Expression patterns and immunohistochemical localization of
PITX2B transcription factor in the developing mouse heart
FRANCISCO HERNANDEZ-TORRES*, DIEGO FRANCO, AMELIA E. ARANEGA and FRANCISCO NAVARRO*
Department of Experimental Biology, Faculty of Experimental Sciences, University of Jaen, Jaen, Spain.
ABSTRACT The Pitx2 gene is involved in the establishment of vertebrate left-right axis with an important role in subsequent heart organogenesis. Mutations in the Pitx2 gene have been associ-ated with Axenfeld-Rieger syndrome, which is characterized by ocular, craniofacial, and umbilical anomalies, as well as cardiac defects. In addition, recent data have unravelled a molecular link between PITX2 loss of function and atrial fibrillation (AF), supporting an important role of Pitx2 not only in development but also in heart homeostasis. Three PITX2 isoforms have been described in mice: PITX2A, PITX2B, and PITX2C. During heart organogenesis, PITX2C seems to play a determi-nant role in left–right signalling from early somitogenesis onwards. However the participation of the PITX2A and/or PITX2B isoforms during cardiogenesis is controversial. Here we report for the first time that the Pitx2a and Pitx2b isoforms are jointly expressed with the Pitx2c isoform during heart development. Interestingly, in terms of relative quantification of mRNA, the Pitx2b and Pitx2c isoforms display similar expression profiles during cardiogenesis, decreasing with further develop-ment but maintaining their expression until adult stages. Moreover, a detailed analysis of PITX2B protein during cardiac development shows that PITX2B is dynamically expressed in the developing ventricular septum and asymmetrically expressed in the tricuspid valve primordia, suggesting a putative role of the PITX2B isoform during ventricular septation as well as in the maturation of the right portion of the atrioventricular canal.
KEY WORDS:
Pitx2 isoforms, heart development, mouse and ventricular septation
The Pitx2 gene is a member of the Bicoid-like homeobox family (Semina et al., 1996). During the last fifteen years several stud-ies reported the implication of Pitx2 in the correct establishment of vertebrate left-right axis. Left-right asymmetry is established through a molecular cascade that gives rise to Pitx2 asymmetric expression in the Lateral Plate Mesoderm (LPM) at early stages of development (Logan et al., 1998). This expression determines left/right polarity of mesoderm-derived organs such as heart, gut, and stomach (Campione et al., 1999; Logan et al., 1998; Ryan et al., 1998; Yoshioka et al., 1998), supporting that Pitx2 could be the molecular transducer of embryonic left–right signalling at the organ level during early stages of development. Mutations in the Pitx2 gene have been associated with Axenfeld-Rieger syndrome, which is characterized by ocular, craniofacial, and umbilical anomalies (Semina et al., 1996) as well as cardiac defects, such as atrial septal defects, atrio-ventricular valve defects, and conduction
www.intjdevbiol.com
*Address correspondence to: Francisco Navarro-Gómez. Department of Experimental Biology, Faculty of Experimental Sciences, University of Jaen, Paraje de
Las Lagunillas, s/n, 23071, Jaen, Spain. Tel: +34-953-212771. Fax: +34-953-211875. E-mail: [email protected] or
Francisco Hernández-Torres. Department of Experimental Biology, Faculty of Experimental Sciences, University of Jaen, Paraje de Las Lagunillas, s/n, 23071, Jaen, Spain. Tel: +34-953-213056. Fax: +34-953-211875. E-mail: [email protected]
Supplementary Material (one figure) for this paper is available at: http://dx.doi.org/10.1387/ijdb.140224fh
Accepted: 5 February 2015.
ISSN: Online 1696-3547, Print 0214-6282 © 2015 UBC Press
Printed in Spain
Abbreviations used in this paper: Pitx2, pituitary homeobox 2 gene; AF, atrial
fibrilla-tion; LPM, lateral plate mesoderm; OFT, outflow tract; AS, alternative splicing. abnormalities (Cunningham et al., 1998; Mammi et al., 1998; Tsai, Grajewski, 1994).
link between Pitx2c and AF has been recently reported (Chinchilla et al., 2011; Kirchhof et al., 2011; Wang et al., 2010). Taken together, all these data suggest an important role of Pitx2c in heart develop-ment and homeostasis. However, Pitx2a and/or Pitx2b contribution during cardiogenesis cannot be completely discarded. Actually, a role for PITX2A during outflow tract (OFT) development has been suggested (Kioussi et al., 2002). In addition, Hjalt et al., (Hjalt et al., 2000), by using an antibody which could detect all PITX2 pro-tein isoforms, detected the presence of PITX2 propro-tein in the right atrial and ventricular wall at stage E12.5 in mouse, contradicting the Pitx2c mRNA expression pattern previously described (Cam-pione et al., 1999; Logan et al., 1998; Ryan et al., 1998; Yoshioka et al., 1998). Moreover, analysis of the fate of Pitx2 daughter cells
in mutant embryos that only express Pitx2a and Pitx2b isoforms (Pitx2 δabccreneo;δc and Rosa26 reporter trans-heterozygotes) (Liu et al., 2002) revealed that fewer lacZ-positive cells were found in the right ventricular and inner curvature myocardium of Pitx2 mu-tant embryos at both 14.5 dpc and 18.5 dpc. Finally, comparative anatomical analysis of hearts from Pitx2 specific null mutant mice (Pitx2abc -/- (Gage et al., 1999), Pitx2ab -/- (Liu et al., 2001) and Pitx2c -/- (Liu et al., 2002)) leads to speculate that Pitx2a and/or Pitx2b could play a role in the development of the OFT (Franco et al., 2003). In spite of all these evidences, the implication of Pitx2a and or Pitx2b isoforms during cardiogenesis still remains elusive. Here we show, for the first time, experimental data demonstrating that Pitx2a and Pitx2b isoforms are also expressed during heart
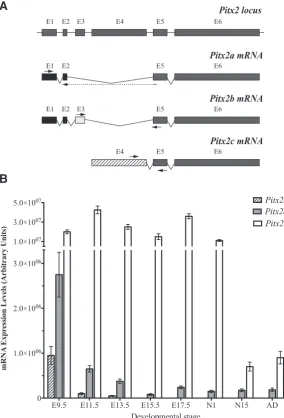
Fig. 1. Pitx2 mRNA expression levels in cardiogenesis.(A) Schematic represen-tation of the strategy followed in order to measure specifically each Pitx2 isoform mRNA. Arrows indicate the primers annealing place used to amplify each isoform.
