0095-1137/08/$08.00⫹0 doi:10.1128/JCM.02405-07
Copyright © 2008, American Society for Microbiology. All Rights Reserved.
Genotypic Diversity of Coagulase-Negative Staphylococci Causing
Endocarditis: a Global Perspective
䌤
Cathy A. Petti,
1,2* Keith E. Simmon,
2Jose M. Miro,
3Bruno Hoen,
4Francesc Marco,
3Vivian H. Chu,
5Eugene Athan,
7Suzana Bukovski,
8Emilio Bouza,
9Suzanne Bradley,
10Vance G. Fowler,
5Efthymia Giannitsioti,
11David Gordon,
12Porl Reinbott,
13Tony Korman,
14Selwyn Lang,
15Cristina Garcia-de-la-Maria,
3Annibale Raglio,
16Arthur J. Morris,
17Patrick Plesiat,
4Suzanne Ryan,
18Thanh Doco-Lecompte,
19Francesca Tripodi,
20Riccardo Utili,
20Dannah Wray,
21J. Jeffrey Federspiel,
5K. Boisson,
4L. Barth Reller,
5David R. Murdoch,
22Christopher W. Woods,
5,6and the International Collaboration on
Endocarditis—Microbiology Investigators
Departments of Medicine and Pathology, University of Utah School of Medicine, Salt Lake City, Utah1; ARUP Laboratories, Salt Lake City,
Utah2; Hospital Clinic Institut d’Investigacions Biome`diques August Pi i Sunyer, University of Barcelona, Barcelona, Spain3;
University Medical Center of Besanc¸on, Besanc¸on, France4; Duke University Medical Center5and Veterans Administration Medical Center,6
Durham, North Carolina; Barwon Health, Geelong, Australia7; University Hospital for Infectious Disease, Zagreb, Croatia8;
Hospital General Universitario Gregorio Maran˜o´n, Madrid, Spain9; Veterans Administration Medical Center, Ann Arbor,
Michigan10; Attikon University General Hospital, Athens, Greece11; Flinders Medical Centre, Adelaide, Australia12;
The University of South Wales, Sydney, Australia13; Southern Health, Clayton, Australia14; Middlemore Hospital,
Auckland, New Zealand15; Ospedali Riuniti di Bergamo, Bergamo, Italy16; Auckland City Hospital,
Auckland, New Zealand17; South Eastern Sydney and Illawarra Area Health Service, Australia18;
CHU Nancy-Brabois, Nancy, France19; Second University of Naples, Naples, Italy20;
Medical University of South Carolina, Charleston, South Carolina21; and
University of Otago, Christchurch, New Zealand22
Received 16 December 2007/Returned for modification 26 February 2008/Accepted 14 March 2008
Coagulase-negative staphylococci (CNS) are important causes of infective endocarditis (IE), but their microbiological profiles are poorly described. We performed DNA target sequencing and susceptibility testing for 91 patients with definite CNS IE who were identified from the International Collaboration on Endocardi-tis—Microbiology, a large, multicenter, multinational consortium. A hierarchy of gene sequences demon-strated great genetic diversity within CNS from patients with definite endocarditis that represented diverse
geographic regions. In particular, rpoB sequence data demonstrated unique genetic signatures with the
potential to serve as an important tool for global surveillance.
Coagulase-negative staphylococci (CNS) are increasingly important causes of community- and health care-associated infective endocarditis (7, 12, 17). Although more than 40 spe-cies make up this heterogeneous group of microorganisms, identification of CNS to the species level often is not per-formed because of laboratory uncertainty about its clinical relevance or the absence of reliable identification systems (10). There is, however, growing evidence that identification of CNS to the species level may alter diagnostic and therapeutic clin-ical decision making where specific species have unique
viru-lence factors (e.g.,Staphylococcus lugdunensis) (2) or unusual
antibacterial resistance patterns (e.g., glycopeptide resistance withS.epidermidisandS.haemolyticus) (4–6). In recent years, partial 16S rRNA gene sequencing has emerged as an accurate and reliable method to identify CNS, but this molecular target is limited by having less than 1% sequence divergence among
some CNS species. Alternative gene targets such astuf
(elon-gation factor Tu) (10) andrpoB(RNA polymerasesubunit)
(9, 14) have been evaluated, but to our knowledge, no studies have applied gene sequencing of these targets from patients with definite CNS endocarditis.
