THE EXPRESSION OF THE TABBY AND CRINKLED GENES IN DIFFERENT GENETIC BACKGROUNDS IN THE MOUSE
BERENICE KINDRED1
Division of Animal Genetics, C.S.I.R.O., P.O. Box 90, Epping, N.S.W., Australia
Received October 7, 1966
WO mutants in the mouse, Tabby and crinkled, produce identical phenotypes when homozygous. The Ta/Ta or cr/cr mouse has a long, narrow face, a very smooth looking coat with bare patches behind the ears, no hair on the tail and a markedly reduced number of secondary vibrissae (FALCONER, FRASER and KING
1952). The coat of the mouse normally consists of four types of hair: guard hairs, awls, auchenes and zigzags (DRY 1962). Closer examination of the hair of Ta/Ta or cr/cr mice reveals that the coat consists entirely of one type of hair which resembles an awl in length and shape but lacks the orderly arrangement of pig- ment granules found in normal awls (Figure 1 ) . The mutants differ in that
GUARD- AWLS A U C H E N E S Z I G Z A G S HAIRS
.
J-
NORMAL
c
V 1
NORMAL
FIGURE 1.-Typical hair from normal and homozygous mice (after FALCONER, FRASER and KING 1951).
Present address. The Institute for Cancer Research, 7701 Burliolme Are., I~hiladelphia, Pennsylvania. 1!)111
1 74 3. KINDRED
7 9 II I3 15 R 19
VIBRISSA NUMBER
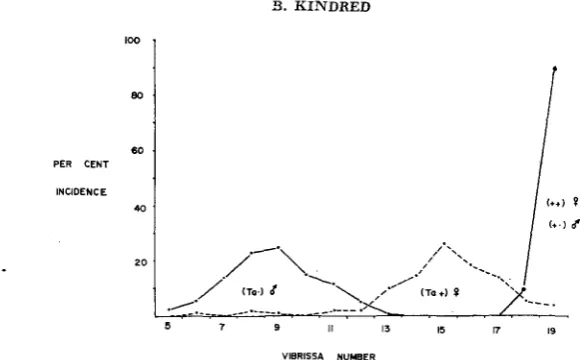
FIGURE 2.-Distribution of vibrissa numbers in unselected
+/+
(or +/Y), Ta/+ and Ta/Ymice.
crinkled is an autosomal recessive and Tabby, a sex linked semi-dominant. Ta/Y
males are phenotypically identical with Ta/Ta females as far as can be deter- mined, while Tu/+ females have striped coats and an intermediate number of secondary vibrissae.
The number of secondary vibrissae in a normal mouse is an almost invariant 19, but in the mutants there is an increase in variance accompanying the decrease in number (Figure 2 ) . These vibrissae are distributed in groups on the face and fore limbs. Selection for high and low vibrissa number in Tabby mice has pro- duced a clear response, not only in Tabbies but also in their non-Tabby sibs thus showing that there was genotypic variation underlying the phenotypic invariance of vibrissa number (DUN and FRASER 1959). After 15 generations of upward selection the modal vibrissa score of Tu/+ females was 19, the normal number, and at the same time the striping of the coat was reduced to such an extent that sometimes adult Ta/+ mice could not be distinguished from +/-I-. Thus as the mean vibrissa score approached the normal canalisation zone, the effects of the Tabby gene on both vibrissae and coat became less. At the same time, in the IOW line the stripes of Ta/+ females were very obvious and the mice often had long narrow faces and, when young, bare patches behind the ears. In other words, the dominance of the gene was changed in both directions by the selection in opposite directions.
G E N E S WITH S I M I L A R P H E N O T Y P E S 175
and Ta/Y cr/cr were not included in this comparison because the production and identification of these genotypes in any quantity is difficult and is complicated in the low line by the reduced viability and fertility of the extreme phenotype.
MATERIALS A N D M E T H O D S
The mouse stocks used were produced by repeatedly backcrossing the cr gene to the high and low vibrissa selection lines, high selection Tabby (HST) and low selection Tabby (LST) and were designated HSTcr and LSTcr. +/+ or +/Y segregants from the selection lines were used for these matings for the first five generations and then Tabby mice were used. The number of secondary vibrissae was scored at 5 days of age and checked a t 10 days using a desk lamp and dark background. Vibrissae that were less than half the normal length were not counted. The groups scored were supra-orbital, post-orbital, post-oral, inter-ramal and ulna-carpal.
Hair samples were taken from the rump region when the mice were 3 weeks old. At this age the first coat is fully grown and the second wave of hair growth has not begun. The hair types were analysed i n the manner described by FRASER (1951); at least 300 hairs from each animal were counted. About 500 hairs from each T a / + mouse were scored in an attempt to average out differences between the agouti and non-agouti stripes. As these stripes are very narrow in 3 week old mice of these stocks, errors should not be great. Hairs were counted as abnormal if there was obvious disorganisation of the pigment granules. All counts were made by the same observer to ensure that bias in classification should be consistent.
