genomic DNA, PCR was performed using primers REP-1 5⬘-GCGCCGICATCAGGC-3⬘and REP-2 5⬘ -ACGTCTTATCAGGCCTAC-3⬘with 30 cycles of 94°C for 1 min., 40°C for 1 min, and 65°C for 8 min. Patterns with three or more different bands were considered to represent differentE. colistrains.
Statistical analysis.For statistical analysis, Sigma Stat version 2.03 (SPSS Inc., Chicago, Ill.) software was used. For analysis of numeric values, the one-way analysis of variance andttest were used. For evaluation of ranks, the Mann-Whitney rank-sum test was used. APvalue of⬍0.05 was considered significant.
RESULTS
Acute colitis. (i) Clinical signs.Acute colitis was induced in mice via administration of DSS in the drinking water. At the end of the experiment, no difference was seen between the groups in terms of body weight and length of the colon (data not shown).
(ii) Histology.Histologically, DSS-induced colitis revealed a patchy pattern of severe mucosal damage that included a loss of crypts, necrosis, and focal influx of inflammatory cells next to unaffected areas of mucosal architecture. No beneficial ef-fect was observed after gastric administration of EcN (histol-ogy score, 3.0⫾0.8 versus 3.1⫾ 0.4 [not statistically signifi-cant]). Animals treated with vancomycin-imipenem, however, developed no histological signs of colitis, as previously docu-mented (3.1⫾0.4 versus 0.2⫾0.1 [Pⱕ0.001]) (31).
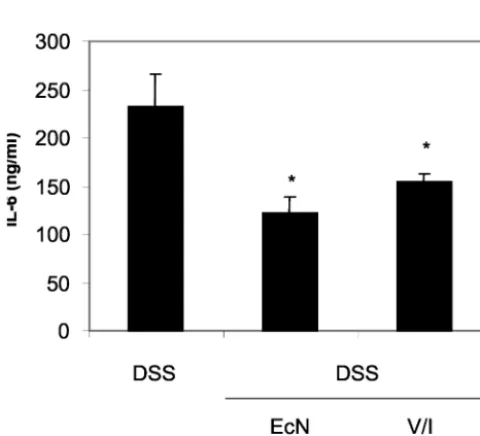
(iii) Cytokine secretion.Despite the lack of an effect of EcN administration on the histological appearance of the mice with acute colitis, levels of the proinflammatory cytokines IFN-␥ (Fig. 1) and IL-6 (Fig. 2) were significantly reduced, while the
TABLE 2. Histological grading of intestinal inflammation
Score Characteristic(s)
0...No inflammation
1...Slightly infiltrating cells in lamina propria 2...Infiltration with mononuclear cells
leading to separation of crypts, mild mucosla hyperplasia
3...Massive infiltration with inflammatory cells leading to disturbed mucosal architecture, loss of goblet cells, marked mucosal hyperplasia 4...Crypt abscesses, ulceration
on August 17, 2020 by guest
secretion of IL-5 and IL-10 by cells from MLN was not changed by probiotic or antibiotic treatment (data not shown).
Chronic colitis. (i) Clinical signs.Chronic murine colitis was induced by transfer of naïve CD4⫹CD62L⫹splenic T lympho-cytes into immunodeficient SCID mice, and recipient animals were treated regularly with EcN immediately after cell trans-fer. At the end of the 8-week treatment period, mice receiving EcN appeared healthier than did untreated mice. This result was reflected by a significantly lower clinical score (Fig. 3).
However, changes in body weight were unaffected by the pro-biotic treatment, as mice given EcN lost weight simultaneously with untreated animals (data not shown).
(ii) Histology.SCID mice injected i.p. with CD4⫹CD62L⫹ T-lymphocytes developed severe intestinal inflammation (Fig. 4). Histologically, the colitis was characterized by a thickening of the mucosa due to edema and an influx of inflammatory cells (Fig. 4A). As shown in Fig. 4B, the intestinal inflammation was ameliorated by oral administration of EcN, a finding reflected by a reduction of the histological score (2.7⫾0.2 versus 1.9⫾ 0.3 [P⫽0.02]).
