RESEARCH ARTICLE
Notch signaling patterns neurogenic ectoderm and regulates the
asymmetric division of neural progenitors in sea urchin embryos
Dan O. Mellott, Jordan Thisdelle and Robert D. Burke*ABSTRACT
We have examined regulation of neurogenesis by Delta/Notch signaling in sea urchin embryos. At gastrulation, neural progenitors enter S phase coincident with expression of Sp-SoxC. We used a BAC containingGFPknocked into theSp-SoxClocus to label neural progenitors. Live imaging and immunolocalizations indicate that Sp-SoxC-expressing cells divide to produce pairs of adjacent cells expressing GFP. Over an interval of about 6 h, one cell fragments, undergoes apoptosis and expresses high levels of activated Caspase3. A Notch reporter indicates that Notch signaling is activated in cells adjacent to cells expressing Sp-SoxC. Inhibition of γ-secretase, injection of Sp-Delta morpholinos or CRISPR/Cas9-induced mutation of Sp-Delta results in supernumerary neural progenitors and neurons. Interfering with Notch signaling increases neural progenitor recruitment and pairs of neural progenitors. Thus, Notch signaling restricts the number of neural progenitors recruited and regulates the fate of progeny of the asymmetric division. We propose a model in which localized signaling converts ectodermal and ciliary band cells to neural progenitors that divide asymmetrically to produce a neural precursor and an apoptotic cell.
KEY WORDS: Neural progenitor, Asymmetric mitosis, Apoptosis, SoxC, Delta, Notch reporter, Neural evolution,Strongylocentrotus purpuratus
INTRODUCTION
Recent analyses of genomes has supplemented our understanding of metazoan neurobiology and led to clearer hypotheses of how complex nervous systems evolved. Sensory cells and neurons are likely to have originated early in metazoan evolution, but nervous systems appear to have arisen later and may have been derived independently in several lineages (Erwin, 2015; Liebeskind et al., 2016; Moroz et al., 2014). Comparative studies of metazoan neurogenesis suggest that the common ancestor of bilaterian animals employed a number of fundamental processes in elaborating a nervous system, and these form the basis for many of the similarities that we see in neural development (Hartenstein and Stollewerk, 2015). As cellular organization diversified and animals became more varied, neurogenesis appears to have drawn upon conserved signaling systems and molecular pathways in a variety of ways. Consequently, the cellular arrangements of neurogenesis and the resulting neural architectures of metazoans are strikingly diverse and it is a challenge to distinguish conserved
features from those that are convergent (Liebeskind et al., 2016). Nowhere is this more evident than in the deuterostomes – an assemblage that includes the chordates (cephalochordates, urochordates and vertebrates) and the Ambulacraria (echinoderms and hemichordates) (Peterson and Eernisse, 2016). Deuterostomes are allied by remarkably close molecular kinship, yet their body plans and neural organization could hardly be more different. Echinoderms and hemichordates are basal deuterostomes with features of neurogenesis that contrast sharply with those of the more elaborate chordates. Thus, detailed models of neurogenesis in ambulacrarians seem key to understanding the origins of the complex nervous systems of vertebrates.
The nervous system that arises in sea urchin embryos becomes the larval nervous system; it has a bilateral plane of symmetry and is organized to control the ciliary band, gut and muscles (Burke et al., 2006). The larval central nervous system is the apical organ, which is derived from a neuroepithelium and is ganglionic in organization (Byrne et al., 2007; Yaguchi et al., 2006). Components of the gene regulatory networks that control neurogenesis in the apical organ have been identified, and many of these genes are orthologs of those expressed in the vertebrate forebrain (Wei et al., 2009, 2016; Yaguchi et al., 2012; Garner et al., 2016). Peripheral sensory and motor neurons, embedded in the ciliary band or associated with the gut, arise in ventral ectoderm, ciliary band, adjacent dorsal ectoderm, and the endoderm (Garner et al., 2016; Wei et al., 2012, 2016). Thus, the embryonic formation of the larval nervous system has the potential to be a revealing model of deuterostome neurogenesis (Burke et al., 2014).
Neurons are postmitotic, and neural fate is manifest in neural precursors arising from the final cell division. Dedicated neural progenitors, which have stem-like properties, are found in many metazoans. These cells divide asymmetrically to produce a daughter cell that retains the progenitor state and another that either differentiates as a neuron or undergoes a limited number of symmetric divisions before differentiating (Knoblich, 2008). In other situations, regions of ectoderm have neurogenic potential and neural fate is specified independently of proliferation (Hartenstein and Stollewerk, 2015). The occurrence and distribution of neurogenic ectoderm and dedicated neural progenitors are key features of embryonic neurogenesis, yet neither is known for most bilaterians (Hartenstein and Stollewerk, 2015).
Notch signaling is a common feature of the patterning of metazoan nervous systems. Notch has diverse roles, and in many organisms it functions reiteratively to regulate neuronal abundance, patterning and diversity. Notch signaling is thought to be primarily responsible for producing theʻsalt and pepper’pattern of neural progenitors through classical lateral inhibition (Lewis, 1998). Within an epithelium, all cells express Notch and low levels of Delta. Notch activation suppresses neurogenesis and the expression of Delta, which produces a pattern of isolated neurons in a field on non-neural cells (Collier et al., 1996). A second distinct function of
Received 9 March 2017; Accepted 21 August 2017
Biochemistry and Microbiology, University of Victoria, Victoria, BC, Canada V8W 2Y2.
*Author for correspondence (rburke@uvic.ca)
R.D.B., 0000-0001-5527-4410
DEVEL
O
Notch signaling is the regulation of asymmetric divisions of progenitors (Yoon and Gaiano, 2005; Louvi and Artavanis-Tsakona, 2006; Truman et al., 2010). In the best-studied situations, a polarized neural progenitor distributes cell fate determinants to daughter cells differentially. Notch signaling is suppressed in one cell and it differentiates as a neuron, whereas the other cell either retains a progenitor state or adopts an alternative fate (Pierfelice et al., 2011). A third role for Notch signaling is emerging in which it is implicated in producing diverse neuronal or glial subtypes (Gaiano et al., 2000; Gaiano and Fishell, 2002; Corbin et al., 2008). There is no clear sense of how these diverse Notch functions are dispersed in metazoans, nor which functions are primary and which are derived.
Although many of the key features of neural development in sea urchin embryos have now been revealed, we know little about the relationship between cell division and neural fate or the details of Notch signaling. Using pulse-chase labeling of cells with a thymidine analog, Garner et al. (2016) determined that neural progenitors of the first neurons to differentiate are in S phase of their last mitotic cycle at the time of hatching. S phase is coincident with the expression of a key neurogenic transcription factor, Sp-SoxC, which is thought to identify neural progenitors (Garner et al., 2016). In addition, there are indications that Notch signaling is crucial to neurogenesis (Wei et al., 2011; Yaguchi et al., 2011, 2012; Lapraz et al., 2009; Rottinger et al., 2006; Saudemont et al., 2010). However, we lack understanding of what aspects of neurogenesis are controlled by Notch signaling. Here, we show that after neural progenitors divide, one cell becomes a neuron and the second cell undergoes apoptosis. We also demonstrate that Notch signaling regulates recruitment of neural progenitors and determines which of the progeny of the final mitosis will become a neuron. Our data suggest a model of embryonic neurogenesis in sea urchin embryos that includes bilaterian features and is likely to represent the components of neurogenesis of the common ancestor of all deuterostomes.
