water, sodium, potassium, chloride, inulin and collagen and of serum for water, sodium, po tassium, chloride and inulin have been previ ously described.3
Adrenal Insufficiency
Male Sprague-Dawley rats weighing 200 to 300 grams were used in all experiments. The animals were bilaterally adrenalectomized through a single dorsal incision. They were maintained for 5 to 7 days by providing 0.9% sodium chloride solution and water to drink. They continued to receive their standard ra tion.° Following this period of stabilization, the salt solution was removed and the animals had only water to drink.
With this regimen evidence of adrenal in sufficiency developed in 3 to 5 days. At that time a bilateral nephrectomy was done and the animals were each given a single intravenous injection of inulin. The usual amount of inulin given was 1 mi/100 gm of body weight of a 10% solution of inulin in 0.9% sodium chlonide.t The exact weight of the material given was de termined by weighing the syringe before and after the injection. The volume given was ob tained from the weight of a 1-mi aiiquot.
Six hours after the injection of inulin, the animals were killed by exsanguination under ether anesthesia. The abdomen of each animal was shaved just prior to this procedure and skin was removed from this area. The major tendons of both hind feet were rapidly dis sected and removed; any attached muscle fi bers were scraped off. Nonadrenalectomized control animals were treated similarly.
The tissues obtained from these animals
0Friskies—Carnation Milk Co.
t U. S. Standard Products, Mount Prospect, Illi
nois. T HE CONCEPT that the connective tissue
of the body, unlike parenchymal cells, participates in direct diffusion equilibrium with the extracellular fluid and blood plasma, and hence can be most properly re garded as the supporting tissue for the ex tracellularfluid,was firstadvanced by Manery and Hastings1 and was reviewed by Manery in 1954.2 Further data in support of this concept were published by Nichols
et al.3
The experiments described herein were undertaken to determine how changes in duced in electrolyte composition of the ex tracellular fluid would be reflected in elec trolyte composition of the connective tissue. For the purpose of this paper, the con nective tissue is defined as the complex of the water (and its contained electrolyte, pro tein and mucopolysaccharide) and the cells associated with the noncalcified supporting collagen and elastin fibers of the body. This heterogenous organ contains both “¿dense― and “¿loose―connective tissue as exemplified by tendon and skin.
PLAN OF EXPERIMENTSAND METHODS
Three types of experiments were performed to determine the pattern of the ionic equilib rium between connective tissue and extracellu lar fluid: altered extracellular fluid electrolyte concentrations were produced by adrenal in sufficiency, reduced extracellular volume was produced by water deprivation and chloride depletion by intraperitoneal dialysis with a chloride-free solution. In vitro studies of tis sues immersed in varying dilutions of an artifi cial extracellular fluid were also carried out.
The methods used for analyses of tissue for
Supported in part by the National Institute of Arthritis and Metabolic Diseases of the National In stitutes of Health, Public Health Service. (Grants G-3754 and A-2384).
ADDRESS:(W.B.W.) 2065 Adelbert Road, Cleveland 6, Ohio.
PEDIATRICS, December 1960
915
THE EFFECTOF ALTERATIONSIN EXTRACELLULARFLUID
ON THE COMPOSITION OF CONNECTIVETISSUE
William B. Weil, Jr., M.D., and William M. Wallace, M.D.
Department of Pediatrics, Western Reserve University School of Medicine, and
were used for insulin assays. Tissues for water and electrolyte determination were obtained from other similarly treated control and adren alectomized animals 6 hours after nephrectomy; these animals were given an equivalent volume of 0.9% sodium chloride solution without inulin.
Water Deprivation
Albino rabbits weighing 3 to 4 kg were used. Shaved skin from the anterior abdominal wall, tendon from one hind leg and blood by cardiac puncture were obtained under ether anesthesia. Two weeks later the animals were offered their dry ration ad libitum, but were deprived of water until they had lost 18 to 20% of their body weight. They were then killed and blood, skin and tendon removed for analy sis.