(iii) Cytokine secretion.Lymphocytes from MLN from un-treated CD4⫹ CD62L⫹ T-cell-injected SCID mice secreted high levels of proinflammatory cytokines (Fig. 5). Administra-tion of EcN led to a significant reducAdministra-tion of IFN-␥(Fig. 5A) (P
ⱕ 0.03), IL-5 (Fig. 5B) (P⫽ 0.02), and IL-6 (Fig. 5C) (P⫽ 0.02), while the level of secretion of the antiinflammatory cy-tokine IL-10 remained unchanged (data not shown).
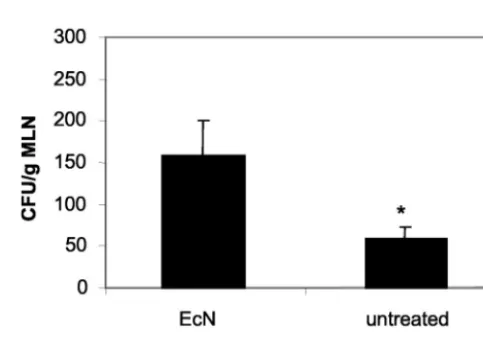
(iv) Analysis of the impact of EcN administration on the residentE. colipopulation in chronic colitis.REP-PCR dem-onstrated successful colonization with EcN in mice treated with EcN (data not shown). Incubation of cecal contents from animals who were fed EcN revealed significantly higher con-centrations of enterobacterial colonies than those of untreated controls (Fig. 6) (Pⱕ0.001), suggesting that no replacement of the resident E. coli flora with EcN occurred. Furthermore, EcN, together with otherEnterobacteriaceae, was grown from the MLN of colitic mice treated with EcN, indicating translo-cation of viable microorganisms (Fig. 7). No bacteria were cultivated from the MLN of healthy BALB/c mice after 8 weeks of EcN administration.
DISCUSSION
The exact etiology of chronic IBD is still unknown but seems complex and multifactorial. From human studies of and animal models for experimental colitis, increasing evidence is gener-ated that the resident intestinal flora plays a critical role in the development of the intestinal inflammation on a given genetic
FIG. 1. Reduction of IFN-␥secretion by MLN in DSS-induced colitis following the administration of EcN or vancomycin-imipenem (V/I). Values given are means⫾SEM (picograms per milliliter). *,P
⫽0.004 versus DSS.
FIG. 2. Reduction of IL-6 secretion by MLN in DSS-induced colitis following the administration of EcN or vancomycin-imipenem (V/I). Values given are means⫾SEM (picograms per milliliter). *,P⫽0.02 versus DSS.
FIG. 3. Clinical score of CD4⫹CD62L⫹SCID mice untreated or
treated with EcN in comparison to the results seen with the control group injected i.p. with PBS. *,P⬍0.001 versus CD62L.
on August 17, 2020 by guest
http://cvi.asm.org/
background. IL-2- and IL-10-knockout mice as well as HLA-B27 transgenic rats developed colitis when raised in specific-pathogen-free conditions, but they remained almost disease free under germfree conditions (32, 38, 39). Furthermore, clin-ical observations demonstrate beneficial effects of probiotic microorganisms in the treatment of IBD. EcN especially was shown to be as effective as mesalazine in the maintenance of remission in patients with ulcerative colitis (17). So far, how-ever, no animal model of experimental colitis has been tested for further investigation of the mechanisms by which EcN may exert its beneficial effects.
Using a preventive setup in the present study, we demon-strated that EcN has immunomodulatory properties in acute
and chronic experimental colitis, possibly mediated by translo-cation into MLN and alteration of the proinflammatory cyto-kine secretion profile—but EcN has no short-term effect on acute toxic mucosal damage.