Given the limited frequency of CNS endocarditis in a single institution, the International Collaboration on Endocarditis— Microbiology (ICE-Micro), a large, multicenter, multinational consortium, provided a unique opportunity to improve our understanding of the spectrum of CNS microorganisms impli-cated in prosthetic and native valve endocarditis. We per-formed gene sequencing with multiple DNA targets to identify CNS from patients with definite endocarditis to the species level. We also evaluated the potential for a hierarchy of se-quence data to provide greater specificity for species identifi-cation, serve as an epidemiologic tool to assess clonality, and predict antimicrobial resistance.
MATERIALS AND METHODS
CNS isolates from patients with definite endocarditis were submitted by ICE-Micro investigators representing a collection from 18 medical centers in 12 countries. Conventional identification and susceptibility testing were performed at a central laboratory with a commercially available panel processed on the Microscan Walkaway instrument (PC-21; Dade Behring, Deerfield, IL) by a * Corresponding author. Mailing address: University of Utah School
of Medicine, ARUP Laboratories, 500 Chipeta Way, Salt Lake City, UT 84108. Phone: (801) 583-2787. Fax: (801) 584-5207. E-mail: cathy [email protected].
䌤Published ahead of print on 26 March 2008.
1780
on May 16, 2020 by guest
http://jcm.asm.org/
standard laboratory protocol. Rifampin susceptibility testing was performed by E-test (AB Biodisk, Solna, Sweden). Interpretation of antimicrobial susceptibil-ity results was based on Clinical and Laboratory Standards Institute guidelines (8). Template DNA preparation and amplification were performed directly on frozen stocks as previously described (18). Amplification of 16S rRNA,tuf, and rpoBgenes was achieved with the following primer pairs: 16S rRNA 5F (5⬘-TT GGAGAGTTTGATCCTGGCTC-3⬘) and 1194R (5⬘-ACGTCATCCCCACCT TCCTC-3⬘),tufTseq271 (5⬘-AAYATGATIACIGGIGCIGCICARATGGA-3⬘) and Tseq1138 (5⬘-CCIACIGTICKICCRCCYTCRCG-3⬘) (13), andrpoB2491F (5⬘-AACCAATTCCGTATIGGTTT-3⬘) and 3241R (5⬘-GCIACITGITCCATA CCTGT-3⬘) (9).
PCR products were bidirectionally sequenced with original amplification prim-ers, and sequences were compared to related sequences in SmartGene IDNS-Bacteria (SmartGene Inc., Raleigh, NC) by a standard laboratory protocol (18). Nucleotide and amino acid sequence alignments and phylogenetic trees were constructed by the neighbor-joining method with Kimura’s two-parameter dis-tance correction model and 1,000 bootstrap replications in the MEGA version 3.1 software package (11). Rifampin resistance was evaluated by using the rifampin resistance-determining region of therpoBgene as described by Murphy et al. (15) for all strains of CNS.
RESULTS
Ninety-one isolates represented 91 patients with definite
endocarditis (prosthetic [n⫽ 36], native [n ⫽34], and other
[n ⫽ 21]) with median and mean ages of 68 and 63 years,
respectively. The numbers of infections classified as health care or community associated were similar. With phylogenetic
anal-ysis as the “gold standard” for final identification,S.
epidermi-dis(n⫽65) was the most common CNS identified, followed by
S.lugdunensis(n⫽8),S.hominis(n⫽6),S.capitis(n⫽5),S. haemolyticus (n ⫽ 3), and other (n ⫽ 3). Identification by conventional methods agreed with gene sequencing results for 79 (87%) of 91 isolates (Table 1). Phenotypic methods
mis-identified sixS.epidermidisisolates asS.hominissubsp.
homi-nis(three),S.capitissubsp.urealyticus(one),S.hyicus(one),
and S. warneri (one). Additionally, phenotypic methods
mis-identified one isolate each ofS.epidermidisasS.aureusandS.