Diffusion Experiments
IN VITRO: Skin and tendon from albino rab
bits were used for these experiments. One piece of skin and tendon, as well as serum, were used for control determinations. Additional pieces of abdominal skin and hind-leg tendon were placed in flasks containing varying dilu tions of an electrolyte solution simulating the ionic pattern of extracellular fluid. Jsotonicity was maintained with mannitol. The flasks were kept at 37°Cand the contents were aerated by bubbling a mixture of 95% oxygen and 5% car bon dioxide in them throughout the equilibra tion period. At the end of this time the tissues were removed, blotted, weighed and analyzed. The actual composition of each bath was deter mined by direct analysis. Tissues were kept in the solutions for 6 hours; previous work had demonstrated that this period was adequate for complete equilibration.4
IN VIvo: Six adult rats were lightly anes thetized with pentobarbitol sodium, and in traperitoneal dialysis was performed for 2 hours in two animals and for 6 hours in the other four. One of the animals that was dia lyzed for 6 hours was killed 18 hours after the dialysis. All other animals were killed upon completion of the dialyzing procedure.
Samples of blood and skin were obtained at the beginning of the experiment and blood, skin and tendon were obtained at the end. These were analyzed for fat, water, sodium, potassium and chloride.
The dialyzing solution contained: sodium, 150 meq/l; potassium, 4.0 meq/l; calcium, 4.5
meq/l; and magnesium, 1.5 meq/l. The sul fate concentration was 160 meq/l. Glucose was added to make a 2.5% solution. The fluid was introduced through a 23 gauge needle in the left upper quadrant of the abdomen and the outflow was through a 15 gauge needle in the right lower quadrant. The solution was given as a continuous infusion at the rate of 40 ml! hr. The effluent was collected and analyzed to determine the absolute amount of chloride re moved from the body.
RESULTS
Adrenal Insufficiency
The results of the analyses of serum, skin and tendon for the normal, the nephrecto mized control, and the nephrectomized adrenalectomized experimental animals are shown in Table I. The values are given both in reference to water and to fat-free dry solids. Similar data are given for the animals studied by Cole5 and by White and Rolf.6 The animals with adrenal insufficiency demonstrated the expected decrease in sodium and increase in potassium with little decrease in chloride of the serum. The water content of the tissues was essentially unchanged and the electrolyte changes in the tissues were in the same direction as in the serum. In the animals in the present study, the decrease in concentrations of sodium and chloride in skin water exceeded the decrease in sodium and chloride in ul trafiltrate of the plasma, producing smaller sodium and chloride “¿spaces―in the adrenal ectomized animals. Although the decrease in chloride in the skin was less than for sodium, with no decrease occurring in the chloride concentration in the serum, the change in chloride in the skin produced a decrease of chloride space comparable to that of the sodium space. Thus, there was no change in the relation of sodium and chloride volumes of distribution. In tendon, loss of electrolyte paralleled the loss in serum so that there was no change in so dium or chloride volume of distribution.
Present SeriesCole'White et al.6NormalNephrect. Control,Neph, ccl. plus Adrenalect.Control Adrenalect.Control AAdrenaleel. C 917 (ECF Cone.)