We have chosen two different models of colitis to test the efficacy of EcN in acute and chronic experimental intestinal inflammation. The modified transfer model (26, 27), based on the CD4⫹CD45RBhighT-cell model, first described by Powrie
et al. (29), is characterized by a massive influx of mononuclear cells into the colonic mucosa, elevated levels of proinflamma-tory cytokines, and a chronic, severe wasting syndrome. The degree of colitis in this model correlates with the extent of luminal bacterial growth (1) and therefore makes it suitable for experimental probiotic therapy. On the other hand, adminis-tration of DSS in drinking water induces acute colitis in mice (10) with patchy mucosal damage, including focal crypt loss, followed by acute transmural infiltration with inflammatory cells. It has been shown that neither T nor B cells are required for the induction of acute DSS-induced colitis, as this model can also be used for SCID mice (13). However, DSS colitis also seems to be dependent on the intestinal microflora (2, 3, 31).
FIG. 4. (A) Histological appearance of colitis induced by transfer of naïve CD4⫹CD62L⫹T lymphocytes in SCID mice. The colitis is
characterized by a thickening of the mucosa due to edema and an influx of inflammatory cells (magnification,⫻100). (B) Administration of EcN led to a significant reduction of the severity of chronic colitis in this model (magnification,⫻100). (C) Histological score of the colonic inflammation in mice following the induction of colitis by transfer of naïve CD4⫹CD62L⫹T lymphocytes and administration of EcN
com-pared to that of untreated colitic mice. *,P⬍0.02 versus CD62L.
on August 17, 2020 by guest
The use of the probiotic microorganism EcN in SCID mice following transfer of CD4⫹CD62L⫹T cells ameliorated the development of intestinal inflammation, as was demonstrated by a healthier clinical appearance of the animals, lower histo-logical scores, and changes in cytokine secretion profiles of mesenteric lymphocytes. A marked reduction in the secretion of the proinflammatory cytokine IFN-␥was observed, which is thought to contribute predominantly to the perpetuation of intestinal inflammation, as seen in many previous murine mod-els of colitis (14). Additionally, MLN cells from EcN-treated colitic SCID mice secreted less IL-5 and IL-6. IL-6 seems to play a major role in intestinal inflammation, since blockage of
the IL-6 receptor in a murine model of colitis successfully prevented both wasting disease and the development of mac-roscopic and histologic lesions (16). On the other hand, there was no effect on the secretion of IL-10, an antiinflammatory cytokine secreted predominantly by regulatory T cells.
Both the CD4⫹CD45RBhighand the CD4⫹CD62L⫹ mod-els are characterized by chronic intestinal inflammation and a
FIG. 5. Reduction of proinflammatory cytokine secretion of MLN following the induction of colitis by transfer of naïve CD4⫹CD62L⫹T
lymphocytes and administration of EcN compared to the results seen with untreated colitic mice. Values given are means⫾ SEM (pico-grams per milliliter). (A) IFN-␥(*,P⬍0.03); (B) IL-6 (*,P⫽0.02); (C) IL-5 (*,P⫽0.02). AllPvalues shown are versus those of CD62L.
FIG. 6. Total concentration ofE. coli-like colonies in cecal content from mice from the transfer model following oral administration of EcN for 8 weeks compared to that of untreated colitic mice. *,P⬍
0.001 versus CD62L.
FIG. 7. Evidence of bacterial translocation into MLN. Total con-centration ofE. coli-like colonies in MLN of mice from the transfer model following oral administration of EcN for 8 weeks compared to that of untreated colitic mice. EcN was identified by REP-PCR. *,P⬍
0.0001 versus CD62L.
on August 17, 2020 by guest
http://cvi.asm.org/
acteristic patchy pattern with noninflamed mucosa appearing next to necrotic tissue. However, the dominant parameter for the histology score in the acute phase is destruction of the mucosal architecture rather than the influx of inflammatory cells, which is most likely due to a primarily toxic effect of DSS. The beneficial effect of probiotic administration, however, is obviously limited to the immunologic modification of colitis, as demonstrated by a pronounced reduction of proinflammatory cytokine secretion, which is more relevant in the chronic phase of intestinal inflammation (13).