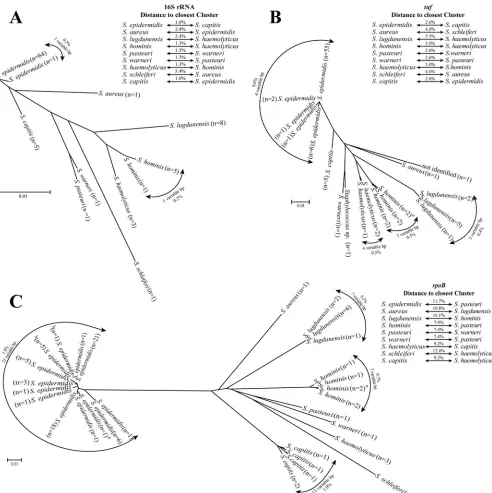
pasteuriasS.warneri. The characterization of 91 CNS isolates by three genetic targets is summarized in Fig. 1. Overall, di-versity between species in decreasing order was found with
rpoB, tuf, and 16S sequences. Neighbor-joining dendrograms
for all gene targets clearly showed distinct clusters for all of the
species, with thetufandrpoBgenes having more intraspecies
variability. No groups with five or more isolates having
identi-calrpoBsequences clustered within specific institutions or
geo-graphic regions including the distinct group ofS. epidermidis
sequences. No association was observed between the type of
valve and the distinctS.epidermidisclusters (data not shown).
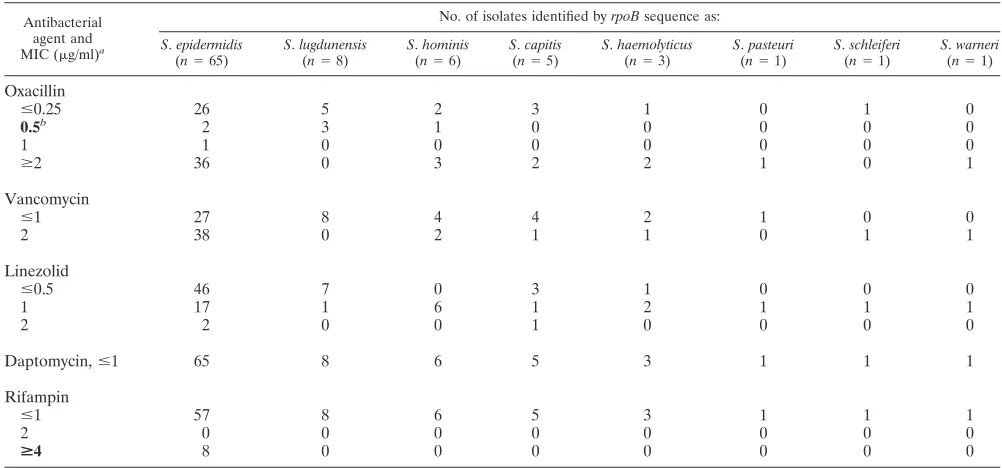
The distribution of antimicrobial susceptibilities for each
spe-cies of CNS is described in Table 2. Notably, 42% of theS.
epidermidisand 100% of theS.lugdunensisisolates tested were susceptible to oxacillin. Susceptibility patterns did not appear
to vary with geographic distribution forS.epidermidis(data not
shown). WhenrpoB amino acid sequences were analyzed for
markers of rifampin resistance in CNS isolates, we identified three distinct sequences resulting from four amino acid alter-ations among all eight rifampin-resistant isolates (Table 3). All
rifampin-resistant isolates wereS.epidermidis(MIC,⬎32g/
ml), each having two unique amino acid substitutions within
therpoBgene that are known to confer rifampin resistance on
S. aureus. Six of these eight isolates had D471E and I527M, one had H481N and I527M, and one had D471Y and H481N amino acid substitutions. No unique amino acid substitutions
were observed for non-S.epidermidisCNS species.