156.1± 1.6 126.8± 7.9± 0.4 11.4± 100.1± 2.2 95.1±
11.2 143.0±2.5 135.0±3.3 3.1 3.8±0.9 6.3±0.8 9.4 104 ±3.6 99.0±2.7 943 ± 4.7
149.1± 2.7 5.4± 0.6 100.5± 2.0 10 246 ±11.7 28.1± 1.9 14.2± 2.2 22.2± 1.5 114.7± 8.8 57.4± 7.6 90.2± 4.7 8 152 ± 5.5
17.8± 1.6 5.7± 0.8 13.1± 2.8 117.3± 13.6 38.9± 5.9 86.4± 8.1 600 ±30 608 ±39 210 ±16 763 783 804 770 342 226
945 ± 5.1
137.5± 2.7 10.8± 2.2 101.2± 4.5 7 246 ±18.1 21.6± 1.5 17.8± 1.8 18.3± 2.3 87.5± 4.1 72.4± 7.7 74.4± 10.9 6
149 ± 10.2
16.4± 2.8 6.5± 0.9 13.1± 1.8 110.3±10.2 43.3± 3.9 87.8± 13.9 418 ±50 525 ±17 208 ±35 630 790 653 771 430 238 10 137 19.1 6.6 19.7 139.4 48.4 148.7 10 136 16.1 8.1 18.1 117.5 59.5 132.5 15 219 21.2 14.5 19.7 97.0 66.4 89.7 678 782 453 7 232 20.0 16.9 19.7 86.5 73.1 84.9 641 777 578 Serum 1120 (nil/i) Na (meq/i) K (meq/i) Ci (meq/i) Skin Number
1120 (mi/i® gm FFDS)
Na (meq/100 gm FFDS) K (meq/100 gm FFDS) Ci (meq/100 gm FFDS) Na (meq/i 11,0) K (meq/l 1120)
Ci (meq/i 11,0)
Tendon
Number
11,0 (ml/100 gm FF05) Na (meq/100 gm FFI)S) K (meq/i00 gm FFI)S) Ci (meq/100 gm FF1)S) Na (meq/I00 gm FF05) K (meq/100 gm FF05) Ci (meq/100 gm FF05)
inulin space
Skin (mi/I 1120) Tendon (mi/i 11,0) Whole body (mi/kg body
weight)
Sodium Space
Skin (mi/i 1W@) Tendon (mi/i 1120)
Chloride Space
Skin (mi/i 11,0) Tendon (mi/i 11,0)
Potassium Space
Skin (mi/i 11,0) Tendon (mi/i 11,0)
SCN Space (mi/kg tissue)
278.5± 3.9 258.8± 4.6
242 ±22.6 201 ±17.1
892 926
1,486 1,393
296 431
947 ± 3.7
147.0± 5.4 4.5± 0.9 103.6± 4.9 16 278 ±34 31.7± 4.2 15.5± 3.7 27.3± 3.7 113.4± 7.1 53.8± 10.7 97.3± 7.0 5
139 ± 4.5
16.8± 1.1 5.4± 1.1 15.3± 0.8 121.3± 8.8 39.4± 8.6 110.1± 8.2 770 824 846 958 340 230
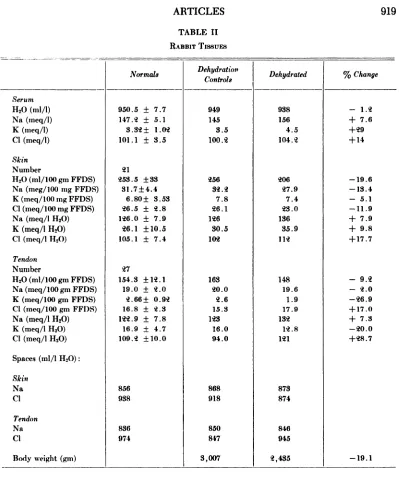
shown in Table II. The values for the pre dehydration tissues and sera are comparable to those for the control group. The period of dehydration required for the animals to lose 18 to 20% of theirbody weight variedfrom
98 to 167 hours. Following this, the water in skin had also decreased about 20%, but ten don water had diminished by only 9%.
Except for the chloride content of tendon, the ratio of the electrolytes to the fat-free dry solids decreased in all animals. The water decreased more than the electrolytes, however, so that there was an increase in extracellular concentration of electrolyte ARTICLES
TABLE I
RAT TIssuEs
water to that in the extracellular fluid. As has been shown before,3 the inulin space is smaller than either the chloride or sodium spaces in tendon and skin. The inulin space of skin and tendon decreased to a greater extent than the sodium and chloride spaces, but the whole-body inulin space was un altered.
Dehydration
per liter of tissue water. This hypertonicity was also noted in the serum. Although there were small changes in the volumes of dis tribution of sodium and chloride, none of these were consistent in all animals.
