RECOMBINATION BETWEEN COMPONENTS OF A
MUTABLE GENE SYSTEM IN MAIZE
JERRY L. KERMICLE
Laboratoy of Genetics, University of Wisconsin, Madison, Wisconsin 53706 Manuscript received November 29, 1982
Revised copy accepted March 15, 1984 ABSTRACT
An unstable component (Z-R) of the R-stippled allele interacts with a linked modifier which enhances stippled’s expression (M-st) to delete the intervening segment. The precision of deletion formation suggests a recombinational basis, specifically unequal crossing over between I-R and M-st. Four deletions were selected as losses of R function from plants homozygous for R-st and M-st. Also lost are stippled’s near-colorless seed phenotype, its paramutagenicity and a closely linked gene, Inhibitor of striate. The deletion chromosomes, missing a 6-cM segment, are transmitted normally by ovules but in reduced frequency by pollen. Homozygous and heteroallelic combinations of the deletions confer defective seed lethality. The four did not differ detectably in transmissibility or breakpoint termini. The recurrence of deletions that have the same termini is explained by recombination between Z-R and M-st. The homology between M-st and Z-R, and their presence in the same chromosome arm, favors the view that M-st originated by Z-R transposition.
WO components underlie the dense pattern of kernel spotting (stippling)
T
commonly found among races of maize indigenous to the Andean High- lands in South America. At the R locus, a unit capable of conferring strong seed coloration (Sc) is expressed irregularly due to a tightly linked inhibiting factor. T h e inhibitor (Z-R) destabilizes (Sc) or other R tissue-specific deter- miners coupled to it through crossing over only in cis linkage phase (ASHMAN1970; KERMICLE 1970). A second component, mapping six units distal, en- hances spotting intensity. Action of this modifier (M-st) is dominant, increases with dosage and is trans active (ASHMAN 1960). Evidence presented in the accompanying paper (WILLIAMS, SATYANARAYANA and KERMICLE 1984) dem- onstrates the transposability of Z-R. T h e finding that I-R often transposed to linked positions, and that it modified R-st expression in the same manner as
M-st, suggests that M-st originated as a transposed I-R.
T h e foregoing effects of Z-R and M-st on stippled were revealed as functional interactions. T h e present paper reports evidence indicating a structural relation between I-R and M-st. Homology between the two predispose chromosomes 10 carrying them to recombine, forming deletions that extend precisely from I-R
to M-st. Thus, the two elements behave as duplicated segments, displaced but oriented in the same direction, which undergo unequal crossing over. T h e deletions were identified initially as single-kernel variants that were colorless
490 J. L. KERMICLE TABLE 1
Classijcation of R alleles in terms of distinguishing pigmenting action, probable genic element constitution and number of Isr (Inhibitor of striate-2) units
Pigmenting action
R allele Aleurone Anthers
No. of Isr units
R-st R-sc r-g:nc3-5 r-g:Sd r-g:e r-g:de r-xl R-r:std r-r:n I9 r-r:n 35 r-r:n I O 1
r-r:W22 R-g: 1 R-g:8 pale
Spotted Solid Faint“ Colorless Colorless Colorless Colorless Solid’ Colorless Colorless Colorless Colorless Solid’ Pale Acyanic Acyanic Ac yanic Ac yanic Acyanic Acyanic Acyanic Red Red Red Red Red Acyanic Acyanic 1 + 1 , 1+ N T c 1 0 0 0 2 1 2 2 0 1 N T Genic element composition (Sc)I-R( Nc) (SC), (Sc)(Nc) (Sc)l-R?(Nc) Unknown Unknown Deletion Deletion (P)(S) (P) (P)(S) (P)(S) (P) (S)
Modified (S) a Incompletely penetrant.
’
R R R and R R r aleurone is solidly colored; r r R is darkly mottled. ‘ Not tested.rather than stippled. T h e variants are characterized here in terms of pheno- type, transmission and deletion termini.
MATERIALS AND METHODS
Various test genotypes were synthesized as combinations of R alleles and marker genes that first had been incorporated by backcrossing into sublines of inbred W22 which carry the complemen- tary genes needed for anthocyanin pigmentation. The following descriptions pertain to the single- character stocks. A summary of distinguishing effects is given in Table 1.