DISCUSSION
The specimen repository of the ICE-Micro multinational consortium provided us with a unique opportunity to examine CNS isolates associated with invasive disease. To our knowl-edge, this study is the first to fully characterize the species distribution and susceptibility patterns of CNS isolates from
patients with definite endocarditis. Sequencing of the rpoB
gene served as a robust target for identification to the species
level, suggested an absence of clonality in strains causingS.
epidermidisendocarditis, and identified high-level rifampin
re-sistance inS.epidermidisisolates.
[image:2.585.42.550.81.244.2]Historically, investigators have relied on conventional meth-ods for the identification and susceptibility testing of CNS; this may have served as a barrier to the full appreciation of the epidemiology of CNS disease. In fact, previous reviews of CNS endocarditis have discussed the spectrum of disease from the perspective of phenotypic identifications with limited attention
TABLE 1. Identification by conventional method and gene sequencing
Final identification by phylogenetic
resolution (no. of isolates)
Identity according to following test method:
Conventionala
16S rRNA tuf rpoB
S.epidermidis(59) S.epidermidis S.epidermidis S.epidermidis S.epidermidis
S.epidermidis(3) S. hominissubsp.hominis S.epidermidis S.epidermidis S.epidermidis
S.epidermidis(1) S. capitissubsp.urealyticus S.epidermidis S.epidermidis S.epidermidis
S.epidermidis(1) S. hyicus S.epidermidis S.epidermidis S.epidermidis
S.epidermidis(1) S. warneri S.epidermidis S.epidermidis S.epidermidis
S.lugdunensis(8) S.lugdunensis S.lugdunensis S.lugdunensis S.lugdunensis
S.hominis(6) S.hominis S.hominis/xylosus S.hominis S.hominis
S.haemolyticus(3) S.haemolyticus S.haemolyticus S.haemolyticus S.haemolyticus
S.capitis(5) S.capitis S.capitis/caprae/arlettae/epidermidis S.capitis S.capitis
S.schleiferi(1) S.schleiferi S.schleiferi Staphylococcussp. S.schleiferi
S.aureus(1) S. epidermidis S.aureus S.aureus S.aureus
S.pasteuri(1) S. warneri S.pasteuri/aureus Staphylococcussp. S.pasteuri
S.warneri(1) S.warneri S.warneri/pasteuri S.warneri S.warneri
a
Boldface type indicates misidentification by conventional methods.
on May 16, 2020 by guest
http://jcm.asm.org/
to specific species (17, 22). We foundS.epidermidisas the most common CNS species, an observation corroborated by previ-ous reports on non-endocarditis patients with invasive CNS
disease (1, 3, 10, 16, 19). The rank order of non-S.epidermidis
infections differs in our report from that in others. We report S.lugdunensisas the second most common pathogen, whereas
non-endocarditis studies have observedS.hominis,S.
haemo-lyticus, andS.capitisas the next most commonly encountered CNS pathogens (1, 19, 21).
The clinical implications of more accurate identification of CNS by gene sequencing are not fully known. In our study, we
[image:3.585.47.539.66.559.2]observed that therpoB gene serves as a reliable indicator of
FIG. 1. Neighbor-joining radial dendrograms of the 16S rRNA (A),tuf(B), andrpoB(C) genes of CNS isolates from patients with endocarditis. Each entry represents a unique sequence among the study isolates. The number of isolates sharing 100% identity with the representative sequence is noted. Intraspecies variability is recorded as the percent difference between isolates of the same species, as well as the number of base pair positions in the interrogated sequence with instability. To the side of each tree, the relative distance from each species to its nearest neighbor is recorded in terms of percent distance. A superscript letteraindicates two isolates identified by Microscan asS.hominissubsp.novobiosepticusthat had uniquetufandrpoBsequences which differed by at least 4 bp (0.6%) and 2 bp (0.3%) from the otherS.hominissequences, respectively. A superscript letterbindicates rifampin-resistant isolates.
on May 16, 2020 by guest
http://jcm.asm.org/
genetic diversity, which may be helpful as an epidemiological tool to distinguish multiple CNS strains. Also, fast identifica-tion of two distinct CNS strains may prove useful when inter-preting the clinical significance of blood cultures in patients with intracardiac devices. With our limited data set, we did not observe distinct regional variations among clusters of CNS isolates; however, the ability to rapidly identify isolates to the species and subspecies levels may prove valuable for monitor-ing the dissemination of unusual strains between and within institutions. Only by gene sequencing were we able to identify
and now report the first case ofS.pasteuriendocarditis.