RESULTS
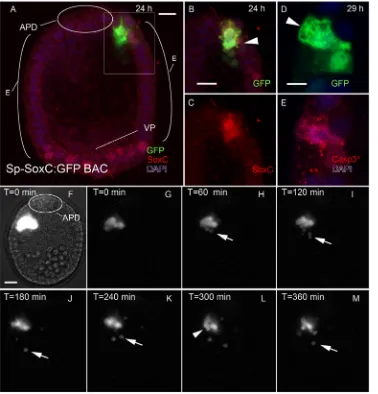
After mitosis, one cell expressing SoxC becomes apoptotic To identify Sp-SoxC-expressing cells in living embryos, we employed the Sp-SoxC:GFP bacterial artificial chromosome (BAC), which carries a 35.4 kb genomic insert in which a GFP open reading frame terminated by a stop codon effectively replaces the coding sequence for SoxC (Lee et al., 2001). The BAC contains Sp-SoxC, with 23 kb 5′and 11.2 kb 3′of the 1.1 kb gene. The adjacent genes (SPU_00331,Sp-Gal3st3) are 30 kb 5′and 66 kb 3′ to the gene, and neither is expressed embryonically. Thus, embryonic regulation ofSoxCappears to reside within this BAC. Embryos injected with Sp-SoxC:GFP express GFP as blastulae (24 h) and throughout development to pluteus. Because of mosaic incorporation, individual embryos express GFP from the transgene in only a few cells, but always in a manner consistent with the tissue-specific expression previously described for endogenous SoxC (Fig. 1, Fig. S1) (Garner et al., 2016). In mesenchyme blastulae (24 h), GFP expression is in subsets of cells in the presumptive ectoderm or mesoderm that normally express SoxC. In early gastrulae, GFP is expressed in ectoderm or the animal pole domain (APD), often in pairs of adjacent cells (Fig. 1A). In early plutei (72 h), some of the cells containing GFP have the form of neurons (Fig. S2). When Sp-SoxC:GFP-injected embryos are prepared for immunofluorescence with anti-GFP and anti-SoxC, cells expressing GFP also express Sp-SoxC, which confirms the fidelity of the Sp-SoxC:GFP BAC (Fig. 1B,C). Commonly in a pair
of GFP-expressing cells, one cell has normal cuboidal epithelial features, whereas the second has an irregular shape with numerous processes and a large intracellular vacuole (Fig. 1B,D). In embryos older than 48 h, fragments of cells containing GFP are apparent within the blastocoel (Fig. S1G-J). To control for the cellular fragments being an artifact, embryos were injected with Sp-Brn1/2/ 4:GFP, a BAC-containing regulatory sequence of Sp-Brn1/2/4, a gene expressed subsequent to the final mitosis by neural precursors. These embryos do not have GFP-positive cell fragments in the blastocoel, nor do embryos injected with mRNA that express GFP in all tissues (Fig. S3). We concluded that the Sp-SoxC:GFP BAC is a reliable means of identifying Sp-SoxC-expressing cells in living embryos.
To establish the fate of Sp-SoxC-expressing neural progenitors, embryos that were injected with Sp-SoxC:GFP were prepared to make a continuous record of cells expressing GFP. Records were made of 41 embryos with a cell or a pair of cells expressing GFP between 19 and 29 h. In 31 of these records, the embryo moved out of the field of view or began to spin. In two recordings, a single GFP-expressing cell divided between 20 and 28 h (Movies 1, 2). In four records, one of a pair of GFP-expressing cells fragmented (Movies 3-5). In all instances in which a pair of GFP cells was identified, the two cells remained in contact throughout the period of recording (Movies 3-5). In one recording made of a GFP-expressing cell that divided at 21 h (Fig. 1F-M, Movie 3), one of the GFP-expressing cells extends processes repeatedly and appears very active. Parts of the cell containing GFP are released and move into the blastocoel, where they are in continuous motion. The cell continues to extend processes and fragments throughout the 6 h interval recorded. Ultimately, the nucleus of the active cell fragments and the remnants of the cell moves into the blastocoel. The second cell of the pair has typical epithelial form throughout the movie. Fragmentation of Sp-SoxC-expressing cells was observed in cells from the ventral ectoderm, ciliary band and APD. For any given pair of cells expressing GFP from the Sp-SoxC:GFP BAC, the cell with an irregular outline is distinctive in having abundant granules in the cytoplasm that are immunoreactive to an antibody against the cleaved, activated form of Caspase3 (Fig. 1D,E) (Wei et al., 2011). We conclude from these experiments that neural progenitors expressing Sp-SoxC undergo a mitotic division and that one of the progeny of this division undergoes apoptosis.
Cells with active Notch signaling are adjacent to cells expressing SoxC
Notch signaling has been documented in vegetal lineages of embryos during late cleavage and gastrulation, where it mediates specification of mesoderm and endomesoderm segregation (Sherwood and McClay, 1999; Sweet et al., 2002; Croce and McClay, 2010). Delta has been reported to be expressed in the APD and in cells scattered throughout the ventral ectoderm and ciliary band, consistent with expression in neurons or neural progenitors (Lapraz et al., 2009; Rottinger et al., 2006; Saudemont et al., 2010). To determine if Notch signaling occurs in the ectoderm and is potentially involved with neural specification, we prepared a Notch reporter (Hansson et al., 2006). Regulatory modules that mediate Notch signaling in urchins have been identified in the 5′ cis -regulatory DNA of theGCMgene (Ransick and Davidson, 2006). The E and P modules contain five Su(H)-binding motifs, and the Su (H) effects are mediated primarily through the three E-module sites. The 3×E-P:GFP reporter construct that we prepared has three tandem duplications of the E-P modules adjacent to an eGFP open reading frame (Fig. 2A). To test the effectiveness of the construct,
DEVEL
O
we co-injected it with RNA encoding either the Notch intracellular domain (NICD) or a dominant-negative form of Su(H) (Ransick and Davidson, 2006). Embryos injected with 3×E-P:GFP plus NICD had robust expression of GFP in mesoderm, endoderm and presumptive ectoderm, whereas embryos co-injected with dominant-negative Su(H) had no expression of GFP (Fig. 2B,C). When 3×E-P:GFP was injected alone, expression of GFP was most prominent in mesoderm and ectoderm (Fig. 2D,E). In hatched blastulae (24 h), single or paired cells contain GFP. In gastrulae (48 h), pairs or small clusters of cells contain GFP. When embryos were examined at 24 h, the predominant tissue of expression is endomesoderm (Fig. 2F). However, over 60% of the embryos had some GFP expression in presumptive ectoderm and 6% expressed GFP in the APD. In 48 h embryos injected with 3×E-P:GFP, the APD is the most common site of expression, with ventral ectoderm and ciliary band being the next most common (Fig. 2G). There was relatively little expression in other tissues.
To localize expression of the Notch reporter relative to SoxC-expressing cells in the ectoderm, we labeled 3×E-P:GFP-injected embryos with antibodies against SoxC and GFP (Fig. 2H-J). All of the ectodermal cells expressing GFP had an adjacent cell that expressed SoxC (26 GFP-expressing cells in 12 embryos). Most of the cells expressing GFP also expressed SoxC (69.2%, 18/26 cells), often at low levels. We conclude from these experiments that during
gastrulation ectodermal cells with activated Notch signaling either express Sp-SoxC or are adjacent to cells expressing Sp-SoxC.