In-vitro Diffusion Studies
The individual values for the concentra tions of sodium and chloride in rabbit skin and tendon compared to the concentration of the ions in the solutions in which the tis sues were immersed, are shown graphically in Figure 2. The straight line equations which have been statistically derived from these data are given in Table III. In these equations the amount of nondiffusible ion
is represented by the constant “¿a―(if the
concentration of the ion in the media were zero, the amount in the tissue would be equal to “¿a―).The slope of tile line “¿b―mul tiplied by 1000 equals the volume of distri
hution of tile diffusible iOn, assuming tile concentration in this volume varies as the
concentration in the medium. The values
for this volume are also shown in Table III.
The 95% confidence limits for the values are given in parentheses. The correlation coefficients between tile concentrations in water of the tissues and in the media were 0.99 for both ions in both tissues. Although there are differences in the slopes of the four lines, they are not significantly differ ent at the 95% confidence level. The differ ences do suggest, however, that the chloride
space may be larger than tile sodium space
in skin and in tendon, even after the non diffusible portion is discounted. There is significantly more chloride than sodium “¿bound―in tendon, and more chloride bound in tendon than there is in skin.
The results of the in-vivo peritoncal di alysis of six rats, using a synthetic serum
SPACES: MLJL. TISSUE
WATER
TENDON
Nx + ADX
SKIN
Nx 1000
900
800
TOO
600
500
400
300
200
100
NA@CC IN NA@ CC IN
FIG. 1. Sodium, chloride, potassium and inulin spaces in skin and tendon from
rats. The nephrectomized (Nx) animals served as controls for the animals that were
adrenalectomized and then nephrectomized (Nx + Adx). The “¿spaces―occupied by
sodium, chloride and inulin were determined by dividing the concentration of the
substance in tissue water by the concentration in extracellular fluid. The (lenonli
nator used for calculating potassium space was tile sum of concentrations of
sodium atiti I)otIlssitIm ill extracelitilar water; the numerator was total potassium
less that calculated to be Pr'5t'lIt in tile extracehiular fluid. Tile potaSsillnl space
was plotted downward from the 1000—mi line to indicate the discre@)ancy between
intracellular water as measured by potiissitim and extraceilular water as measured
by inuhin. This difference, represented by tile clear area in tile third column of
each set, would theoretically represent the connective tissue protein phase (see
Figure 3).
Nx NX+ADX
ARTICLES 919
TABLE II
RABBIT Tissum
Dehydration
Controls
NormaLs Dehydrated % Change
Sernm 1120 (mi/i) Na (meq/1) K (meq/1) Ci (meq/i) Skin Number
H,0 (ml/100 gm FFDS) Na (meg/100 mg FFDS)
K (meq/100 mg FFDS)
Cl (:neq/100 mg FFDS)
Na (meq/i 1120)
K (meq/1 1120) Ci (meq/i 1120)
Tendon Number
1130(mi/100 gm FFDS)
Na(meq/100gmFFDS)
K (meq/100 gm FFDS)
Cl (meq/100 gm FFDS)
Na (meq/l 1120) K (meq/l 1120) Cl (meq/1 1120)
Spaces (mi/I 1120):
Skin Na Ci Tendon Na
ci
Body weight (gm)
950.5 ± 147.2 ± 3.32 ± 101.1 ± 7.7 5.1 1.02 3.5 —¿ 1.2 + 7.6 +29 +14 —¿19.6 —¿13.4 —¿ 5.1 —¿11.9 + 7.9 + 9.8 +17.7 —¿ 9.2 —¿ 2.0 —¿26.9 +17.0 + 7.3 —¿20.0 +28.7 —¿19.1 21 253.5 ±33 31.7± 4.4 6.80± 3.53
26.5 ± 2.8
126.0 ± 7.9 26.1 ±10.5
105.1 ± 7.4
27
154.3 ±12.1
19.0 ± 2.0
2.66± 0.92 16.8 ± 2.3 122.9 ± 7.8 16.9 ± 4.7 109.2 ±10.0 856 938 836 974 949 145 3.5 100.2 256 32.2 7.8 26.1 126 30.5 102 163 20.0 2.6 15.3 123 16.0 94.0 868 918 850 847 3,007 938 156 4.5 104.2 206 27.9 7.4 23.0 136 35.9 112 148 19.6 1.9 17.9 132 12.8 121 873 874 846 945 2,435
ultrafiltrate with the chloride ion replaced by sulfate, are shown in Table IV. Up to 50% of the total body chloride was removed in this manner, and this is reflected in the levels of chloride in the serum. In general, the decrease in chloride in serum and skin correlate well with the amount of chloride lost by dialysis. The tendon, however, did not share in the deficit to the same extent. The number of animals studied is too small to be more specific than to state that the equilibrium in skin is comparable to that
seen in the in-vitro experiments with rab bit tissue.