R alleles: Clones of intensely pigmented cells spot the aleurone layer of the endosperm and the scutellum of the embryo in kernels carrying the R-stippled allele. The spotting pattern is dense when the linked modifier M-st is present. In the absence of the modifier, i.e., R-st m-st, the pattern appears coarser and less dense. R-st reverts germinally to a stable, uniformly colored kernel phe- notype termed self-colored (R-sc). Neither R-st nor R-sc pigment vegetative parts of the adult plant, although the primary leaf tip of R-sc seedlings is red.
ASHMAN (1965) described a second seed-pigmenting unit in R-st whose effects are most clearly seen in the absence of the dark spotting. Cells showing this “near-colorless” phenotype are weakly colored and are interspersed with colorless in a mottled pattern. The (Nc) component maps distal to I-R in the R-st complex; the linear order is centromere - (Sc) I-R (Nc). The particular source of R-st used in the present investigation is the same as that reported previously for various fine structural and paramutational studies at the University of Wisconsin.
r-g:Sd designates a colorless seed, green plant allele of r introduced in a multiple recessive stock obtained from L. J. STADLER. I t is phenotypically null in R expression and is stable.
P - - -
r - x i
r-g:de
I
R-Lc H
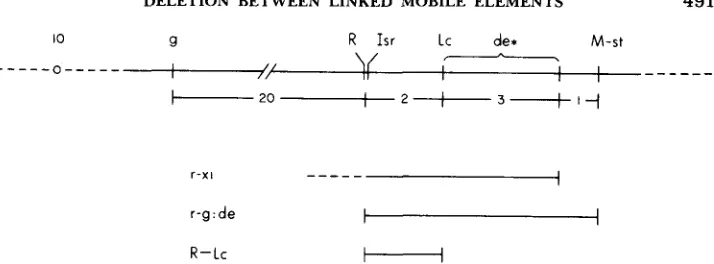
FIGURE 1.-Genetic map of the long arm of maize chromosome 10 indicating R, flanking markers and the segments spanned by three deletions. de* symbolizes a recessive lethal, defective endosperm phenotype associated with deletions r-xl and r-g:de but not with R-Lc.
aleurone stock is revealed by the phenotype of FI kernels produced from a cross with r-g:nc?-5 m- st. R. B. ASHMAN isolated r-g:nc?-5 from a homozygous R-st m-st line and proposed its use as an M-st tester.
The r-g:de variants here under analysis, 4-128, 4-137, 4-139 and 4-142, were isolated by R. A. BRINK and J. D. AXTELL as single, colorless kernel selections from large scale testcrosses of homo- zygous R-st M-st plants by r-r/r-r males. The R-st M-st/R-st M-st parent also carried the variegated pericarp allele, P - w , in heterozygous combination with colorless pericarp, P-wr. Three of the four plants grown from the selected kernels lacked P-vu and tested negative for the presence in the genome of Modulator, the transposable component of P-vu. A homozygous P-wr subline of the fourth isolate (4-139) was derived for present use. Phenotypically, the four variants have lost stippled's near-colorless mottling as well as the intense spotting. When homozygous, each confers seed abortion at an early stage of kernel development. The symbol r-g:de denotes this "defective seed" phenotype. Additional features of the four r-g:de isolates are presented in RESULTS.
r-g:e is a colorless seed, green plant accession which behaves recombinationally and in interaction with chlorophyll-striping genes as a local deficiency for R and a closely linked inhibitor of chlo- rophyll striping, Zsr.
r-xZ is an X-ray-induced deletion of R and Zsr isolated by STADLER. It belongs to the r-g phenotypic class and is not pollen transmitted (STYLES and BRINK 1969).
R-rcstandard has separate seed (S) and plant (P) components organized as members of a direct duplication, with (P) proximal to (S) (DOONER and KERMICLE 1974).
r-r:nl9 descends from R-ntandard. It carries (P) but not (S) or the duplication.
r-r:n35 and r-r:nlOZ also descend from R-rstandard. Both retain (P) and the duplication but r-r:W22 designates the colorless seed, red plant allele of R carried in the commercial form of R-g:Z descends from R-mtandard. It retains (S) but not (P) or the duplication.