Inhibiting Notch signaling results in supernumerary neural progenitors and neurons
DAPT, which interferes with Notch signaling, has been shown to increase the number of neurons in the endoderm and APD (Wei et al., 2011; Yaguchi et al., 2011, 2012). In addition, Yaguchi et al. (2012) report that a morpholino that suppresses Delta expression produces supernumerary serotonergic neurons. As these studies examined only endodermal neurons or a subset of the APD neurons, we asked if suppression of Notch signaling would enhance neurogenesis throughout the entire larval nervous system. We employed DAPT, a morpholino against Delta (Materna and Davidson, 2012), and CRISPR/Cas9 genome editing of theDelta gene (Mali et al., 2013; Lin and Su, 2016).
[image:3.612.49.427.58.452.2]Untreated control embryos have a scattering of SoxC-expressing cells in the APD, in the ventral ectoderm, and adjacent to the ciliary band (Fig. 3A). Injection of the Sp-Delta morpholino produced embryos that at 48 h have large numbers of Sp-SoxC-expressing neural progenitors in the APD and in bilateral clusters in the position of the postoral cells (Fig. 3B). Similarly, treatment of embryos with DAPT from cleavage stages produced embryos that had supernumerary Sp-SoxC-expressing neural progenitors in the
Fig. 1.S. purpuratusembryos injected with the Sp-SoxC:GFP BAC express GFP in neural progenitors and reveal that after mitosis one SoxC-expressing cell becomes apoptotic. (A) Projection of a confocal stack of images for a 24 h embryo injected with Sp-SoxC:GFP. APD, animal pole domain; E, ectoderm; VP, vegetal pole. (B,C) An optical section showing the pair of GFP-expressing cells boxed in A. One of the cells has GFP in the nucleus, whereas the second cell (arrowhead) is irregular in outline and has GFP throughout the cytoplasm. The cells co-express SoxC. (D,E) A pair of ectodermal cells expressing GFP in a 29 h embryo injected with Sp-SoxC:GFP. One cell (arrowhead) is irregular in outline and contains granular material that binds an antibody to cleaved (activated) Caspase3 (Casp3*). (F-M) A continuous time-lapse record (Movie 3) was made of a pair of cells that had completed mitosis as the sequence begins (t=0). (F) An initial frame with brightness and contrast adjusted to reveal the location of the cells in the ectoderm. Within 60 min, fragments (arrows) containing GFP appear near the most active of the cells. The number of fragments increases over the next 4 h. After 5 h, the nucleus fragments (arrowhead). Scale bars: 10 µm.
DEVEL
O
ciliary band and ventral ectoderm (Fig. 3C). The CRISPR/Cas9 injections with two gRNAs directed against Sp-Deltaresulted in embryos that had very large numbers of Sp-SoxC-expressing cells throughout the ciliary band and APD (Fig. 3D). Within the APD, the Sp-SoxC-expressing cells are restricted to the periphery and form a ring (Fig. 3D, inset). A ring of Sp-SoxC-expressing cells also formed around the anus. In early plutei (72 h), there are normally neurons, identified with anti-Synaptotagmin B (SynB), throughout the ciliary band that contribute axons to a tightly bundled tract at the base of the ciliary band (Fig. 3E). Embryos that develop from eggs injected with the Sp-Delta morpholino develop to prism, although they frequently exogastrulate and are inflated in comparison with control embryos. In 72 h Sp-Delta morpholino-injected embryos,
there are a few clusters of differentiated neurons in the ciliary band and apical organ, but the axonal tracts are indistinct and frequently disorganized (Fig. 3F). Plutei that had been treated with DAPT from hatching had large numbers of neurons in the ciliary bands and in the apical organ (Fig. 3G). Similarly, there are substantial numbers of cells in the ciliary band and the apical organ in the CRISPR/Cas9 plutei (Fig. 3H). Detailed examination of the postoral transverse ciliary band in these embryos shows that almost all of the ciliary band cells, identified with Hnf6, also express SynB (Fig. 3I-K). In the apical organ, a similar situation exists in which there are exceptionally large numbers of neurons that extend into the ventral and dorsal ectoderm adjacent to the thickened ectoderm. These results indicate that suppressing Sp-Delta produces supernumerary neural progenitors throughout the nervous system. Although the morpholino produces an early phenotype similar to the other treatments, there is no corresponding increase in the number of neurons in plutei, suggesting that the morpholino functions best to block early expression of Sp-Delta. However, DAPT treatments or CRISPR/Cas9-induced mutations of Sp-Delta produce increased numbers of neural progenitors and neurons throughout the nervous system. We conclude from these experiments that Notch signaling is crucial to restricting the number of neurons that differentiate throughout the larval nervous system.
Early inhibition of Notch enhances recruitment of Sp-SoxC-expressing progenitors, and late inhibition of Notch increases pairs of Sp-SoxC-expressing cells
[image:4.612.51.300.58.576.2]To determine the stage in neurogenesis at which Notch signaling produces additional neural progenitors, we examined the initial stages at which Sp-SoxC is normally expressed. Previously, we reported that, prior to hatching, all cells of the APD begin to express Sp-SoxC at the same time as they express the APD marker Sp-NK2.1, but by hatching the number of cells expressing Sp-SoxC is reduced (Takacs et al., 2004; Garner et al., 2016) (Fig. 4A-D,M). In embryos treated with DAPT beginning in early cleavage stages or injected withCas9
Fig. 2. A Notch reporter construct demonstrates Notch signaling by ectodermal cells expressing Sp-SoxC.(A) The Notch reporter construct (3×E-P:GFP) based on the regulatory modules in the 5′cis-regulatory DNA of theGCMgene (Ransick and Davidson, 2006). The E and P modules contain five Su(H)-binding motifs, and the reporter has three tandem duplications of the E-P modules adjacent to an eGFP open reading frame. (B) To confirm responsiveness of 3×E-P:GFP to transcriptional co-regulation by the Notch intracellular domain (NICD), we co-injected mRNA encoding LvNICD (Sherwood and McClay, 1999). These preparations produced robust expression of GFP in all regions of the embryo. (C) Similarly, we used a dominant-negative Su(H) construct [DN Su(H)] (Ransick and Davidson, 2006) to demonstrate competitive inhibition of GFP expression. None of the embryos co-injected with DN Su(H) exhibited GFP expression. (D) 24 h embryo injected with 3×E-P:GFP at fertilization. In these embryos, most of the expression of GFP is in the endomesoderm, but ectodermal cells also express GFP. (E) A 48 h gastrula expresses GFP in a patch of ciliary band cells. As with Sp-SoxC:GFP injections, GFP-containing cell fragments in the blastocoel are common in gastrulae of embryos that were injected with 3×E-P:GFP. (F) Quantification of the proportion of 24 h embryos with GFP expression in the APD, ectoderm, and endomesoderm (n=47 cells). (G) Quantification of the proportion of 48 h embryos with GFP expression in the APD, ventral ectoderm, ciliary band, dorsal ectoderm, mesoderm, or endoderm (n=39 cells). (H-J) Confocal image showing expression of 3×E-P:GFP relative to Sp-SoxC-expressing cells in the ectoderm. Embryos that had been injected with 3×E-P: GFP were fixed and probed with antibodies to Sp-SoxC and GFP. All of the ectodermal cells expressing GFP had an adjacent cell that expressed Sp-SoxC (26 GFP-expressing cells). These data indicate that cells with active Notch signaling express Sp-SoxC or are adjacent to cells expressing Sp-SoxC. Scale bars: 10 µm.