DISCUSSION
Tissue Concentration = a +b Medium
(rneq/l tiss. 1120) (meq) (meq/lConcentration 1120)Skin
sodium Skin chloride
Tendon sodium
Tendon chloride(19)=
8.5 (±7.5) +0.877 (±.176)
(19) = 4.3 (±2.1) +0.922 (± .058)
(30) = 6.9 (±2.2) +0.925 (± .050)
(30) = 10.0 (± 1 .3) +0.936 (± .035)Bath
sodium
Bath chloride
Bath sodium Bath ehiori(IeVolume
of distribution of diffusil)ie ion: Skin sodium =877 (mI/I tiss. 1120)
Skill chloride = 922 (mI/I tiss. 1120)
Tendon sodium = 925 (mI/i tiss. 1120) Tendon chloride = 936 (mi/i tiss. 1120)
TISSUE TISSUE
CONG CONC.
(MEQ/L.) (MEO/L.)
., $
150 SKIN —¿Na@ @‘¿B TENDON —¿NA@
NA,: 6.9(±22) + O.925(t.05O) NA,,' •¿
NA3. 8.5(±7@5)+O.877(t.I76) NAB /
i
•¿@• /3
/
,
IOO@ ‘¿ 00 #4
, ,
S
I, S/S
/ 5'
A /
S., /,
50 50 ,‘
,,.@# / @5S ,@/
5/
, S_I' #45
/ / / /
C, / , /
___________ 5' fr, 1,93'l
0@ •¿1 0 ,•
0 50 / 0 50
/
, ,
/ dVP$
‘¿1 ,
“¿I
, ,
/ ,
50 / 50
5/ “¿SI
SKIN—¿Ci ‘¿ TENDON—¿Ci
, /
, Cs, :4.3(±2.I)+ O.922(±.058)Cs, ‘¿ Cs, =IOO(±I3)+O.936(±.035)Cs,
0@ , 0 I
0 50 00 50 0 50 100 50
BATH CONG. (sEn./L.) BATH CONC. (MEQJL)
FIG. 2. In-vitro equilibration of sodium and chloride in skin and tendon from rab
bits. The values for each ion in each tissue are expressed as meq/l of tissue water and compared with the measured concentrations in the solutions in which they
were immersed. The dashed line represents the equations for these values, which
are the least squares estimate of the relationship. The numbers in parentheses are the 95% confidence limits for the values.
TABLE III
RABBITS: IN-VITRO I)IALYSIS4
Serum Cl
(meq/l)Skin
Cl
(meq/l 1120)Tendon Cl (meq/l 1120)Skin
Cl (meq/100
gm FFDS)Tendon
Cl (meq/100
qm FFDS)Cl
removed (meq/100 gm
body weight)Dialysis
Time (hr)98.4
45.9 56.3 67.6 79.0
80.5
83.198.0
46.8 50.9 67.7
70.0 67.2
81.5Control
Values (mean for six animals)
110.2 ( 28.0 15.3
Experimental Values
68.2 13.2 10.2
98.2 13.4 14.0
83.6 17.7 12.3
96.4 ‘¿20.2 13.6
15.3
97.3 ‘¿22.6 15.30
1.72 1.56 1.19
1.27 0.72
0.490
6 6 6
6* 2
‘¿2
ARTICLES 921
TABLE IV
RATS: IN-VIVO DIALYSIS
* Kiiied 18 hours later.