R-g:8 pale was isolated as a pale-seeded variant of R-g:8, another (S) derivative of R-mtandard. Linked markers (see Figure I ) : Homozygous golden-1 plants are yellowish green. The g locus lies centromeric to R, recombining with it in the background of inbred W22 with a frequency of approximately 20%.
Zsr, an inhibitor of certain recessive chlorophyll-striping mutations such as striate-2, lies imme- diately distal to R. It is encompassed within the duplicated segment of the R-rstandard chromo- some, is carried but once in derivatives such as r-r:nl9 and R-g:Z and is absent or null in r-g:e (KERMICLE and AXTELL 1981).
Lc, an anthocyanin leaf color factor, isolated from Ecudorian strain 1172, maps one to two units distal to R. It is represented in a recombinationally short, displaced duplication of R which includes Isr. Unequal crossing over between R and Le yields R to Lc duplicate and deficient products (DOONER and KERMICLE 1976).
Unlinked markers: Wc* designates a dominant factor for white capping of the endosperm that is not (S) function.
J. L. KERMICLE
0
o x
A
D
* *
I I I I I I
40 50 60 70 80 90
LEAF
WIDTH
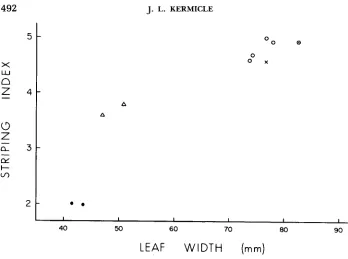
(mm)FIGURE 2.-Chlorophyll striping in homozygous striate-2 plants of different R locus constitution. R-st strains (A) are intermediate between the weakly striped, narrow-leafed r-r strains (0) that carry the R region in duplicate and the extensively striped, broad-leafed strains that carry the null or deficient alleles, r-g:e (0) and r-xl (X). The four r-g:de variants of R-st are indicated by open circles. Each of the R alleles was evaluated in combination with r-g:e.
epistatic to yellow. When carried homozygously in male parents, it served to identify kernels derived from other than the intended source of pollen. The asterisk signifies that the allelic relationship of this factor to others of similar phenotype is not known.
wx designates the recessive endosperm mutation waxy. Located in chromosome 9, it was used also as a pollen marker.
Large scale testcrossing: The large populations of kernels needed for intralocus recombination analysis (Tables 4 and 5 ) were produced in isolated plots where detasseled test heterozygotes were wind pollinated by interplanted male rows. The male parent combined either r-g or R-g:8 pale with the linked marker genes g and m-st (absence of M-st) as well as unlinked wx. Colored kernels, taken as putative recombinants, were progeny tested for heritability of the selected phenotype, classified for golden:green plant, crossed with r-g:nc3-5 for M-st:m:st evaluation and either self- pollinated or crossed by a Wx wx heterozygote to confirm the intended paternity.
Evaluation of paramutation: The seed-pigmenting potential of many R alleles, including R- mtandard and R-g:I, is heritably reduced in heterozygous combination with certain other alleles, among which R-stippled and its derivatives have been most extensively studied (see review by BRINK 1973). The extent of reduction is determined by evaluating kernel-pigmentating action of R alleles from control homozygotes and test heterozygotes in crosses on r r. The resulting kernels, of r r R aleurone genotype, were evaluated here in the manner detailed by BRINK et al. (1960), whereby test samples are graded against a set of six reference kernels, defining seven color classes (class 1 = colorless; class 7 = full colored).