The microbial analysis of cecal contents of mice from the transfer model revealed that after 8 weeks of oral administra-tion, EcN did not replace the residentE. coli population but rather led to an increase in the total E. coli concentration. Furthermore, subcultures of homogenized MLN showed a translocation of viable members of Enterobacteriaceae. Via REP-PCR, these colonies were identified as resident Enter-obacteriaceaefrom the intestinal flora and as colonies from the orally administered EcN. This finding is even more interesting in context with our observation that bacteria could not be isolated from MLN derived from healthy BALB/c mice follow-ing administration of EcN for 8 weeks.
Intestinal inflammation leading to a breakdown of the intes-tinal epithelial barrier function that would subsequently allow microorganisms from within the gut lumen to translocate to mucosal lymphatic tissue was always suggested (4, 19, 45). There are previous reports that bacterial translocation occurs also in healthy mice but is limited to 10 to 20% of mice analyzed (5, 44). In as yet unpublished observations (for which healthy BALB/c mice were used), we were able to detect a peak concentration of translocated microorganisms within MLN and Peyer’s patches at 6 to 12 h post-oral administration, followed by a steady decline until no bacteria could be isolated from lymphoid tissue at 48 h post-oral administration (unpub-lished data).
The fact that neither translocated EcN nor translocated res-identE. colibacteria could be isolated from MLN of healthy BALB/c mice after 8 weeks of treatment confirms the hypoth-esis that under healthy conditions only a minor translocation of luminal intestinal bacteria to mesenterical lymphatic tissue takes place. These are then cleared rapidly by phagocytosis (45). However, under inflammatory conditions, the epithelial barrier is disrupted, and luminal bacteria, including the orally administered EcN, are able to continuously translocate in very large quantities to Peyer’s patches and MLN, as seen in the
fying intestinal immunologic responses which result in the amelioration of chronic experimental murine colitis but have no clinical effect on the severity of acute mucosal damage. These findings are in accordance with clinical observations in which probiotic therapy was beneficial only in the maintenance of remission of chronic pouchitis and ulcerative colitis but had no effect for acute IBD. However, despite the successful use of probiotic therapy in cases of human pouchitis and ulcerative colitis, our suggestions of possible underlying antiinflammatory mechanisms, deployed by probiotic bacteria, cannot be fully extrapolated to the human situation since these experiments were performed under standardized conditions that are not feasible for human trials.
The analysis of intestinal microflora in this experimental setting was limited toEnterobacteriaceae, and the impact on the composition of the microflora by probiotic therapy therefore remains elusive. Note that no final conclusion can be drawn regarding a direct impact of EcN on cytokine production that reduces inflammatory activity as demonstrated previously in other studies (35, 41). On the other hand, an alteration in cytokine levels because of reduced inflammatory activity that is mediated by a modulation of the gut flora can also be antici-pated (42). However, as demonstrated in this report, EcN is able to translocate into MLN and to modulate the proinflam-matory cytokine secretion profile of mesenterial lymphocytes in both models of murine colitis, without significant ameliora-tion of the acute DSS colitis. It can therefore be speculated that EcN directly modulates the immune response.
ACKNOWLEDGMENTS
We thank Ardeypharm, Herdecke, Germany for providing E. coli strain Nissle 1917.
M.S. is supported by a research grant from the University of Re-gensburg, ReRe-gensburg, Germany, as part of the ReForM program.
REFERENCES
1. Aranda, R., B. C. Sydora, P. L. McAllister, S. W. Binder, H. Y. Yang, S. R. Targan, and M. Kronenberg.1997. Analysis of intestinal lymphocytes in mouse colitis mediated by transfer of CD4⫹, CD45RBhighT cells to SCID recipients. J. Immunol.158:3464–3473.
2. Axelsson, L. G., E. Landstro¨m, A. C. Bylund-Fellenius, and T. Midtvedt. 1997. Dextran sulfate sodium induced colitis and sulfasalazine treatment in bacteria free mice. Gastroenterology108:A775.