were 150 ml/100 gm FFDS, there would be 70 ml of water associated with collagen in each kilogram of body weight; this would represent about one third of the ex tracellular fluid. Over one half of the con nective tissue water would be in skin and most of the remainder associated with a more dense form of collagen in bone, mus cle sheaths and tendon. In the adult rat the skin, as an organ, is second only to muscle in weight; and bone, in which the protein is almost entirely collagen,9 is the next largest organ in the fat-freeanimal.1°
Two pertinent questions regarding the role of the connective tissue in body fluid homeostasis are:
1) Are the electrolytes and water of the connective tissue in equilibrium with the rest of the extracellular fluid and therefore available to participate pro portionally with volume and ionic changes in the total extracellular fluid? 2) Is the connectivetissuea storehouse,
i.e.,can electrolytesor water be pref
erentially removed from, or deposited in, this tissue?
One reason for examining the connective tissue in this regard is that in total-body analysis, when bone is excluded, the chlo ride “¿space―is larger than the sodium “¿space―6and connective tissue is the major organ in which this relationship has been shown to occur.'1 The other factor is that
at the pH of body fluid'2 some of the con nective tissue protein has been shown to be on the alkaline side of its isoelectric point, thus acting as an anion; but mucopoly saccharides and proteins may function as polyelectrolytes and to a degree bind both anions and cations.― Closely allied with the ionic attraction is the water of hydration of these proteins; this is important in relation to the question of the extent to which the connective tissue can contribute its water and salt to the body as a whole.
922 CONNECTIVE TISSUE periments and in the literature. The volume
of fluid occupied by substances such as mu un, sucrose and thiocyanate in skin or ten don has always been found to be much less than either sodium or chloride spaces3'13 and the distribution of these materials in short time periods would define the rapidly equilibrating phase.
In the adrenalectomized rats the decrease in inulin space in the connective tissue is greater than the decrease in either the so dium or chloride spaces. It is also greater than the increase in the potassium space. (The potassium space is calculated from the premise that the intracellular concen tration of potassium is the same as the sum of the extracellular concentrations of so dium plus potassium.) This could be ex plained by an increase in the connective tis sue solids phase, which contains no appreci able potassium or inulin. This concept is di agrammed in Figure 3b. There are two
ways in which this apparent increase in connective tissue protein phase could occur. The penetration of inulin into the connec tive tissue may be decreased by swelling of the fibers or change in the ground sub stance, or the amount of electrolyte bound to these proteins may be increased by the metabolic changes occurring in adrenal in sufficiency. Desoxycorticosterone actetate
has been shown to increase the inulin space in the total body of monkeys without change in salt or water content.15 It would seem reasonable that the reverse situation could occur in adrenal insufficiency. The data of White et al.6 is consistent with this interpretation. Day'6―7 has demonstrated some of the effects of changes in pH and in salt concentration on the state of hydration, spatial distribution and electrolyte attrac tion of the connective tissue proteins. Both of his studies, as well as our own, are com patible with the concept of connective tis
ADRENAL CONTROL
1000
500-EXTRACELLULAR PHASE
RAPIDLY EQUILIBRATING
OR
‘¿INULIN SPACE'
CONNECTIVE TISSUE
PROTEIN PHASE
BOUND SPACE
INTRACELLULAR PHASE
POTASSIUM SPACE
K@4-PROTEIN
B
0-A
Ftc. 3. Hypothetical structure for the volumes of distribution of
sodium and chloride in skin water for control animals (A) and
adrenaietcomized animals (B). In this representation of the
sodium and chloride of connective tissue, these ions are shown
in terms of their “¿volumesof distribution,― thus there will be
more sodium than chloride actually present in the tissue if the
“¿spaces―are equal, because of the greater concentration of
sodium than of chloride in extracellular fluid.