RESULTS
After five generations the mean scores of
+/+
mice from the backcross lines were very close to those of+/+
or +/Y from a comparable generation of the main selection line, indicating that the background genotype of each pair of lines was similar. The data from cr/cr males and females were pooled as there was no apparent difference. The correspondence between these and the selection lines Ta/Y males was again close (Table 1 ).
The difference of 0.9 in the loiw line was entirely due to a single group of vibrissae, the post-orals. The generation of inbreeding necessary to produce cr/cr mice from the backcross lines was carried out without selection and is probably responsible for most if not all of the differ- ence found because the post-oral score of Ta/Y males increased by 0.4 vibrissae after one generation of relaxed selection.In unselected Tabby mice the mean vibrissa score for T u / + was roughly inter- mediate between
+/+
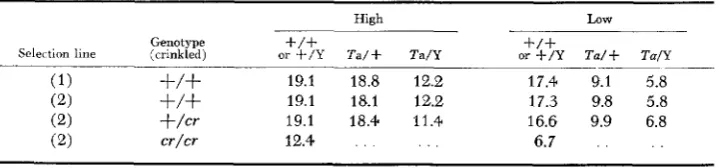
and Ta/Y (Table 2). Selection for high vibrissa scoreTABLE 1
Mean vibrissa scores of selection and backcross lines
High LOW
+/+
or +/Y T a / + Tam
Genotype
+/+
~~~
Sele< tinn line (crinkled) or + / Y Ta/+ Ta/Y
-
(1)
+/+
19.1 18.8 12.2 17.4 9.1 5.8( 2 )
+/+
19.1 18.1 12.2 17.3 9.8 5.8(2) + / c r 19.1 18.4 11.4 16.6 9.9 6.8
( 2 ) cr/cr 12.4 . . . . . 6.7 . . . .
176 B. K I N D R E D
TABLE 2
Mean vibrissa scores of unselected mice (taken from FRASER, NAY and KINDRED 1959)
Crinkled genotype f / f or + / Y Ta/ f Ta/Y
+/t
+/cr
13 9.4 6.6
13 10.9 6.1
increased the difference (DUN and FRASER 1959), i.e. high selection reduced the effect of a single Tabby substitution while low selection increased the effect. This was paralleled by changes in the coat. Most Tu/+ mice could easily be classified as HST or LST according to the degr3e of striping of the coat. In fact some HST Tu/+ showed no striping at all.
FRASER, NAY and KINDRED (1959) found no difference between the vibrissa scores of
+/+
and +/cr mice in an unselected stock and in the present study no difference was found in HSTcr. In LSTcr however the mean of +/cr mice was significantly lower than the mean of+/+
(Table 1 ).
This suggests a change in the effect of the cr gene in parallel <with the increasing expression of the Tu gene in the low line. At the Ta/+ level the scores of Tu/+ +/cr are slightly higher than those of Tu/++/+
in HSTcr but not in LSTcr. A similar effect was found by FRASER, NAY and KINDRED (Table 2) but was interpreted as the result of crossing two different stocks. At the Tu/Y level +/cr mice had higher scores than+/+
in LSTcr. It should be noted that in the results of FRASER, NAY and KINDRED the ulnacarpal group was not scored making the normal No. 6 vibrissae less.Coat characteristics: The frequency of auchenes in the coat of the mouse showed a proportionately greater response to selection on vibrissae than did the vibrissae themselves (FRASER and KINDRED 1962). The difference between
+/+
and +/cr in LSTcr also showed more clearly in the coat. Because making fibre arrays and counting hairs is extremely tedious, only ten mice were used in each group, but the difference between the means OP 4.3% auchenes for+/+
and 1.8% auchenes for +/cr was significant and most mice could be classified as+/+
or +/cr on the basis of the frequency of auchenes with only a small chance of error. Again no significant difference was found in HSTcr.Mean estimates for frequency of auchenes, zigzags and abnormal hairs in Tu/+
+/+
andTu/+
+/cr females are given in Table 3 . There are differencesTABLE 3
Numbers of auchenes, zigzags and abnormal hairs in the coat, expressed as a percentage of the total hairs counted
High LOW
Genotype Auchenes Zigzags Abnormal hairs Auchenes Zigzags Abnormal hairs
4.7 78.9 0 4.3 76.3 0
0
+/Y
+/+
+/y +/cr 4.1 77.6 0 1.8 75.2
Ta/+
+/+
3.1 50.4 15.0 0.8 35.5 15.0GENES WITH SIMILAR PHENOTYPES 177
in frequency of zigzags associated with the stripes of Ta/+ mice
(FALCONER
1953) so 300 to 500 hairs were taken in groups of 50 to 100 to ensure that both dark and agouti regions (would be included. The coats of Tal+ mice contain up to 50% of grossly abnormal hairs but only normal hairs have been included. I n auchene frequency as in vibrissa score, the difference between+/+
and +/cr is evident. When the comparison of+/+
and +/cr is made in the presence of Tu/+, results are difficult to interpret but this is very likely due to the numbers of abnormal hairs in Ta/+. The commonest of these resemble those of Ta/Ta and cr/cr but there are also zigzags with two rows of pigment granules and auchenes with a single row of granules on one side of the constriction. These demonstrate that the hairs not usually formed in homozygotes (FALCONER, FRASER and KING 195 1 )are also affected by the mutant genes.