DEVEL
O
mRNA and twoDeltagRNAs, all of the cells of the APD express Sp-SoxC initially, but as late as 28 h there is no reduction in their number (Fig. 4E,F,M). If embryos are injected with mRNA encoding NICD, there is no expression of Sp-SoxC (Fig. 4G,H,M). To determine the effects of altered Notch signaling on the recruitment of neural progenitors to the peripheral nervous system, we examined the appearance of the postoral neural progenitors. At about 28 h, two pairs of cells appear in the ventral ectoderm adjacent to where the postoral arms will form (Garner et al., 2016) (Fig. 4I,N). However, when embryos are treated with DAPT or injected withCas9mRNA and twoDeltagRNAs, the number of cells expressing Sp-SoxC in the location of the postoral cells increases to 10-14 (Fig. 4J,K,N). We conclude from these experiments that Notch signaling has a role in regulating the number of cells recruited as neural progenitors in the APD and as peripheral neurons.
To determine whether Notch signaling has a role in the asymmetric division of neural progenitors, and because our data
[image:5.612.120.494.59.337.2]indicate that Notch signaling may occur between pairs of cells expressing Sp-SoxC, we treated embryos after the initial recruitment phase with DAPT and examined the number and distribution of cells expressing Sp-SoxC. In these embryos, there is a marked increase in the number of cell pairs expressing Sp-SoxC (Fig. 4L,O). This suggests that the fate of the cells from the final division is determined in part by Notch signaling. To examine this further, we injected the Sp-SoxC:GFP BAC and either treated embryos with DAPT to suppress Notch signaling or co-injected NICD mRNA to enhance Notch signaling. In control embryos injected with Sp-SoxC:GFP and treated with DMSO, there are ectodermal cells expressing GFP and cell fragments containing GFP (Fig. 5A,F). In embryos injected with Sp-SoxC:GFP and treated with DAPT, almost all of the GFP expression is found in pairs of ectodermal cells (Fig. 5B-D,F). In embryos co-injected with NICD mRNA and Sp-SoxC:GFP, GFP expression can be found in cells within the ectoderm, but as well there is an increase in fragments within the
Fig. 3. Inhibiting Notch signaling produces supernumerary neural progenitors and neurons.(A-D) Disrupting Sp-Delta expression with a morpholino antisense oligonucleotide (MASO), inhibiting Notch receptor cleavage with DAPT, or making a CRISPR/Cas9-mediated deletion in theSp-Deltagene increases the abundance of cells expressing Sp-SoxC in gastrula stage embryos. (A) Ventral view of a control embryo with scattered Sp-SoxC expression in ventral ectoderm. (B) Embryos injected with Sp-Delta MASO (Materna and Davidson, 2012) are normal in overall appearance, but, unlike the controls, most of the cells of the APD (oval) express Sp-SoxC and there are clusters in the position of postoral (PO) cells. (C) Embryos treated with DAPT (10 µM) have more cells expressing Sp-SoxC in the APD (oval) and in clusters lateral to the presumptive mouth (m). (D) Embryos injected withSp-DeltagRNA and mRNA encoding Cas9 have the largest number of cells expressing SoxC. There is a circle of Sp-SoxC-expressing cells at the periphery of the APD (inset). Sp-SoxC is also expressed by cells in the ciliary band and adjacent ectoderm and in a ring of cells surrounding the anus (a). CRISPR/Cas9-induced mutations reveal the entire neurogenic ectoderm of the gastrula stage embryo. (E-H) The abundance of neurons in early plutei is increased in embryos in which Notch signaling has been disrupted by DAPT treatment or CRISPR/Cas9-induced mutation ofSp-Delta. (E) Control embryos have 30-40 neurons, mostly in the ciliary bands, and tracts of axons at the base of the ciliated cells. There are several neurons associated with the esophagus. (F) Embryos injected with the Delta MASO are somewhat inflated in appearance and have fewer neurons and less distinct axonal tracts than control embryos. (G) Treatment with DAPT increases the number of cells expressing Sp-Synaptotagmin B (Sp-SynB). Most of the cells have cytoplasmic Sp-SynB in some regions of the ciliary band, and the apical organ (AO) is enlarged. There are frequently supernumerary axonal tracts. (H) Embryos in which CRISPR/Cas9 was used to target mutation ofSp-Deltahave an excess of neurons. The neurons are within the ciliary band, although the apical organ is enlarged and there are neurons underlying the thickened epithelium. Where there are normally one to two postoral neurons, there are 10-12 in clusters. (I) Detail of dorsal view of apical organ of a 72 h control embryo injected with gRNA alone. There are few neurons (SynB+) relative to ciliated cells (Hnf6+) and all are within the thickened epithelium. (J,K) Embryos in which CRISPR/Cas9 was used to target mutation ofSp-Delta
probed with anti-SynB and anti-Hnf6, a marker for ciliary band cells. Almost all of the cells expressing Hnf6 also express SynB. Axonal tracts are robust and less compact. (K) The apical organ in these embryos is enlarged, and most of the cells co-express Hnf6 and SynB. The neurons appear to have expanded beyond the thickened epithelium. A-H are ventral views of embryos. Scale bars: 10 µm.
DEVEL
O
blastocoel (Fig. 5E,F). We conclude from these experiments that Notch signaling regulates apoptosis of one of the cells that results from the final mitotic division of neural progenitors.
DISCUSSION
The relationship between cell division and neural fate specification is an important aspect of early neurogenesis and a wide variety of patterns have arisen (Hartenstein and Stollewerk, 2015). We have previously established that S phase of the last cell cycle for the first neurons to differentiate in urchin embryos occurs at hatching (Garner et al., 2016). In addition, the onset of Sp-SoxC expression by these cells is coincident with S phase. Expression of GFP from a BAC clone containingSp-SoxC regulatory DNA enables observation of neural progenitors in living embryos. Our analysis of time-lapse sequences of embryos injected with the Sp-SoxC:GFP BAC indicates that, after mitosis, one of the progeny of the Sp-SoxC-expressing
[image:6.612.112.501.56.427.2]progenitors becomes apoptotic. These direct observations of apoptosis are also supported by the frequent appearance of GFP-labeled cell fragments in the blastocoel following injection with either the Sp-SoxC:GFP BAC or the 3×E-P:GFP Notch reporter. Apoptosis within embryonic neuronal cell lineages was first recognized when predictable cell death was shown to be part of the cellular fate maps inC. elegans(Hartenstein and Stollewerk, 2015). The pattern we have observed in urchin neurons appears to be the simplest form of asymmetric division; there are no intermediate progenitors, nor are there fixed lineages like those observed in several Drosophila neural types and in mammalian cortical neural progenitors (Rogulja-Ortmann et al., 2007; Lundell et al., 2003; Shen et al., 2006). Truman et al. (2010) describe asymmetric division of secondary lineages in the segmental nervous system of Drosophila, where division of neural progenitors produces a single neuron and a cell that undergoes apoptosis. In Drosophila and
Fig. 4. Notch signaling and the recruitment of neural progenitors to the APD or as postoral neurons.(A-D) In 20 h embryos, prior to hatching the APD is a thickened epithelium that expresses NK2.1. Initially, all cells of the APD express Sp-SoxC at a low level, but in the 28 h mesenchyme blastula expression is restricted to two to four cells along the dorsal margin. (E,F) In 28 h embryos treated with DAPT, the APD cells continue to express Sp-SoxC, indicating that Notch signaling is necessary to restrict expression to a few cells on the dorsal margin. (G,H) In 25 h embryos injected with mRNA encoding the Notch intracellular domain (NICD), the APD forms and expresses NK2.1, but none of the cells expresses Sp-SoxC. (I-K) To determine the role of Notch signaling in peripheral neurons, we examined the recruitment of Sp-SoxC-expressing cells as postoral ( po) neurons. (I) In control (DMSO) 28 h mesenchyme blastulae, there are two to four Sp-SoxC-expressing cells located at the base of the presumptive postoral arms. (J) In embryos treated with DAPT, there are 10-14 cells either in bilateral clusters or spread along the ventral surface. (K) In embryos injected with twoSp-DeltagRNAs and mRNA encoding Cas9, there are bilaterally disposed clusters of cells expressing Sp-SoxC where the postoral arms will form. (L) To determine the effect of inhibition of Notch after the initial phase of recruitment, embryos were treated with DAPT beginning at 24 h and the number of cells in adjacent pairs was counted. Arrowheads indicate pairs of cells. (M) Quantification of the number of cells in the APD expressing Sp-SoxC. (N) Quantification of the number of cells at the base of the postoral arms expressing Sp-SoxC. (O) Quantification of the proportion of Sp-SoxC-expressing cells in pairs after treatment with DAPT beginning at hatching (24 h). (M-O)n=5 embryos; s.e.m.