PROTEIN
PROTEIN
ARTICLES 923 tonic expansion of tissue fluids assumed to be similar to that found by Leaf would give values for skin sodium space of 805 ml/l of tissue water, for chloride—876 ml/l of tis sue water; for tendon sodium space—714 ml/l and for chloride—756 ml/l. The small amount of electrolyte assumed to be non diffusible would not be appreciably affected by this dilution effect. The in-vitro diffusion studies indicate that connective tissue may act as a storage area for sodium and chlo ride to a very limited extent; the prepond erance of these ions and the water in this tissue are in equilibrium with the extracel lular fluid of the body.
The modifications of extracellular fluid composition, volume and concentration all demonstrate by direct analysis what Swan and Pitts2t found by infusion of hydro chloric acid, what Cotlove'4 suggested from long-term inulin infusion, and what Wolf and McDowell2' demonstrated by determin ing the osmotic volume of distribution of chloride, namely, the connective tissue is an insignificant storage area for chloride or sodium and that the fluid and extracellular electrolytes of the connective tissue will vary proportionatelywith alterationsof the total extracellular fluid.
SUMMARY
Variations in the volume and composition of the extracellular fluid were produced by adrenal insufficiency, dehydration and intra peritoneal perfusion with a chloride-free solution. Direct analysis of blood, skin and tendon were performed to determine the ex tent to which loose and dense types of con nective tissue varied with the changes in the plasma ultrafiltrate. Less than 10% of the sodium and chloride in tissue is associated with connective tissue solids in such a way as not to be readily diffusible.
On the basis of these studies, a structure for connective tissue has been postulated with three divisions: A rapidly-equilibrat ing phase, a noncellular connective tissue solids phase and an intracellular phase. The noncellular solids, acting as polyelectro lytes, could bind ions to a variable extent sue already described. This interpretation
suggests that the connective tissue can act as a storage site for electrolytes to a limited extent by variable binding of cations or anions when there is alteration in acid-base balance or of concentration in body fluids. A second means by which connective tissue could act as a reservior results from the rela tively slow or incomplete way in which dense connective tissue (e.g., tendon) ap pears to reach equilibrium with the re mainder of the extracellular fluid. With these limited qualifications, the connective tissue appears to contribute its share to the body fluid losses in adrenal insufficiency. The data of White and Rolf6 on analyses of whole tissue would confirm this.
The tissue studies in the dehydrated rab bits show that the percentage loss of water from the skin is similar to the loss of body weight. Water loss from the tendon is only half this amount, however. Although total body analyses were not done, it would ap pear that considering the connective tissue as a single organ, the water loss would be proportional to the loss in the entire animal, since tendon and other dense connective tissue is a small part of the total. The simi larity of the relation between sodium and chloride spaces in the skin of the normal and dehydrated animals indicates that this part of the connective tissue does not func tion as a storage place for either ion inde pendently. The increase of chloride space in tendon is slight, but is suggestive as of some importance in explaining part of the chlo ride retention known to occur in hypertonic dehydration.'@ The dehydration in these rabbits was of a hypertonic nature, since sodium concentration in the serum went from 145 to 156 meq/l.
CONNECTIVE TISSUE depending on acid-base and fluid tonicity
changes.
The reduced rate of equilibration of dense connective tissue with extracellular fluid produces a phenomenon which would appear as “¿storage―in certain situations.
Thus by binding and by delayed equilib ria, connective tissue has a small capacity to act as a storage area for extracellular electrolyte, but in general, the fluid and salt of connective tissue is in direct diffu sion equilibrium with plasma ultrafiltrate.
REFERENCES
1. Manery, J. F., and Hastings, A. B.: The distribution of electrolytes in rat tissues. Am. J. Physiol., 119:372, 1937. 2. Manery, J. F.: Water and electrolyte
metabolism. Physiol. Rev., 34:334, 1954. 3. Nichols, C., Jr., Nichols, N., Weil, W. B., and Wallace, W. M.: The direct meas urement of the extracellular phase of tissues. J. Cliii. Invest., 32:1299, 1953. 4. Weil, W. B.: Unpublished data, 1952. 5. Cole, D. F.: The effects of adrenalectomy
on the electrolyte composition of the skin of the rat. Acta endocrinol., 11:9, 1952. 6. White, H. L., and Rolf, D.: Whole tis
sue electrolyte analyses in normal and adrenalectomized rats. Am. J. Physiol., 180:287, 1955.