* *
x50 60 70 80
LEAF
WIDTH
(mm)
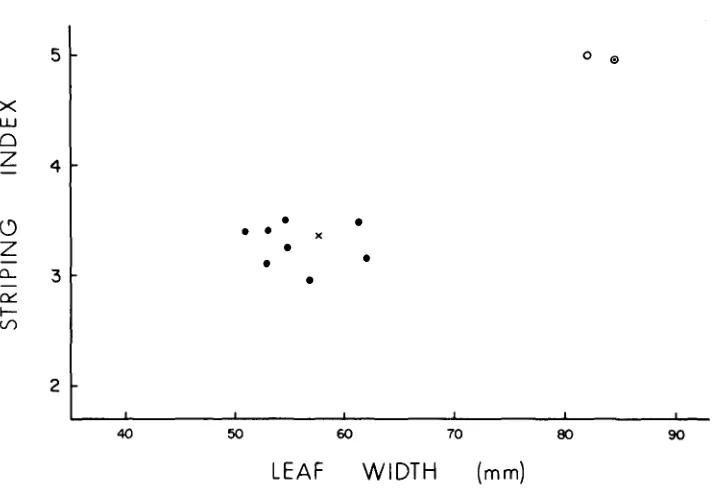
FIGURE 3.-Modification of chlorophyll striping by r-r:nl9 (X), r-g:deZ28 (0) and by eight of the R-sc derivatives (0) obtained from heterozygous combination of the two alleles. Each of the alleles was tested in combination with r-g:e. Homozygous r-g:e is indicated by (0).
RESULTS
Transmission and complementation of the r-g:de variants: Part a of Table
2
reports the incidence of r-g:de transmission by pollen. Tested as r-g:de/R-g
heterozygotes, the four r-g:de isolates accounted for 36.6-38.9% of the pollen grains effecting fertilization. The deviations from the Mendelian expectation of 50% are highly significant, with no evidence of significant heterogeneity between variants (P = 0.6). A comparable level of reduced r-g:de transmission through ovules would be expected to yield irregularly filled ears. Ovule abor- tion above the ordinary background level, however, was not observed. Neither did the ratios deviate appreciably from 50% in 1000-kernel samples of the two
r-g:de cases tested (part b, Table 2).
Self-pollination of a set of r-g:de/R-st heterozygotes revealed an additional feature of r-g:de expression. Each ear bore a nonviable class of kernels that had failed at an early stage of seed development. All of the plump seeds, furthermore, were stippled, except for an occasional self-colored (R-sc) rever- tant. The absence of a colorless class in the presence of an abortive kernel class indicates recessive lethality of r-g:de homozygotes.
494 J. L. KERMICLE TABLE 2
Transmission of r-g:de variants in reciprocal crosses of r-g:de/R heterozygotes with r/r
No. of kernels
r-g:de isolate Colored Colorless 76 colorless
(a) r / r X r-g:de/R-g
4-128 995 630 38.8
4-137 723 418 36.6
4-1 39 1084 674 38.3
4-142
-
Total
- 1054 670
3856 2392
- 38.9
38.0
-
(b) r-g:de/R-st X r / r
4-128 507 493 49.3
4-137 507 493 49.3
seed (stippled with occasional R-sc) in accordance with those obtained from self-pollination. An absence of complementation was regularly observed also in crosses of plants in the four r-g:de/R-st progenies to ones carrying the R-
deficient chromosome r - x l . Crosses of genotype r - x l / R - r female X r-g:de/R-st
male yielded plump seed comprised of uniformly colored (R-r carrying) and stippled (r-xl/R-st) but not a colorless class corresponding to r-xllr-g:de.
Rather, a class of severely defective seeds was present on each ear.
Paramutagenicity of R-st and its r-g:de variants: Table 3 gives data comparing the effect of R-st and its r-g:de variants on R-g:l, on allele that is sensitive to
stippled’s paramutagenic action. In testcrosses on r-g:Sd/r-g:Sd females of eight
R-g R-g control plants, aleurones of the resulting r-g r-g R-g endosperms reg- ularly were darkly mottled, scoring as class 6. Kernels of the same genotype produced by testcrossing R-g/R-st plants scored from 3.43 to 3.87 in the four families studied. These means reflect the weakly pigmented phenotypes char- acteristic of an allele that is sensitive to the pigment-reducing action of R-st in heterozygotes.