3. Axelsson, L. G., T. Midtvedt, and A. C. Bylund-Fellenius.1996. The role of intestinal bacteria, bacterial translocation and endotoxin in dextran sodium sulphate-induced colitis in the mouse. Microb. Ecol. Health Dis.9:225–237. 4. Baumgart, D. C., and A. Dignass.2002. Intestinal barrier function. Curr.
Opin. Clin. Nutr. Metab. Care5:685–694.
5. Berg, R. D.1983. Bacterial translocation from the gastrointestinal tracts of
on August 17, 2020 by guest
mice receiving immunosuppressive chemotherapeutic agents. Curr. Micro-biol.8:285–292.
6. Bienenstock, J., R. E. Riley, G. S. Neigh, S. Waserman, and P. Keith.2002. Probiotics in the management and prevention of atopy. Clin. Rev. Allergy Immunol.22:275–285.
7. Borruel, N., M. Carol, F. Casellas, M. Antolin, F. de Lara, E. Espín, J. Naval, F. Guarner, and J. R. Malagelada.2003. Increased mucosal tumor necrosis factor␣production in Crohn’s disease can be downregulated ex vivo by probiotic bacteria. Gut51:659–664.
8. Boudeau, J., A. L. Glasser, S. Julien, J. F. Colombel, and A. Darfeuille-Michaud.2003. Inhibitory effect of probioticEscherichia colistrain Nissle 1917 on adhesion to and invasion of intestinal epithelial cells by adherent-invasiveE. colistrains isolated from patients with Crohn’s disease. Aliment. Pharmacol. Ther.18:45–56.
9. Cong, Y., C. T. Weaver, A. Lazenby, and C. O. Elson.2002. Bacterial-reactive T regulatory cells inhibit pathogenic immune responses to the enteric flora. J. Immunol.169:6112–6119.
10. Cooper, H. S., S. N. S. Murthy, R. S. Shah, and D. J. Sedergran.1993. Clinicopathologic study of dextran sulfate sodium experimental murine co-litis. Lab. Investig.69:238–249.
11. Cremonini, F., S. Di Caro, E. C. Nista, F. Bartolozzi, G. Capelli, G. Gas-barrini, and A. Gasbarrini.2002. Meta-analysis: the effect of probiotic ad-ministration on antibiotic-associated diarrhea. Aliment. Pharmacol. Ther. 16:1461–1467.
12. Dieleman, L. A., M. S. Goerres, A. Arends, D. Sprengers, C. Torrice, F. Hoentjen, W. Grenther, and R. B. Sartor.2003.LactobacillusGG prevents recurrence of colitis in HLA-B27 transgenic rats after antibiotic treatment. Gut52:370–376.
13. Dieleman, L. A., B. U. Ridwan, G. S. Tennyson, K. W. Beagley, R. P. Bucy, and C. O. Elson.1994. Dextran sulfate sodium-induced colitis occurs in severe combined immunodeficient mice. Gastroenterology107:1643–1652. 14. Elson, C. O.2000. Commensal bacteria as targets in Crohn’s disease.
Gas-troenterology119:254–257.
15. Isolauri, E.2003. Probiotics for infectious diarrhea. Gut52:436–437. 16. Ito, H.2003. Anti-interleukin-6 therapy for Crohn’s disease. Curr. Pharm.
Des.9:295–305.
17. Kruis, W., P. Fric, M. Stolte, and the Mutaflor Study Group.2001. Main-tenance of remission in ulcerative colitis is equally effective withEscherichia coliNissle 1917 and with standard mesalamine. Gastroenterology120:A680. 18. Kruis, W., E. Schutz, P. Fric, B. Fixa, G. Judmaier, and M. Stolte.1997. Double-blind comparison of an oralEscherichia colipreparation and me-salazine in maintaining remission of ulcerative colitis. Aliment. Pharmacol. Ther.11:853–858.