7. Wallace, W. M., Weil, W. B., and Taylor, A.: The effect of variable protein and mineral intake upon the body composi tion of the growing animal. Ciba Foun dation Colloquia on Ageing, 4:116, 1958.
8. Idem: Unpublished data, 1959.
9. Rogers, H. J., Weidmann, S. M., and Parkinson, A.: Studies on skeletal tis sue. 2. The collagen content of bones from rabbits, oxen and humans. Bio chem. J., 50:537, 1951-52.
10. Caster, W. 0., Poncelet, J., Simon, A. B., and Armstrong, W. D.: Tissue weights of the rat. I. Normal values determined by dissection and chemical methods.
Proc. Soc. Exper. Biol. & Med., 91:122, 1956.
11. Manery, J. F., and Hasting, A. B.: The distribution of electrolytes in mammal ian tissues. J. Biol. Chem., 127:657, 1939.
12. Theis, E. R., and Jacoby, T. F.: The acid base-binding capacity of collagen.
J. Biol.Chem.,146:163,1942.
13. Ludwig, A. W., Chen, D. K., and Soffer.L. J.: The relationship between connec tive tissue mucopolysaccharide content and tissue electrolyte composition. J. Clin. Endocrinol., 12:965, 1952. 14. Cotlove, E.: Mechanism and extent of dis
tribution of inulin and sucrose in chlor ide space of tissues. Am. J. Physiol.,
176:396, 1954.
15. Sweet, A. Y., Levitt, M. F., and Hodes, H. L.: The effect of desoxycorticoster one acetate on water and electrolyte distribution. J. Clin. Invest., 37:65, 1958.
16. Day, T. D.: The mode of reaction of inter stitial connective tissue with water. J. Physiol., 109:380, 1949.
17. Idem: The permeability of interstitial con nective tissue and the nature of the interfibrillary substance. J. Physiol., 117:1, 1952.
18. Weil, W. B., and Wallace, W. M.: Hyper tonic dehydration in infancy. PEDI ATRICS, 17:171, 1956.
19. Heymann, W.: Zur Pathogenese des Sauglingssklerems. I. Quellungsverhalt nisse. Ztschr. Kinderh., 48:238, 1929. 20. Leaf, A.: On the regulation of intracellu lar fluid volume. Scientific Souvenir, produced on the occasion of eighth an nual conference of the Indian Associa tion of Pathologists and the committee meeting of the Indian Council of Medi cal Research, 1957, pp. 78-84. 21. Swan, R. C., and Pitts, R. F.: Neutraliza
tion of infused acid by nephrectomized dogs. J. Clin. Invest., 34:205, 1955. 22. Wolf, A. V., and McDowell, M. E.: Ap
1960;26;915
Pediatrics
William B. Weil, Jr. and William M. Wallace
COMPOSITION OF CONNECTIVE TISSUE
THE EFFECT OF ALTERATIONS IN EXTRACELLULAR FLUID ON THE
Services
Updated Information &
http://pediatrics.aappublications.org/content/26/6/915
including high resolution figures, can be found at:
Permissions & Licensing
http://www.aappublications.org/site/misc/Permissions.xhtml
entirety can be found online at:
Information about reproducing this article in parts (figures, tables) or in its
Reprints
http://www.aappublications.org/site/misc/reprints.xhtml
1960;26;915
Pediatrics
William B. Weil, Jr. and William M. Wallace
COMPOSITION OF CONNECTIVE TISSUE
THE EFFECT OF ALTERATIONS IN EXTRACELLULAR FLUID ON THE
http://pediatrics.aappublications.org/content/26/6/915
the World Wide Web at:
The online version of this article, along with updated information and services, is located on
American Academy of Pediatrics. All rights reserved. Print ISSN: 1073-0397.