The role of noise in brain dynamics processes
150
0
0
Full text
Figure


+7
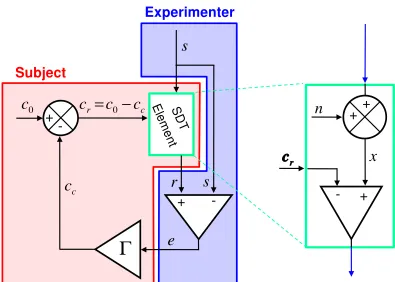

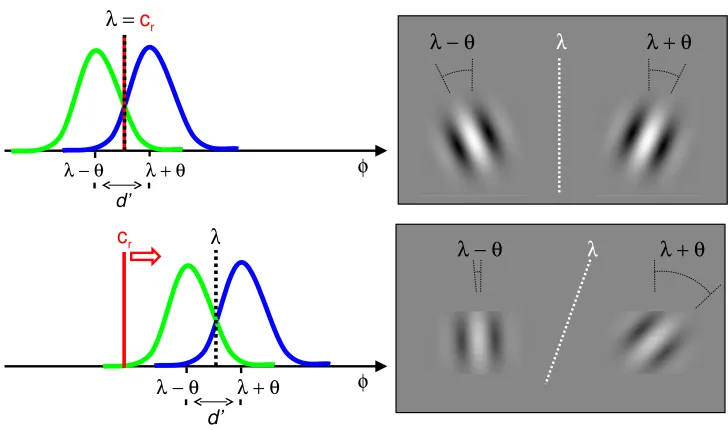

Related documents