DEVEL
O
C. elegans, if apoptosis is blocked the cell differentiates as a neuron with minor effects on function. Thus, apoptosis appears to be a process that reduces the number of cells that differentiate as neurons. The apoptosis of one of the two progeny of the last division of neural progenitors in sea urchin embryos is a conserved process that limits the number of differentiating neurons.
Inhibition of the activating proteolysis of the Notch receptor or suppression of expression of the Notch ligand Sp-Delta results in an increase in the number of neural progenitors and neurons throughout the larval nervous system. This indicates that Notch signaling mediates the restriction of neural differentiation. The initial effect of inhibiting Notch signaling or suppressing Sp-Delta expression is on the recruitment of neural progenitors in the APD and the first peripheral neurons to form, the postoral neurons. However, use of the 3×E-P:GFP Notch reporter in combination with immunolocalization of SoxC indicates that cells with activated Notch signaling either express SoxC or are adjacent to cells expressing SoxC. These observations suggest that Notch also mediates fate determination during the asymmetric division of neural progenitors. This is supported by the preponderance of cell pairs resulting from inhibition of Notch signaling beginning at the time the final cell divisions are taking place. In addition, experiments in which Notch inhibition produces mostly pairs of ectodermal cells expressing GFP from the Sp-SoxC:GFP BAC indicate that Notch signaling functions in controlling apoptosis. In a similar manner, augmenting Notch signaling by co-injecting NICD mRNA leads to enhanced numbers of GFP-labeled cell fragments appearing in the blastocoel. These observations support a conclusion that Notch signaling is necessary and sufficient to induce apoptosis in SoxC-expressing neural progenitors. We have concluded from these data that Notch signaling has at least two effects on restriction of neurogenesis: first during recruitment of neural progenitors and again after the final mitotic division of neural progenitors.
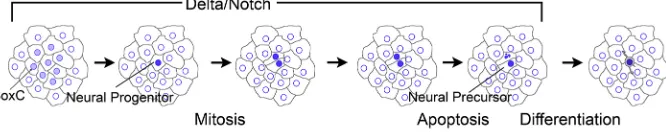
[image:7.612.50.399.56.362.2]We propose a model in which neurogenic potential is established in regions of ectoderm that have not experienced BMP signaling (LaPraz et al., 2009) (Fig. 6). Within the APD, ventral ectoderm, ciliary band, and dorsal ectoderm adjacent to the ciliary band, clusters of epithelial cells establish a neurogenic regulatory state. These cells express low levels of SoxC and Delta and activate Notch signaling in adjacent cells, which suppresses neurogenic potential in all but a few cells. This is an example of classical lateral inhibition (Lewis, 1998), and it produces a small number of neural progenitors co-expressing SoxC and Delta. Neural progenitors complete their last cell cycle and divide. The two progeny continue to express Delta, but one of the cells dominates and Notch signaling is activated in the adjacent cell, which undergoes apoptosis. This second phase of Notch signaling is an example of Notch regulating an asymmetric cell division (Torii, 2012; Truman et al., 2010). The resulting single neural precursor initiates expression of neural differentiation genes (Brn1/2/4, Zhfx1/Sip1, Tph, SynB) and differentiates as a neuron (Yaguchi et al., 2012; Wei et al., 2016; Garner et al., 2016). The mechanism that activates Notch signaling in one of the mitotic progeny is unclear. It could be attributable to a molecular asymmetry in which there is an unequal distribution of a component that suppresses Notch signaling (Jan and Jan, 1995; Bhat, 2014; Dong et al., 2012). Alternatively, direct Notch/Delta signaling between the two daughter cells is a potential mechanism. Throughout larval development, neurons are added to the ciliary band (Nakajima et al., 2004; Beer et al., 2001). How can neurons arise in larvae without dedicated neural progenitors that self-replicate and produce neurons? An explanation for this might be that the larval epidermis is in a regulatory state that retains the potential to enter a neurogenic pathway. Localization of endogenous Sp-SoxC in epidermal cells of plutei indicates there is a continuous appearance of cells expressing this protein. The model we have developed for the first neurons to appear in the embryo might be repeated throughout
Fig. 5. Inhibition or activation of Notch signaling alters the distribution of GFP-expressing neural progenitors in embryos injected with Sp-SoxC:GFP BAC. (A) Control preparations (DMSO) of 48 h embryos derived from eggs injected with Sp-SoxC:GFP have cells expressing GFP in ectoderm and in fragments within the blastocoel. (B-D) 48 h embryos derived from eggs injected with Sp-SoxC:GFP and then treated with DAPT beginning 3 h after fertilization have GFP-containing cells in pairs throughout the blastoderm. Single cells expressing GFP and GFP-containing fragments of cells are rare. (E) 48 h embryos derived from eggs co-injected with Sp-SoxC: GFP and mRNA encoding NICD have few cells in the blastoderm expressing GFP and a large number of GFP-containing cell fragments within the blastocoel.
(F) Quantification of the distribution of GFP-containing cells in embryos derived from eggs injected with Sp-SoxC:GFP and treated to inhibit or enhance Notch signaling. Cell fragments containing GFP in the blastocoel were not quantified. DMSO,n=51 cells, 11 embryos; DAPT,n=52 cells, 17 embryos; NICD,n=48 cells, 17 embryos. Scale bars: 10 µm.
DEVEL
O
larval development. Thus, patches of epidermal cells respond to localized signaling and initiate neurogenesis. The subsequent onset of Notch signaling restricts neurogenesis to a single cell.