Each group of R-g/R-st plants was accompanied by r-g:de/R-g sibs, both gen- otypes descending from the cross r-g:de/R-st X R-g R-g. Testcross r-g r-g R-g
kernels from R-glr-g:de plants representing the four r-g:de variants were either darkly mottled, class 6, or, occasionally, uniformly pigmented, class
7
(Table 3). Thus, all four variants differed categorically from the progenitor allele R - st by not reducing the pigmenting potential of R-g:l in heterozygotes.Modification of chlorophyll striping: T h e data pictured in Figure 2 test whether the four r-g:de variants differ from the progenitor allele R-st in interaction with the chlorophyll-striping gene striate-2. KERMICLE and AXTELL (1 98 1) found the amount of albino tissue and the width of leaf blades in sr2 sr2 plants
TABLE 3
Aleurone color scores of r-g r-g R-g kernels obtained in testcrosses of
sib R-g/R-st and R-g/r-g:de pollen parents on r-g r-g 99
Aleurone color score‘
r-g:da isolate R-g/R-st R-glr-g:de
4-128 3.87 (8)* 6.27 (7)
4-137 3.71 (7) 6.08 (5)
4-139 3.76 (9) 6.04 (7)
4- 142 3.43 (5) 6.12 (8)
The two classes of plants compare the paramutagenic effect of
@ Aleurone score of eight R-g R-g control plants testcrossed on
R-st and its r-g:de variants on R-g.
r-g r-g = 6.00.
Number of testcross ears is given in parentheses.
TABLE 4
Number and linked marker composition of the self-colored variants obtained from
R-stir-r:n 19 heterozygotes and from hemizygotes of R-st with the r deficient chromosomes
r-xl and r-g:e
~ ~~
NO. of R-sc variants according to flanking marker composition
-
Genotype R-st gametes
- - L A -
17,410 21 0 8 7
35,940 51 0 1 10
19,330 25 0 0 6
G R-st LC
g r-r:nl9 IC
G r-xl IC g R-st LC
g R-st LC
G r-p:e IC
The male parent for the test crosses was g r-g IC.
of r-r:n35 and r-r:nlOl. These r-r alleles represent chromosomes in which the
R locus and the closely linked gene Inhibitor of striate are duplicated. Corre- sponding values for the four r-g:de test stocks are seen to plot not with R-st
but near the r-g:e and r-xl points. Thus, the change from R-st to r-g:de is associated with loss of the factor that inhibits expression of sr2.
Proximal breakpoint of the r-g:de variants: Tables 4 and 5 compare the re- version frequency to self-colored of R-st and its r-g:de derivatives when heter- ozygous with a colorless seed but colored plant allele, r-r:nl9. When homo- zygous, this r-r allele gave no colored seed in 42,800 gametes tested (DOONER and KERMICLE 1974).
A majority class of the R-sc’s obtained from R-st/r-r:n19 plants was borne by chromosomes carrying the flanking markers of parental R-st (Table 4). If
496 J. L. KERMICLE TABLE 5
Number and linked marker composition of self-colored derivatives obtained from G r-g:de/g r-r:nl9 heterozygotes in crosses by g r-g and g R-g:8 pale males
No. of R-sc's according to linked marker class
G R-sc de" g R-sc -k G R-sc -k g Rsc de"
I -L 1
r-g:de isolate r-g:de gametes
-
4-128 30,580 0 1 27 0
4-137 7,330 0 0 12 0
4-1 39 8,180 0 3 14 0
4-142 26,240 0 0 33 0
Tested either by reduced pollen transmission of the R-sc-bearing chro- mosome or the presence of defective kernels on selfed ears.
recessive allele of golden (20 units proximal) which entered the cross with r-
r. Seven were found. Two percent or only 0.7 cases should have carried the distal marker ( I C ) of r-T, by similar reasoning. Eight, however, were observed. An excess over expectation of self-coloreds belonging to this recombinant class was reported previously from other R-st heterozygotes (KERMICLE 1970). T h e excess was attributed to the origin of some R-sc variants by separation of a component of R-st which confers seed coloration (Sc) from an instability factor
(I-R) slightly distal. Thus, crossing over evidently separates I-R from ( S c ) and introduces a segment from the other allele, in this case r-r, which supports strong seed coloration.
A second entry in Table 4 reports the linked marker constitution of R-sc's
derived from R-st in combination with the r-deficient chromosome r-XI. If the excess of R-sc's bearing a particular recombinant class of flanking markers from
r-T heterozygotes was in fact d u e to crossing over, the excess should not be observed from r-xl/R-st hemizygotes. T h e number of cases bearing the prox- imal marker of r-r agrees with that predicted for a 20-unit interval (ten of 62
or 16.1 %). In this case, however, only one of the 62 R-sc's was an apparent recombinant for the distal marker. A third entry reports similar results involv- ing r-g:e, an allele believed to constitute a more localized deficiency for R .