In LSTcr, where there was a range of vibrissa scores, the percentage of ab- normal hairs was negatively correlated with the vibrissa score. The correlation coefficients for Ta/+ + / c r and Ta/+
+/+
were -0.39 and -0.32 respectively. Only the former was significant. Correlation coefficients could not be calculated for the high line since most animals have vibrissa scores of 19, but there were just as many abnormal hairs as in the low line and the same types occurred in about the same proportions. The few animals with lower vibrissa scores also tended to be the ones with the greatest number of abnormal hairs. In both high and low lines all three types of abnormal hair occurred through the whole range of vibrissa scores. It may appear contradictory that there should be a within line correlation when the numbers of abnormal hairs in the high and low lines were similar but it was only vibrissae that were under selection and it has been shown (FRASER and KINDRED 1962) that the canalisation of vibrissae and hair is different.
DISCUSSION
The reduction of vibrissa scores in low Tabby selection (LST) means that the scores of all mice 'were moved away from the normal canalisation zone a t 19. Furthermore, the strength of canalisation is reduced in LST (KINDRED 1963). Under these conditions the Ta/+ phenotype became more extreme and the +/cr
phenotype became distinguishable, i.e. crinkled was no longer completely reces- sive. Thus dominance, or the magnitude of the effect of the substitutiion of a mutant for a normal gene, depends not only on the action of the mutant but also on a developmental system which acts to reduce phenotypic variation. Dominance modification is well known and is claimed by FORD (1964) to be common in wild populations. As dominance causes more individuals to have the same phenotype, it is a means of reducing phenotypic variation. Canalisation also reduces pheno- typic variation and it is interesting that selection for low vibrissa number has reduced both canalisation and dominance. If selection for dominance modification is as strong as FORD suggests, selection for canalisation may also be much stronger than has been supposed.
178 B. K I N D R E D
forced one another. However, if T a and
Ta+
should be competing for a substrate neither may be getting enough to be effective in hair production and upsetting the balance in either direction could be an improvement.In HST Ta/+ the reduction of striping is probably due to an increase in the number of zigzags in the coat. As zigzags always have a sub-terminal yellow band they contribute considerably to a normal looking agouti coat. However, although the coat appears more normal, there are just as many abnormal hairs in LST.
Selection on vibrissa number has changed the number of vibrissae. the frequen- cies of hair types (particularly auchenes) in the coat, the striping of the coat and the dominance of the Tabby gene as it is reflected in these characters, but ab- normality in the internal structure of the hairs has not been affected. It seems that there may be two aspects to the action of the Tabby mutant, only one of which has been influenced by selection.
S U M M A R Y
The autosomal gene crinkled ( c r ) was crossed into stocks which had been selected for high and low vibrissa number in Tabby ( T a ) , a sex linked mutant. The extreme phenotypes of Tabby and crinkled are identical. The vibrissa scores of cr/cr mice are very similar to those of T a I y from the same stock and there is also a similarity in the effect on the dominance of the mutant.
LITERATURE CITED
DUN, R. B., and A. S. FRASER, 1959 DRY, F. W., 1926
FALCONER, D. S., 1952 A totally sex-linked gene in the house mouse. Nature 169: 664-665. FALCONER, D. S., A. S. FRASER, and J. W. B. KING, 1951 The genetics and development of
FORD, E. B., 1964 Ecological Genetics, Methuen, London; Wiley, New York. FRASFX, A. S., 1951 Growth of the mouse coat. J. Exptl. Zool. 117: 15-29. FRASER, A. S., and B. M. KINDRED, 1962
in the house mouse. 111. Correlated responses. Australian J. Biol. Sci. 12: 188-206. FRASER, T. S., T. NAY, and B. M. KINDRED, 1959
Australian 3. Biol. Sci. 12: 331-339. KINDRED, B., 1963
Selection for a n invariant character, vibrissa number in
the house mouse. Australian J. Biol. Sci. 12: 506-523.
The coat of the mouse. J. Genet. 16: 287-341).
- 1953 Total sex-linkage in the house mouse. Z. Ind. Abst. Vererb. 85: 210-219.
'crinkled' a new mutant in the house mouse. J. Genet. 50: 324-344..
Selection for an invariant character, vibrissa number,
Variation of vibrissa number i n the mouse.