One of the conserved features of a central neuroepithelium is the ability to produce abundant neurons by employing dedicated neural progenitors (Hartenstein and Stollewerk, 2015). A feature of dedicated neural progenitors or neuronal stem cells is that they occupy specific locations and produce a patterned array of neurons. The APD is a distinctive region of neuroectoderm that gives rise to a bilaterally symmetric apical organ (Byrne et al., 2007). The APD is established by ectodermal patterning resulting from Wnt and Nodal signaling (Yaguchi et al., 2006, 2016; Range and Wei, 2016). It is initially patterned as concentric rings of cells expressing Six3 and Hbn (Wei et al., 2009), with neurogenic potential being restricted to cells in the outer ring. Nodal secreted from the ventral ectoderm suppresses neurogenesis on the same side, and neurons only form bilaterally at the interface with dorsal ectoderm (Yaguchi et al., 2007, 2016). The APD appears to retain a similar initial patterning, as a circular pattern of SoxC-expressing cells in this region is evident in embryos in which CRISPR/Cas9 was used to mutate Sp-Delta. Thus, peripheral neurons arise in broad regions of ectoderm without apparent patterning, yet the neurons of the apical organ arise from a patterned neuroepithelium and produce a ganglion with bilateral symmetry. This suggests that there might be cells with stem-like properties within the APD, and it remains possible that cells with more complex patterns of asymmetry exist. It is notable that central nervous system patterning and the effects of Notch signaling are not uniform in other organisms (Truman et al., 2010). Neurogenesis in sea urchin embryos shares many of the features common to neurogenesis in all bilaterians (Hartenstein and Stollewerk, 2015). These include an initial phase during which ectoderm is patterned into neurogenic regions, including a specialized neuroepithelium that gives rise to a central nervous system. In most bilaterians, there are dedicated neural progenitors within the central nervous system and a widespread neurogenic potential is retained within the general ectoderm. In urchin embryos, broad regions of ectodermally derived epidermis appear to be a source of neurons added to the ciliary band throughout larval development. Bilaterians all have a Notch-regulated gene regulatory network, in which Notch signaling suppresses neurogenesis. In urchin embryos, Notch signaling is primarily responsible for the scattered pattern of neurogenic cells in the ectoderm. In bilaterians, there is limited proliferation of neural progenitors and neural progenitors complete their last cell cycle before there is a subsequent phase of movement for neural precursors and formation of axonal projections. A distinctive feature of urchin embryonic neurogenesis is that there is apparently no proliferation
of neural progenitors in the peripheral nervous system, nor are there dedicated neural progenitors. Furthermore, movement of neural precursors is very modest, since most arise close to the ciliary band. Thus, echinoid embryos appear to utilize a set of neurogenic mechanisms common to bilaterians.
A more complete understanding of the evolution of neurogenesis has been afforded by the emergence of novel models with tractable features of neurogenesis (Rentzsch et al., 2017; Lowe et al., 2003; Cunningham and Casey, 2014; Cheatle Jarvela et al., 2016; Ryan et al., 2016; Hudson, 2016). Among these models, the sea urchin embryo remains relevant as a means of reconstructing the ancestry of neurogenesis in deuterostomes. Urchin embryos employ a set of neurogenic mechanisms shared with other bilaterians, suggesting these are features that existed in the common ancestor of deuterostomes.
MATERIALS AND METHODS Embryo culture and injection
Gametes were collected from Strongylocentrotus purpuratus induced to spawn by shaking. Sperm was diluted 1:1000 in seawater prior to fertilization, and embryos were grown at 12-14°C. Staging follows Foltz et al. (2004). Eggs were prepared for microinjection as described previously (Krupke et al., 2014). Injection solutions of DNA, morpholino or RNA were prepared in 120 mM KCl. DNA injections followed Arnone et al. (2004). Bacterial cells containing the Sp-SoxC:GFP (Sp_3114N15) and Sp-Brn1/2/ 4:GFP (Sp_16C24) BACs were obtained from Eric Davidson (CalTech) and grown in LB plus chloramphenicol. BACs were purified using a NucleoBond BAC 100 Kit (Macherey-Nagel), linearized withNotI (New England BioLabs), and drop-dialyzed against TE buffer (10 mM Tris, 1 mM EDTA, pH 8.0) on 0.025 µm mixed cellulose ester membranes (EMD Millipore). Eggs were injected with 300-600 copies of BAC per 4 pl. A Notch reporter construct consisting of three tandem duplications of the E-P modules (Ransick and Davidson, 2006) adjacent to an eGFP open reading frame was synthesized by Biobasics (supplementary Materials and Methods). Eggs were injected with 400 copies of reporter along with a 5-fold excess of carrier DNA. Morpholino antisense oligonucleotides were obtained from GeneTools, diluted to a suitable stock concentration (1 mM), and further diluted and filtered before being used to fill injection needles.
Delta morpholino (5′-CAAGAAGGCAGTGCGGCCGATCCGT-3′) was
injected at 400 µM (Materna and Davidson, 2012). DAPT (N-[(3,5-difluorophenyl)acetyl]-L-alanyl-2-phenylglycine-1,1-dimethylethyl ester) (Tocris) was diluted to 50 mM in DMSO, then further diluted to 8 or 10 µM in embryo cultures. All experiments had at least three replicates, and treatment with DMSO alone was used as a control.
Live embryo imaging
[image:8.612.141.474.61.127.2]Live images were made using a Leica DM6000 epifluorescence microscope and Hamamatsu Orca camera controlled withμManager (1.4; https://micro-manager.org/). Injected embryos were pipetted onto NewSilane Adhesive-Coated Slides (Newcomer Supply) and trapped under a glass coverslip Fig. 6. Model of recruitment and final mitotic division of neural progenitors in sea urchin embryos.Initially, neurogenic potential is established in regions of ectoderm that have not experienced BMP signaling. Within the APD, ventral ectoderm, ciliary band, and dorsal ectoderm adjacent to the ciliary band, clusters of epithelial cells establish a neurogenic regulatory state. These cells express low levels of Sp-SoxC and Sp-Delta and activate Notch signaling in adjacent cells, which suppresses neurogenic potential in all but a few scattered cells. This lateral inhibition produces a small number of neural progenitors co-expressing Sp-SoxC and Sp-Delta. Neural progenitors complete their last cell cycle and divide, with the progeny continuing to express Sp-Delta. One of the cells dominates and Notch signaling is activated in the adjacent cell, which undergoes apoptosis. The single neural precursor resulting from this asymmetric division initiates expression of neural differentiation genes and differentiates as a neuron. The model is consistent with our data for a single interval of Notch-Delta interactions that spans the recruitment and asymmetric mitosis of a neural progenitor.
DEVEL
O
attached along two edges with double-sided adhesive tape (3 M). Paraffin oil was applied to the open edges of the coverslip to reduce evaporation, and the room temperature was controlled to 16°C. Embryos could routinely be maintained for 6-8 h. Movies were prepared using ImageJ (1.51 h) and rendered to MP4 format in Adobe Photoshop CS6.
Immunofluorescence microscopy
S. purpuratusembryos were collected at the desired time point and fixed for 5-10 min in PEM buffer with 4% paraformaldehyde (Vielkind and Swierenga, 1989). Embryos were washed with PBS, blocked for 1 h in SuperBlock (ThermoFisher Scientific), probed with primary antibody, and washed three times with PBS. Alexa Fluor fluorescent secondary antibodies (Invitrogen; 1:1500) were used to visualize antibody labeling on a Zeiss LSM 700 confocal microscope. Quantification and image analysis was performed using ZEN 2009 or ZEN-Lite software (Carl Zeiss). Cell counts were made by manually counting fluorescently labeled cells in each image of a through-focus stack. Data were analyzed with a one-way ANOVA with Dunnett’s or Tukey’s post-tests using GraphPad Prism (5.04). ImageJ, ZEN-Lite or Adobe Photoshop was used to adjust channel color, image contrast and brightness and for final editing. Antibodies employed were: anti-SynB (1E11) (Nakajima et al., 2004), 1:10; anti-SoxC (Garner et al., 2016), 1:10; anti-GFP (Abcam, ab6673), 1:400; anti-cleaved Caspase3 (Asp175; Cell Signaling Technology, #9661), 1:400; anti-Hnf6 (Yaguchi et al., 2010), 1:400; anti-NK2.1 (Takacs et al., 2004), 1:400; anti-CRISPR-Cas9 (7A9-3A3; Abcam, ab191468), 1:400.