T h e data in Table 5 report experiments testing whether r-r:n19 and r-g:de can recombine to yield R-sc. Whether the r-g:de variants, although conferring colorless seed, nevertheless retain the tissue-specific component of stippled's (Sc) element was the question posed. Each combination involving the four r-
g:de variants yielded self-colored variants. Of the 90 R-sc isolates, 86 were recombinant. They carried the G allele from r-g:de proximally but like r-r:n19
gave plump kernels when selfed and were transmitted normally by pollen. Four were marked as parental r-r:nl9. A likely origin for this minority class is recombination at R with a coincident exchange in the g to R interval. Were it
not for strong chiasma interference acting in this region (KERMICLE 1970), still more such cases would be expected. It is noteworthy that none of the 90 R-sc
TABLE 6
Kernels of stiffled phenotype obtained from heterozygotes of r-g:nc3-5 m-st with
r-g:de M-st and r-r:W22 M-st
Kernels
% recombina-
r allele Total Stippled tion
r-g:del28 3,360 1 0.06
r-g:de 139 5,390 2 0.07
0.08
25.450 7 0.06
r-g:de137 8,730 1 0.02
r-g:del42 7,970 - 3
-
r-r:W22 6,150 243 7.9
6 . 3
r-r:W22 7,050 223
13,200 466 7.1
-
In the foregoing test, the reduced pollen transmission and recessive lethal features of r-g:de were used routinely to classify distal marker constitution of the R-sc derivatives. Eight of the recombinant R-sc isolates obtained from r-
g:del28/r-r:n19 were also evaluated for Zsr. T h e r-r:n19 allele lacks the R-
r:standard duplication and, correspondingly, is seen to inhibit striate-2 action at a level that is intermediate between the duplication carrying r-r alleles reported in Figure 2 and the null effects of r-g:e and r-g:de. T h e values for the eight R-sc derivatives (Figure 3) surround r-r:n19, clearly separated from the permissive effect on striping of parental r-g:de128.
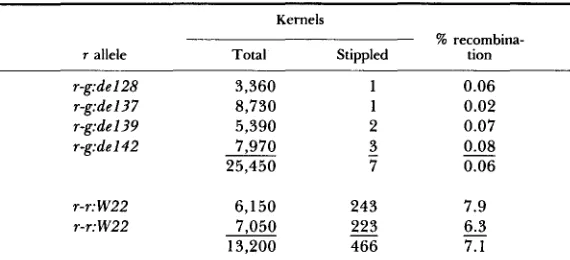
Distal breakpoint of the r-g:de variants: Experiments reported in Table 6 utilized M-st as a reference point for placing the distal breakpoint of the r-g:de
isolates. In the absence of M-st, the test allele r-g:nc3-5 expresses only the faint, near-colorless phenotype derived from its progenitor, R-stippled. In the pres- ence of M-st, either in cis or trans combination, the aleurone is intensely spotted, resembling stippled itself. Thus, kernels produced in the cross of r-
g:nc3-5 m-st with r-r:W22 M-st are stippled. Testcrosses of this heterozygote by
r-g:Sd males produced 466 stippled kernels among a population of 13,200. A stippled kernel results from this cross when r-g:nc3-5 and M-st recombine. T h e percent recombination, increased by a factor of two because only one crossover class is detected in this test, is 7.1. This value compares with an average of 5.7% for recombination between R and M-st reported by ASHMAN (1960). Parallel combinations involving r-g:nc3-5 with the four r-g:de M-st strains gave totals of only one to three stippled selections in populations ranging from 3360
to 8730 kernels, for an average recombination frequency of 0.06%. This value estimates the upper limit of recombination in that mechanisms other than crossing over, such as nondisjunction, which deliver M-st and r-g:nc3-5 to the same test gamete, would yield a stippled kernel.