19. Kucharzik, T., N. Lugering, K. Rautenberg, A. Lugering, M. A. Schmidt, R. Stoll, and W. Domschke.2000. Role of M cells in intestinal barrier function. Ann. N. Y. Acad. Sci.915:171–183.
20. Lammers, K. M., U. Helwig, E. Swennen, F. Rizzello, A. Venturi, E. Cara-melli, M. A. Kamm, P. Brigidi, P. Gionchetti, and M. Campieri.2002. Effect of probiotic strains on interleukin-8 production by HT-29/19A cells. Am. J. Gastroenterol.97:1182–1186.
21. Lilly, D. M., and R. H. Stilwell.1965. Probiotics: growth promoting factors produced by microorganisms. Science47:747–748.
22. Mack, D. R., S. Michail, S. Wei, L. McDougall, and M. A. Hollingsworth. 1999. Probiotics inhibit enteropathogenicE. coliadherence in vitro by in-ducing intestinal mucin gene expression. Am. J. Physiol.276:G941–G950. 23. Madsen, K. L., A. Cornish, P. Soper, C. McKaigney, H. Jijon, C. Yachimec,
J. Doyle, L. Jewell, and C. De Simone.2001. Probiotic bacteria enhance murine and human intestinal epithelial barrier function. Gastroenterology 121:580–591.
24. Madsen, K. L., J. S. Doyle, L. D. Jewell, M. M. Tavernini, and R. N. Fedorak. 1999.Lactobacillusspecies prevents colitis in interleukin 10 gene-deficient mice. Gastroenterology116:1107–1114.
25. Malin, M., H. Suomalainen, M. Saxelin, and E. Isolauri.1996. Promotion of IgA immune response in patients with Crohn’s disease by oral bacterio-therapy withLactobacillusGG. Ann. Nutr. Metab.40:137–145.
26. Mudter, J., S. Wirtz, P. R. Galle, and M. F. Neurath.2003. A new model of chronic colitis in SCID mice induced by adoptive transfer of CD62L⫹CD4⫹ T cells: insights into the regulatory role of interleukin-6 on apoptosis. Patho-biology70:170–176.
27. Neurath, M. F., B. Weigmann, S. Finotto, J. Glickmann, E. Nieuwenhuis, H.
Iijima, A. Mizoguchi, E. Mizoguchi, J. Mudter, P. R. Galle, A. Bhan, F. Autschbach, B. M. Sullivan, S. J. Szabo, L. H. Glimcher, and R. S. Blum-berg.2002. The transcription factor T-bet regulates mucosal T cell activation in experimental colitis and Crohn’s disease. J. Exp. Med.195:1129–1143. 28. O¨ lschla¨ger, T., A. Altenho¨fer, and J. Hacker.2001. Inhibition ofSalmonella
typhimuriuminvasion into intestinal cells by the probiotic strainE. coliNissle 1917. Gastroenterology120:A1682.
29. Powrie, F., M. W. Leach, S. Mauze, L. B. Caddle, and R. L. Coffman.1993. Phenotypically distinct subsets of CD4⫹T cells induce or protect from chronic intestinal inflammation in C. B-17scidmice. Int. Immunol.5:1461– 1471.
30. Powrie, F., and D. Mason.1990. OX-22highCD4⫹T cells induce wasting disease with multiple organ pathology: prevention by the OX-22lowsubset. J. Exp. Med.172:1701–1708.
31. Rath, H. C., M. Schultz, R. Freitag, L. A. Dieleman, F. Li, H.-J. Linde, J. Schoelmerich, and R. B. Sartor.2001. Different subsets of enteric bacteria induce and perpetuate experimental colitis in rats and mice. Infect. Immun. 69:2277–2285.
32. Rath, H. C., H. H. Herfarth, J. S. Ikeda, W. B. Grenther, T. E. Hamm, Jr., E. Balish, J. D. Taurog, R. E. Hammer, K. H. Wilson, and R. B. Sartor.1996. Normal luminal bacteria, especiallyBacteroides species, mediate chronic colitis, gastritis, and arthritis in HLA-B27/human2 microglobulin trans-genic rats. J. Clin. Investig.98:945–953.