Addi-tionally, rpoB gene sequencing affords greater specificity by
providing a unique genetic signature for CNS species that may have implications for global surveillance. While multilocus se-quence typing schemes have emerged as important tools to
assess clonal complexes forS.epidermidis, the optimal
discrim-inatory loci to serve as the gold standard have not been firmly
established (20). The use of therpoBgene shows promise as a
marker for unique clones, but its use as a sole epidemiological target warrants further study by multilocus sequence typing.
Accurate isolate identification to the species level is also important for examining susceptibility patterns and alerting clinicians to those species with known increased virulence. Although we did not observe reduced susceptibilities to
glyco-peptides inS. epidermidis and S. haemolyticus, these species
reportedly exhibit heterogeneous susceptibility to glycopep-tides (4–6) and their identification may heighten clinician awareness of therapeutic failures of vancomycin therapy.
Sim-ilarly, identification ofS.lugdunensismay alter the diagnostic
approach because this species is considered to be more sus-ceptible to beta-lactam agents, more virulent, and associated with a higher mortality compared with other CNS species (2). Finally, we report the first use of the rifampin
resistance-determining region of therpoBgene to predict rifampin
resis-tance in isolates identified asS.epidermidis. Given the need for
[image:4.585.41.542.80.314.2]rifampin in patients with CNS prosthetic valve endocarditis, a reliable method to identify locations known to confer rifampin resistance by single-step mutations may be clinically important. In conclusion, the ICE-Micro consortium enabled us to eval-uate a large number of CNS isolates from patients with definite endocarditis from diverse geographic regions with a hierarchy of gene sequence data. From this extensive global repository, we provide novel and valuable information about the genetic diversity of CNS species that cause endocarditis, suggesting
TABLE 2. Distribution of antimicrobial susceptibilities of CNS isolates
Antibacterial agent and MIC (g/ml)a
No. of isolates identified byrpoBsequence as:
S.epidermidis (n⫽65)
S.lugdunensis (n⫽8)
S.hominis (n⫽6)
S.capitis (n⫽5)
S.haemolyticus (n⫽3)
S.pasteuri (n⫽1)
S.schleiferi (n⫽1)
S.warneri (n⫽1)
Oxacillin
ⱕ0.25 26 5 2 3 1 0 1 0
0.5b 2 3 1 0 0 0 0 0
1 1 0 0 0 0 0 0 0
ⱖ2 36 0 3 2 2 1 0 1
Vancomycin
ⱕ1 27 8 4 4 2 1 0 0
2 38 0 2 1 1 0 1 1
Linezolid
ⱕ0.5 46 7 0 3 1 0 0 0
1 17 1 6 1 2 1 1 1
2 2 0 0 1 0 0 0 0
Daptomycin,ⱕ1 65 8 6 5 3 1 1 1
Rifampin
ⱕ1 57 8 6 5 3 1 1 1
2 0 0 0 0 0 0 0 0
>4 8 0 0 0 0 0 0 0
aBreakpoints for susceptibility are those defined by the Clinical and Laboratory Standards Institute (8). An MIC in boldface type represents the breakpoint for
resistance.
bThe breakpoint forS.lugdunensisoxacillin resistance isⱖ4g/ml.