Cas9 and gRNA preparations
Procedures for CRISPR/Cas9 mutation of theDeltagene followed Lin and Su (2016) and are described in detail in the supplementary Materials and Methods and Fig. S4.
Acknowledgements
We thank Andy Ransick, Eric Davidson and Isabelle Peter for generously providing reagents and unpublished data. Andy Ransick also provided essential technical advice.
Competing interests
The authors declare no competing or financial interests.
Author contributions
Conceptualization: R.D.B.; Methodology: D.O.M., J.T., R.D.B.; Validation: D.O.M., J.T., R.D.B.; Formal analysis: D.O.M., R.D.B.; Investigation: D.O.M., J.T., R.D.B.; Resources: D.O.M., J.T., R.D.B.; Data curation: R.D.B.; Writing - original draft: R.D.B.; Writing - review & editing: D.O.M., J.T., R.D.B.; Visualization: R.D.B.; Supervision: R.D.B.; Project administration: R.D.B.; Funding acquisition: R.D.B.
Funding
This work was supported by a Discovery Grant from the Natural Sciences and Engineering Research Council of Canada (2016-03737) to R.D.B.
Supplementary information
Supplementary information available online at
http://dev.biologists.org/lookup/doi/10.1242/dev.151720.supplemental
References
Arnone, M. I., Dmochowski, I. J. and Gache, C.(2004). Using reporter genes to study cis-regulatory elements.Method Cell Biol.74, 621-652.
Beer, A.-J., Moss, C. and Thorndyke, M.(2001). Development of serotonin-like and SALMFamide-like immunoreactivity in the nervous system of the sea urchin Psammechinus miliaris.Biol. Bull.200, 268-280.
Bhat, K. M.(2014). Notch signaling acts before cell division to promote asymmetric cleavage and cell fate of neural precursor cells.Sci. Signal.7, ra101.
Burke, R. D., Angerer, L. M., Elphick, M. R., Humphrey, G. W., Yaguchi, S., Kiyama, T., Liang, S., Mu, X., Agca, C., Klein, W. H. et al.(2006). A genomic view of the sea urchin nervous system.Dev. Biol.300, 434-460.
Burke, R. D., Moller, D. J., Krupke, O. A. and Taylor, V. J.(2014). Sea urchin neural development and the metazoan paradigm of neurogenesis.Genesis52, 208-221.
Byrne, M., Nakajima, Y., Chee, F. C. and Burke, R. D.(2007). Apical organs in echinoderm larvae: insights into larval evolution in the Ambulacraria.Evol. Dev.9, 432-445.
Cheatle Jarvela, A. M., Yankura, K. A. and Hinman, V. F. (2016). A gene regulatory network for apical organ neurogenesis and its spatial control in sea star embryos.Development143, 4214-4223.
Collier, J. R., Monk, N. A. M., Maini, P. K. and Lewis, J. H.(1996). Pattern formation by lateral inhibition with feedback: a mathematical model of Delta-Notch intercellular signalling.J. Theor. Biol.183, 429-446.
Corbin, J. G., Gaiano, N., Juliano, S. L., Poluch, S., Stancik, E. and Haydar, T. F.
(2008). Regulation of neural progenitor cell development in the nervous system.
J. Neurochem.106, 2272-2287.
Croce, J. C. and McClay, D. R.(2010). Dynamics of Delta/Notch signaling on endomesoderm segregation in the sea urchin embryo.Development137, 83-91.
Cunningham, D. and Casey, E. S.(2014). Spatiotemporal development of the embryonic nervous system of Saccoglossus kowalevskii.Dev. Biol.386, 252-263.
Dong, Z. Q., Yang, N., Yeo, S.-Y., Chitnis, A. and Guo, S.(2012). Intralineage directional notch signaling regulates self-renewal and differentiation of asymmetrically dividing radial glia.Neuron74, 65-78.
Erwin, D. H.(2015). Early metazoan life: divergence, environment and ecology.
Philos. Trans. R. Soc. B Biol. Sci.370, 20150036.
Foltz, K. R., Adams, N. L. and Runft, L. L.(2004). Echinoderm eggs and embryos: procurement and culture.Methods Cell Biol.74, 40-74.
Gaiano, N. and Fishell, G.(2002). The role of Notch in promoting glial and neural stem cell fates.Annu. Rev. Neurosci.25, 471-490.
Gaiano, N., Nye, J. S. and Fishell, G.(2000). Radial glial identity is promoted by Notch1 signaling in the murine forebrain.Neuron26, 395-404.
Garner, S., Zysk, I., Byrne, G., Kramer, M., Moller, D., Taylor, V. and Burke, R. D.
(2016). Neurogenesis in sea urchin embryos and the diversity of deuterostome neurogenic mechanisms.Development143, 286-297.
Hansson, E. M., Teixeira, A. I., Gustafsson, M. V., Dohda, T., Chapman, G., Meletis, K., Muhr, J. and Lendahl, U.(2006). Recording Notch signaling in real time.Dev. Neurosci.28, 118-127.
Hartenstein, V. and Stollewerk, A.(2015). The evolution of early neurogenesis.
Dev. Cell32, 390-407.
Hudson, C.(2016). The central nervous system of ascidian larvae.Wiley Interdiscip. Rev. Dev. Biol.5, 538-561.
Jan, Y. N. and Jan, L. Y.(1995). Maggots hair and bugs eye-role of cell-interactions and intrinsic-factors in cell fate specification.Neuron14, 1-5.
Knoblich, J. A.(2008). Mechanisms of asymmetric stem cell division.Cell132, 583-597.
Krupke, O., Yaguchi, S., Yaguchi, J. and Burke, R. D.(2014). Imaging neural development in embryonic and larval sea urchins.Methods Mol. Biol.1128, 147-160.
Lapraz, F., Besnardeau, L. and Lepage, T.(2009). Patterning of the dorsal-ventral axis in echinoderms: insights into the evolution of the BMP-chordin signaling network.PLoS Biol.7, e1000248.
Lee, E.-C., Yu, D. G., de Velasco, J. M., Tessarollo, L., Swing, D. A., Court, D. L., Jenkins, N. A. and Copeland, N. G.(2001). A highly efficient Escherichia coli-based chromosome engineering system adapted for recombinogenic targeting and subcloning of BAC DNA.Genomics73, 56-65.
Lewis, J.(1998). Notch signalling and the control of cell fate choices in vertebrates.
Semin. Cell Dev. Biol.9, 583-589.
Liebeskind, B. J., Hillis, D. M., Zakon, H. H. and Hofmann, H. A.(2016). Complex homology and the evolution of nervous systems.Trends Ecol. Evol.31, 127-135.
Lin, C.-Y. and Su, Y.-H.(2016). Genome editing in sea urchin embryos by using a CRISPR/Cas9 system.Dev. Biol.409, 420-428.
Louvi, A. and Artavanis-Tsakonas, S.(2006). Notch signalling in vertebrate neural development.Nat. Rev. Neurosci.7, 93-102.