498 J. L. KERMICLE
population of 107 1 r-xl gametes yielded 12 verified crossovers, indicating 1.1 % recombination between M-st and the distal breakpoint of r-xl.
DISCUSSION
Coincident change involving a number of different functions characterizes the family of variants reported here. Accompanying the loss of seed coloration, by which the r-g:de cases were isolated initially, was loss of the linked gene Inhibitor of striate, reduced pollen transmission, loss of stippled’s paramuta- genic property and the inception of recessive lethality. Recombination in the interval between R and M-st was eliminated or virtually so. These observations are consistent with the view that the variants originated by deletion of a chro- mosome segment extending from R to M-st.
T h e end points of the four deletions are decidedly nonrandom. T h e proxi- mal terminus of each is such that R function was eliminated without losing (Sc), the component which specifies strong seed color. In combination with an allele conferring plant but not seed color, (Sc) expression was regained in conjunction with the recombination of flanking markers. (Sc) expression was uniformly strong and in no instance reflected action of the instability factor I- R. This outcome is understandable if the deletions start either within the (Sc) to I-R interval or at I-R.
Comparable evidence concerning the distal terminus of the deletions derives from a consideration of the R-st modifier function. This function was retained by the r-g:de variants, although recombination between the modifier and R was virtually eliminated. T h e modifier in r-g:de behaves as though it is fused
T h e precision of deletion formation suggests the involvement of recombi- nation. Each of the four variants occurred as a separate, whole-kernel event as anticipated if homologous, meiotic recombination (albeit unequal) were causal. In particular, variants of the sort obtained are those expected from recombination between stippled’s cis-acting instability component I-R and the
trans modifier, M-st. Accordingly, M-st can be viewed as a displaced duplication that is capable of pairing and crossing over with I-R, forming duplicate and deficient products. Such an interaction has been described between the leaf color factor Lc and R (DOONER and KERMICLE 1976). Lc is situated two units distal to R in a recombinationally small segment which includes the locus of
Zsr.
An alternative mode of rearrangement is suggested by the unique charac- teristics of I-R and M-st. MCWHIRTER and BRINK (1962) pointed out the re- semblance between stippled’s instability and that of unstable systems in maize known to be based on transposable components. Various workers have re- ported instances in which R-st reversion to R-sc was associated with the occur- rence in the genome of a positive modifier of R-st expression. In a systematic study of this phenomenon, WILLIAMS, SATYANARAYANA and KERMICLE (1 984) relate the new modifiers to transposed I-R elements. T h e activity of the mod- ifiers ranged from barely perceptible up to a level equal to M-st. Some were linked with various intensities to R , whereas others assorted independently. M -
s t may also have originated as a linked, transposed I-R. If I-R and M-st respond to the same signals for excision, then simultaneous transposition o r an inter- active transposition involving both might lead to loss of the entire intervening region. T h e fact that a modifier function remains upon deletion formation is not inconsistent with this model. Retention of the mobile element at the orig- inal site appears to be a regular feature of transposition and deletion formation involving some bacterial insertion sequences and transposons (see reviews by STARLINCER 1980; CALOS and MILLER 1980). It is also possible that a hybrid unit composed of I-R and M-st components was formed at the deletion junc- tion.
A reciprocal product of the r-g:de variants reported here was not recovered. A complementary, duplication product, such as would be generated by unequal crossing over, should be flanked by R-st and M-st and encompass a hybrid R - st:M-st unit at the duplication junction. T h e phenotype of such a complex might not be readily distinguished from that of parental stippled. Duplication as well as deficient products are well known for the white locus of Drosophila
melanogaster. Recently, GOLDBERC et al. (1 983) reported finding a sequence that resembles the transposable element copia at the rearrangement breakpoints of duplications and deficiencies formed by unequal crossing over between the alleles white apricot and white apricot
4.
If crossing over were to occur intrachromosomally between directly oriented elements such as I-R and M-st, a ring chromosome bearing one of the elements might be produced. Although the resemblance of such a unit to transposable units identified in other organisms is evident, its origin and fate in the present connection is conjectural. T h e possibility that this product was present when the deletions first occurred but subsequently was lost by segregation cannot be excluded.