33. Rembacken, B. J., A. M. Snelling, P. M. Hawkey, D. M. Chalmers, and A. T. Axon.1999. Non-pathogenicEscherichia coliversus mesalazine for the treat-ment of ulcerative colitis: a randomised trial. Lancet354:635–639. 34. Sartor, R. B.2002. Microbial agents in the pathogenesis, differential
diag-nosis, and complications of inflammatory bowel disease, p. 435–458.InM. J. Blaser, P. D. Smith, and J. I. Ravdin (ed.), Infections of the gastrointestinal tract. Raven Press, Ltd., New York, N.Y.
35. Schultz, M., H.-J. Linde, N. Lehn, K. Zimmermann, J. Grossmann, W. Falk, and J. Schoelmerich.2002. Immunomodulatory consequences of oral ad-ministration ofLactobacillus rhamnosusstrain GG (L. GG) in healthy vol-unteers. J. Dairy Res.70:165–173.
36. Schultz, M., J. Schoelmerich, and H. C. Rath.2003. Rationale for probiotic and antibiotic treatment strategies in inflammatory bowel disease. Dig. Dis. 21:105–128.
37. Schultz, M., C. Veltkamp, L. A. Dieleman, R. B. Wyrick, S. L. Tonkongy, and R. B. Sartor.2002.Lactobacillus plantarum299v in treatment and prevention of spontaneous colitis in IL-10-deficient mice. Inflamm. Bowel Dis.8:71–80. 38. Schultz, M., S. L. Tonkonogy, R. K. Sellon, C. Veltkamp, V. L. Godfrey, J. Kwon, W. B. Grenther, E. Balish, I. Horak, and R. B. Sartor.1999. IL-2-deficient mice raised under germfree conditions develop delayed mild focal intestinal inflammation. Am. J. Physiol.276:G1461–G1472.
39. Sellon, R. K., S. L. Tonkonogy, M. Schultz, L. A. Dieleman, W. B. Grenther, E. Balish, D. Rennick, and R. B. Sartor.1998. Resident enteric bacteria are necessary for development of spontaneous colitis and immune system acti-vation in interleukin-10-deficient mice. Infect. Immun.66:5224–5231. 40. Tannock, G. W., K. Munro, H. J. Harmsen, G. W. Welling, J. Smart, and
P. K. Gopal.2000. Analysis of the fecal microflora of human subjects con-suming a probiotic product containingLactobacillus rhamnosusDR20. Appl. Environ. Microbiol.66:2578–2588.
41. Ulisse, S., P. Gionchetti, S. D’Alo`, F. P. Russo, I. Pesce, G. Ricci, F. Rizzello, U. Helwig, M. G. Cifone, M. Campieri, and C. De Simone.2001. Expression of cytokines, inducible nitric oxide synthetase and matrix metalloproteinases in pouchitis: effects of probiotic treatment. Am. J. Gastroenterol.96:2691– 2699.
42. Venturi, A., P. Gionchetti, F. Rizzello, R. Johansson, E. Zucconi, P. Brigidi, D. Matteuzzi, and M. Campieri.1999. Impact on the composition of the faecal flora by a new probiotic preparation: preliminary data on maintenance treatment of patients with ulcerative colitis. Aliment. Pharmacol. Ther.13: 1103–1108.
43. Versalovic, J., T. Koeuth, and J. R. Lupski.1991. Distribution of repetitive DNA sequences in eubacteria and application to fingerprinting of bacterial genomes. Nucleic Acids Res.19:6823–6831.
44. Wells, C. L., M. A. Maddaus, and R. L. Simmons.1987. Role of the mac-rophage in the translocation of intestinal bacteria. Arch. Surg.122:48–53. 45. Wiest, R., and H. C. Rath.2003. Bacterial translocation in the gut. Best
Pract. Res. Clin. Gastroenterol.17:397–425.