TABLE 3. Variability of therpoBamino acid sequence for
S.epidermidisisolatesa
No. of isolates
Nucleotide change(s) inrpoB
Amino acid alteration
Rifampin resistance (MIC关g/ml兴)
55 None None Susceptible (⬍1)
1 GCT3GAT A5343D Susceptible (⬍1) 1 CGT3CAT R5033H Susceptible (⬍1) 1 GAT3TAC D4713Yb Resistant (⬎32)
CAC3AAC H4813Nb
1 CAC3AAC H4813Nb Resistant (⬎32) ATA3ATG I5273Mb
6 GAT3GAA D4713Eb Resistant (⬎32) ATA3ATG I5273Mb
a
Amino acid sequences of therpoBgene were compared to GenBank se-quence NP_763861 (ATCC 12228).
b
Regions known to be involved with resistance to rifampin (15).
on May 16, 2020 by guest
http://jcm.asm.org/
[image:4.585.45.283.543.698.2]that strains causing CNS endocarditis have unique genetic sig-natures which are found across vast geographic distances. The clinical significance of more accurate identification to the
spe-cies level remains to be defined, butrpoB sequence analysis
may serve as a useful tool for surveillance and may improve our understanding of the host-pathogen relationships in native and prosthetic valve endocarditis.
ACKNOWLEDGMENTS
There are no conflicts to declare.
ICE-Micro receives support from Cubist Pharmaceuticals (C.W.W.).
REFERENCES
1.Aldea-Mansilla, C., D. G. de Viedma, E. Cercenado, P. Martin-Rabadan, M. Martin, and E. Bouza. 2006. Comparison of phenotypic with genotypic procedures for confirmation of coagulase-negativeStaphylococcus catheter-related bloodstream infections. J. Clin. Microbiol.44:3529–3532. 2.Anguera, I., A. Del Rio, J. M. Miro, X. Matinez-Lacasa, F. Marco, J. R.
Guma, G. Quaglio, X. Claramonte, A. Moreno, C. A. Mestres, E. Mauri, M. Azqueta, N. Benito, C. Garcia-de la Maria, M. Almela, M. J. Jimenez-Exposito, O. Sued, E. De Lazzari, J. M. Gatell, and the Hospital Clinic Endocarditis Study Group.2005.Staphylococcus lugdunensisinfective endo-carditis: description of 10 cases and analysis of native valve, prosthetic valve, and pacemaker lead endocarditis clinical profiles. Heart91:e10.
3.Becker, K., D. Harmsen, A. Mellmann, C. Meier, P. Schumann, G. Peters, and C. Von Eiff.2004. Development and evaluation of a quality-controlled ribosomal sequence database for 16S ribosomal DNA-based identification of Staphylococcusspecies. J. Clin. Microbiol.42:4988–4995.
4.Bertin, M., A. Muller, X. Bertrand, C. Cornette, M. Thouverez, and D. Talon.2004. Relationship between glycopeptide use and decreased suscep-tibility to teicoplanin in isolates of coagulase-negative staphylococci. Eur. J. Clin. Microbiol. Infect. Dis.23:375–379.
5.Biavasco, F., C. Vignaroli, R. Lazzarini, and P. E. Varaldo.2000. Glycopep-tide susceptibility profiles ofStaphylococcus haemolyticusbloodstream iso-lates. Antimicrob. Agents Chemother.44:3122–3126.
6.Bourgeois, I., M. Pestel-Caron, J. F. Lemeland, J. L. Pons, and F. Caron.
2007. Tolerance to the glycopeptide vancomycin and teicoplanin in coagu-lase-negative staphylococci. Antimicrob. Agents Chemother.51:740–743. 7.Chu, V. H., C. H. Cabell, E. Abrutyn, G. R. Corey, B. Hoen, J. M. Miro, L.
Olaison, M. E. Stryjewski, P. Pappas, K. J. Anstrom, S. Eykyn, G. Habibi, N. Benito, V. G. Fowler, and the International Collaboration on Endocarditis Merged Database Study Group.2004. Native valve endocarditis due to coagulase-negative staphylococci: report of 99 episodes from the Interna-tional Collaboration on Endocarditis Merged Database. Clin. Infect. Dis.
39:1527–1530.
8.Clinical and Laboratory Standards Institute.2007. Performance standards for antimicrobial susceptibility testing; seventeenth informational
supple-ment. CLSI document M100-S17. Clinical and Laboratory Standards Insti-tute, Wayne, PA.