Lowe, C. J., Wu, M., Salic, A., Evans, L., Lander, E., Stange-Thomann, N., Gruber, C. E., Gerhart, J. and Kirschner, M.(2003). Anteroposterior patterning in hemichordates and the origins of the chordate nervous system.Cell113, 853-865.
Lundell, M. J., Lee, H. K., Pérez, E. and Chadwell, L.(2003). The regulation of apoptosis by numb/notch signaling in the serotonin lineage of Drosophila.
Development130, 4109-4121.
Mali, P., Yang, L. H., Esvelt, K. M., Aach, J., Guell, M., DiCarlo, J. E., Norville, J. E. and Church, G. M.(2013). RNA-guided human genome engineering via Cas9.Science339, 823-826.
Materna, S. C. and Davidson, E. H.(2012). A comprehensive analysis of Delta signaling in pre-gastrular sea urchin embryos.Dev. Biol.364, 77-87.
Moroz, L. L., Kocot, K. M., Citarella, M. R., Dosung, S., Norekian, T. P., Povolotskaya, I. S., Grigorenko, A. P., Dailey, C., Berezikov, E., Buckley, K. M. et al.(2014). The ctenophore genome and the evolutionary origins of neural systems.Nature510, 109-114.
Nakajima, Y., Kaneko, H., Murray, G. and Burke, R. D.(2004). Divergent patterns of neural development in larval echinoids and asteroids.Evol. Dev.6, 95-104.
Peterson, K. J. and Eernisse, D. J. (2016). The phylogeny, evolutionary developmental biology, and paleobiology of the Deuterostomia: 25 years of new techniques, new discoveries, and new ideas.Org. Divers. Evol.16, 401-418.
Pierfelice, T., Alberi, L. and Gaiano, N.(2011). Notch in the vertebrate nervous system: an old dog with new tricks.Neuron69, 840-855.
DEVEL
O
Range, R. C. and Wei, Z.(2016). An anterior signaling center patterns and sizes the anterior neuroectoderm of the sea urchin embryo.Development143, 1523-1533.
Ransick, A. and Davidson, E. H. (2006). cis-regulatory processing of Notch signaling input to the sea urchin glial cells missing gene during mesoderm specification.Dev. Biol.297, 587-602.
Rentzsch, F., Layden, M. and Manuel, M.(2017). The cellular and molecular basis of cnidarian neurogenesis.Wiley Interdiscip. Rev. Dev. Biol.6, doi: 10.1002/ wdev.257.
Rogulja-Ortmann, A., Luer, K., Seibert, J., Rickert, C. and Technau, G. M.
(2007). Programmed cell death in the embryonic central nervous system of Drosophila melanogaster.Development134, 105-116.
Rottinger, E., Croce, J., Lhomond, G., Besnardeau, L., Gache, C. and Lepage, T.
(2006). Nemo-like kinase (NLK) acts downstream of Notch/Delta signalling to downregulate TCF during mesoderm induction in the sea urchin embryo.
Development133, 4341-4353.
Ryan, K., Lu, Z. Y. and Meinertzhagen, I. A.(2016). The CNS connectome of a tadpole larva of Ciona intestinalis (L.) highlights sidedness in the brain of a chordate sibling.Elife5, e16962.
Saudemont, A., Haillot, E., Mekpoh, F., Bessodes, N., Quirin, M., Lapraz, F., Duboc, V., Rottinger, E., Range, R., Oisel, A. et al.(2010). Ancestral regulatory circuits governing ectoderm patterning downstream of nodal and BMP2/4 revealed by gene regulatory network analysis in an echinoderm.PLoS Genet.
6, e1001259.
Shen, Q., Wang, Y., Dimos, J. T., Fasano, C. A., Phoenix, T. N., Lemischka, I. R., Ivanova, N. B., Stifani, S., Morrisey, E. E. and Temple, S.(2006). The timing of cortical neurogenesis is encoded within lineages of individual progenitor cells.
Nat. Neurosci.9, 743-751.
Sherwood, D. R. and McClay, D. R.(1999). LvNotch signaling mediates secondary mesenchyme specification in the sea urchin embryo.Development126, 1703-1713.
Sweet, H. C., Gehring, M. and Ettensohn, C. A.(2002). LvDelta is a mesoderm-inducing signal in the sea urchin embryo and can endow blastomeres with organizer-like properties.Development129, 1945-1955.
Takacs, C. M., Amore, G., Oliveri, P., Poustka, A. J., Wang, D., Burke, R. D. and Peterson, K. J.(2004). Expression of an NK2 homeodomain gene in the apical ectoderm defines a new territory in the early sea urchin embryo.Dev. Biol.269, 152-164.
Torii, K. U.(2012). Two-dimensional spatial patterning in developmental systems.
Trends Cell Biol.22, 438-446.
Truman, J. W., Moats, W., Altman, J., Marin, E. C. and Williams, D. W.(2010). Role of Notch signaling in establishing the hemilineages of secondary neurons in Drosophila melanogaster.Development137, 53-61.
Vielkind, U. and Swierenga, S. H. (1989). A simple fixation procedure for immunofluorescent detection of different cytoskeletal components within the same cell.Histochemistry91, 81-88.
Wei, Z., Yaguchi, J., Yaguchi, S., Angerer, R. C. and Angerer, L. M.(2009). The sea urchin animal pole domain is a Six3-dependent neurogenic patterning center.
Development136, 1179-1189.
Wei, Z., Angerer, R. C. and Angerer, L. M.(2011). Direct development of neurons within foregut endoderm of sea urchin embryos.Proc. Natl. Acad. Sci. USA108, 9143-9147.
Wei, Z., Range, R., Angerer, R. and Angerer, L. (2012). Axial patterning interactions in the sea urchin embryo: suppression of nodal by Wnt1 signaling.
Development139, 1662-1669.
Wei, Z., Angerer, L. M. and Angerer, R. C.(2016). Neurogenic gene regulatory pathways in the sea urchin embryo.Development143, 298-305.
Yaguchi, S., Yaguchi, J. and Burke, R. D.(2006). Specification of ectoderm restricts the size of the animal plate and patterns neurogenesis in sea urchin embryos.Development133, 2337-2346.
Yaguchi, S., Yaguchi, J. and Burke, R. D. (2007). Sp-Smad2/3 mediates patterning of neurogenic ectoderm by nodal in the sea urchin embryo.Dev. Biol.302, 494-503.
Yaguchi, S., Yaguchi, J., Angerer, R. C., Angerer, L. M. and Burke, R. D.(2010). TGF beta signaling positions the ciliary band and patterns neurons in the sea urchin embryo.Dev. Biol.347, 71-81.
Yaguchi, S., Yaguchi, J., Wei, Z., Jin, Y. H., Angerer, L. M. and Inaba, K.(2011). Fez function is required to maintain the size of the animal plate in the sea urchin embryo.Development138, 4233-4243.
Yaguchi, J., Angerer, L. M., Inaba, K. and Yaguchi, S.(2012). Zinc finger homeobox is required for the differentiation of serotonergic neurons in the sea urchin embryo.Dev. Biol.363, 74-83.
Yaguchi, J., Takeda, N., Inaba, K. and Yaguchi, S.(2016). Cooperative Wnt-nodal signals regulate the patterning of anterior neuroectoderm.PLoS Genet. 12, e1006001.
Yoon, K. and Gaiano, N.(2005). Notch signaling in the mammalian central nervous system: insights from mouse mutants.Nat. Neurosci.8, 709-715.