Transient gene transfer and expression of Smad7 prevents bleomycin induced lung fibrosis in mice
Full text
Figure
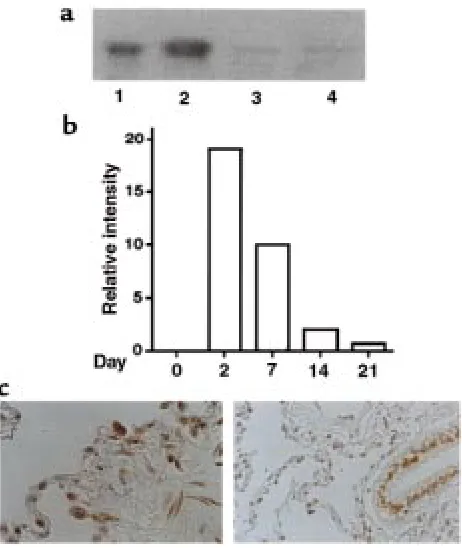

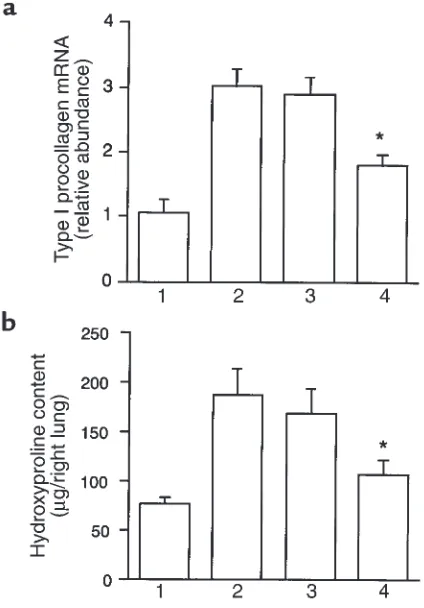
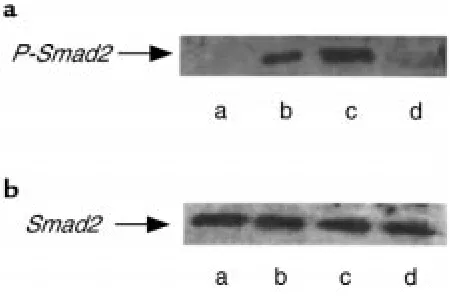
Related documents
Adiponectin significantly enhanced NK cell ability to kill target cells in lean controls, compared to when adiponectin was absent (42% vs 47% target cells killed, n = 10, **p =
Keywords : Technical Change, Institutional Change, Cognition, Knowledge Creation, Environmental.. Management,
• The ease of maintenance of calibration of sensors and instruments was judged to meet the criterion for remote operation but a number of the fixtures in the AWTP were
Here, we describe the case of a 33-year-old Japanese female whose pelvis had contained retained lipiodized oil that had been suspected as residual metallic material.. The
Our proposal signifies that people need to change their mentality about politics and join it in true professional spirit so that we can be a developed nation and present an example
Already now we use routinely Cinematic Rendering for teaching human anatomy on the base of CT and MR 3D data sets (virtual anatomy), because Cinematic Rendering is able to
There is a need for academic programs of nursing to expand distance course and curricular offering in order to meet the rising demands for nurses in the workforce [4] [19] [20].
In order to satisfy this demand while main- taining low C emission, the effects of different agricultural management practices such as the use of no-tillage and alternative