9.Drancourt, M., and D. Raoult.2002.rpoBgene sequence-based identifica-tion ofStaphylococcusspecies. J. Clin. Microbiol.40:1333–1338.
10.Heikens, E., A. Fleer, A. Paauw, A. Florijn, and A. C. Fluit.2005. Compar-ison of genotypic and phenotypic methods for species-level identification of clinical isolates of coagulase-negative staphylococci. J. Clin. Microbiol.43:
2286–2290.
11.Kumar, S., K. Tamura, and N. Nei.2004. MEGA 3: Integrated software for molecular evolutionary genetics analysis and sequence alignment. Brief. Bioinform.5:150–163.
12.Lalani, T., Z. A. Kanafani, V. H. Chu, L. Moore, G. R. Corey, P. Pappas, C. W. Woods, C. H. Cabell, B. Hoen, C. Selton-Suty, T. Doco-Lecompte, C. Chirouze, D. Raoult, J. M. Miro, C. A. Mestres, L. Olaison, and S. Eykyn.
2006. Prosthetic valve endocarditis due to coagulase-negative staphylococci: findings from the International Collaboration on Endocarditis Merged Da-tabase. Eur. J. Clin. Microbiol. Infect. Dis.25:365–368.
13.Martineau, F., F. J. Picard, D. Ke, S. Paradis, P. H. Roy, M. Ouellette, and M. G. Bergeron.2001. Development of a PCR assay for identification of staphylococci at genus and species levels. J. Clin. Microbiol.39:2541–2547. 14.Mellmann, A., K. Becker, C. von Eiff, U. Keckevoet, P. Schumann, and D. Harmsen.2006. Sequencing and staphylococci identification. Emerg. Infect. Dis.12:333–336.
15.Murphy, C. K., S. Mullin, M. S. Osburne, J. van Duzer, J. Siedlecki, X. Yu, K. Kerstein, M. Cynamon, and D. M. Rothstein.2006. In vitro activity of novel rifamycins against rifamycin-resistantStaphylococcus aureus. Antimi-crob. Agents Chemother.50:827–834.
16.Persson, L., H. Strid, U. Tidefelt, and B. Soderquist.2006. Phenotypic and genotypic characterization of coagulase-negative staphylococci isolated in blood cultures from patients with haematological malignancies. Eur. J. Clin. Microbiol. Infect. Dis.25:299–309.
17.Rupp, M. E., and G. L. Archer.1994. Coagulase-negative staphylococci: pathogens associated with medical progress. Clin. Infect. Dis.19:231–243. 18.Simmon, K. E., A. C. Croft, and C. A. Petti.2006. Application of SmartGene
IDNS software to partial 16S rRNA gene sequences for a diverse group of bacteria in a clinical laboratory. J. Clin. Microbiol.44:4400–4406. 19.Spanu, T., M. Sanguinetti, D. Ciccaglione, T. D’Inzeo, L. Romano, F. Leone,
and G. Fadda.2003. Use of the VITEK 2 system for rapid identification of clinical isolates of staphylococci from bloodstream infections. J. Clin. Micro-biol.41:4259–4263.
20.Thomas, J. C., M. R. Vargas, M. Miragaia, S. J. Peacock, G. L. Archer, and M. C. Enright.2007. Improved multilocus sequence typing scheme for Staph-ylococcus epidermidis. J. Clin. Microbiol.45:616–619.
21.Weinstein, M. P., S. Mirrett, L. van Pelt, M. McKinnon, B. L. Zimmer, W. Kloos, and L. B. Reller.1998. Clinical importance of identifying coagulase-negative staphylococci isolated from blood cultures: evaluation of MicroScan rapid and dried overnight gram-positive panels versus a conventional refer-ence method. J. Clin. Microbiol.36:2089–2092.
22.Whitener, C., G. M. Caputo, M. R. Weitekamp, and A. W. Karchmer.1993. Endocarditis due to coagulase-negative staphylococci: microbiologic, epide-miologic, and clinical considerations. Infect. Dis. Clin. N. Am.7:81–96.