Analysis of the r-g:de alterations was facilitated by the r-g:nc3-5 tester strain. This stock, itself a derivative of R-st, expresses the nearly colorless phenotype of stippled but not its intense spotting unless M-st is also present in the genome. T h e r-g:nc3-5 variant can be viewed as a derivative of R-st in which I-R is transposition defective. In effect, an independent (autonomous) mutable system has become a two-element one, composed of a cis-acting receptor and a trans-
acting regulator unit. Parallel transformations are a well-known feature of the maize transposable elements Activator, Enhancer and Suppressor-Mutator (re- viewed by FINCHAM and SASTRY 1974).
T h e r-g:de deletions and the X-ray-induced deficiency r-xl overlap beginning at R and extending to the distal terminus of r - x l , about one unit proximal to
M-st (Figure 1). T h e locus of the recessive lethal effect seen in r-g:de homo- zygotes and r-g:de/r-xl combinations should lie within this region. It is placed in that portion distal to Lc, moreover, because R to Lc deletions do not show this effect. T h e r-xl chromosome induces a high frequency of aneuploids (WE- BER 1973), a feature not characteristic of r-g:de. Proximally, the r-xl deletion extends beyond R toward golden. T h e region proximal to R evidently carries the gene or genes whose absence in r-xl is responsible for nondisjunction. T h e same can be inferred concerning genes responsible for the complete failure of
500 J. L. KERMICLE
Paper no. 2579 from the Laboratory of Genetics. Supported by Department of Energy contract AC02-76EVO 1300 and National Science Foundation grant PCM-8209036.
LITERATURE CITED
ASHMAN, R. B., 1960 Stippled aleurone in maize. Genetics 45: 19-34.
ASHMAN, R. B., 1965 Mutants from maize plants heterozygous R-r R-st and their association with ASHMAN, R. B., 1970 T h e compound structure of the R-st allele in maize. Genetics 64: 239- BRINK, R. A., 1973 Paramutation. Annu. Rev. Genetics 7: 129-152.
BRINK, R. A., D. F. BROWN, J. KERMICLE and W. H. WEYERS, 1960 paramutant R phenotype in maize. Genetics 45: 1297-1 3 12.
CALOS, M. P. and J. H. MILLER, 1980 Transposable elements. Cell 2 0 579-595. DOONER, H. K. and J. L. KERMICLE, 1974
DOONER, H. L. and J. L. KERMICLE, 1976 FINCHAM, J. R. S. and G. R. K. SASTRY, 1974
COLDBERG, M. L., J.-Y. SHEEN, W. J. GEHRINC and M. M. GREEN, 1983 crossing over. Genetics 51: 305-3 12.
245.
Locus dependence of the
Reconstitution of the R-r compound allele in maize. Displaced and tandem duplications in the long arm of Controlling elements in maize. Annu. Rev. Genet. Unequal crossing-over associated with asymmetrical synapsis between nomadic elements in the Drosophila melanogaster genome. Proc. Natl. Acad. Sci. USA 8 0 5017-5021.
KERMICLE, J. L., 1970 Somatic and meiotic instability of R-stippled, an aleurone spotting factor in maize. Genetics 64: 247-258.
KERMICLE, J. L. and J. D. AXTELL, 1981 Modification of chlorophyll striping by the R region.
MCWHIRTER, K. S. and R. A. BRINK, 1962 Continuous variation in level of paramutation at the STARLINGER, P., 1980 IS elements and transposons. Plasmid 3: 241-259.
STYLES, E. D. and R. A. BRINK, 1969. T h e metastable nature of paramutant R alleles in maize. IV. Parallel enhancement of R action in heterozygotes with r and in hemizygotes. Genetics
A test of distributive pairing in Zea mays utilizing doubly nonosomic plants. R-stippled maize as a Genetics 7 8 691-701.
chromosome 10 in maize. Genetics 82: 309-322. 8 15-50.
Maydica 2 6 185-197.
R locus in maize. Genetics 47: 1053-1074.
61: 801-811. WEBER, D. F., 1973
WILLIAMS, W. M., K. V. SATYANARAYANA and J. L. KERMICLE, 1984 Theor. Appl. Genet. 43: 167-173.
transposable element system. Genetics 107: 477-488.