(B) Pitx2a, Pitx2b and Pitx2c mRNA expression levels during mouse heart develop-ment as measured by quantitative RT-PCR. Data are expressed as ratio of each Pitx2
isoform mRNA to b-actin mRNA. Embryonic day (E) 9.5 to 17.5; N1, 1 day neonate mouse heart; N15, 15 day neonate heart; Ad, adult mouse heart.
development. In addition, we report a detailed analysis of PITX2B protein during cardiac development showing that PITX2B is dynamically expressed in the developing ventricular septum and asymmetrically expressed in the tricuspid valve primordia.
Results
Pitx2a, Pitx2b and Pitx2c are dynamically expressed during mouse heart development
In order to analyse the mRNA expression pattern of each Pitx2 isoform during cardiogenesis, we performed qRT-PCR experiments in embryonic, neonatal and adult mouse whole hearts. As shown in Fig. 1 the three murine Pitx2 isoforms are expressed during cardiogenesis. Pitx2c expression levels are clearly higher than Pitx2a and Pitx2b isoforms, displaying different dynamic expression profiles. Thus, Pitx2c presents a maximal expression level at stage E11.5, decreasing at E13.5 and E15.5, increasing again at E17.5 and, subsequently decreasing at neonatal and adult stages where Pitx2c displayed its lowest expression levels. On the other hand, Pitx2b presents a maximal ex-pression level at stage E9.5, decreasing strongly at E11.5 and maintaining similar expression levels until adulthood. Although Pitx2c is always expressed at higher levels than Pitx2b, the difference in terms of fold-times varies through heart development. Thus this difference is 7.3x, 65.4x, 66.6x, 187.5x, 150x, 73.3x, 4x and 4,8x for the stages of development E9.5, E11.5, E13.5, E15.5, E17.5, N1, N15 and adult, respectively. Finally, Pitx2a, the lowest expressed isoform, also presents a maximal expression level at E9.5 stage, followed by a strong decrease at stage E11.5 but, contrary to Pitx2c and Pitx2b, Pitx2a expression was not detected from E13.5 stage onwards
Taken together, our results demonstrate for the first time that, in spite of Pitx2c presenting the highest levels of mRNA expression, Pitx2a and Pitx2b are coexpressed simultaneously during cardiogenesis. Moreover we also show that, although Pitx2a, Pitx2b and Pitx2c are coex-pressed during early cardiogenesis, only Pitx2b and Pitx2c maintain their expression during embryonic, foetal, neonatal and adult stages.
PITX2B protein expression profile during cardiogenesis The presence of Pitx2b mRNA during heart develop-ment and its maintenance until adult stage suggests an
E1 E2 E3 E4 E5 E6
G
B
C
D
E
F
H
I
J
K
L
M
important role of this isoform in cardiogenesis. Thus, in order to analyse the distribution of murine PITX2B protein during cardiac development, we performed immunohistochemistry assays by us-ing specific antibodies against PITX2B protein (Hernández-Torres et al., 2008). In order to demonstrate the specificity of anitPITX2B antibodies, we used immortalized cardiomyocytes isolated from mice at stage E9.5 (p38a+/+ and p38a-/-) (Adams et al., 2000) and
demonstrated that lack of p38a Map Kinase avoid Pitx2a, Pitx2b
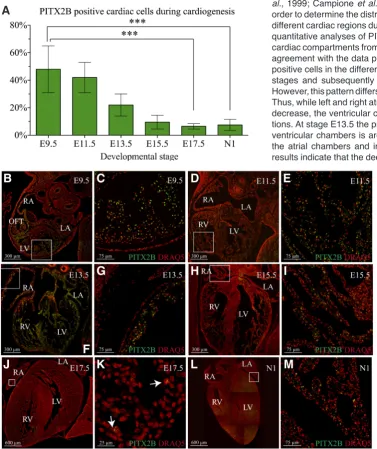
and Pitx2c mRNA expression (Supplementary Fig. 1). In addition the results in Supplementary Fig. 1 B-F, demonstrate the specificity of antiPITX2B antibodies. Consequently, the use of these antiPI-TX2B antibodies allowed us to quantify the proportion of PIantiPI-TX2B positive nuclei in the heart at different developmental stages. As illustrated in Fig. 2 the PITX2B positive cardiac cell profile is in agreement with the mRNA expression profile described above. Thus, the number of PITX2B positive cells is higher at early stages of development, showing a maximal number of positive cells at
stage E9.5. Subsequently, the proportion of PITX2B positive nuclei decreases until foetal and neonatal stages, where the proportion of PITX2B positive nuclei is around 6~7%.
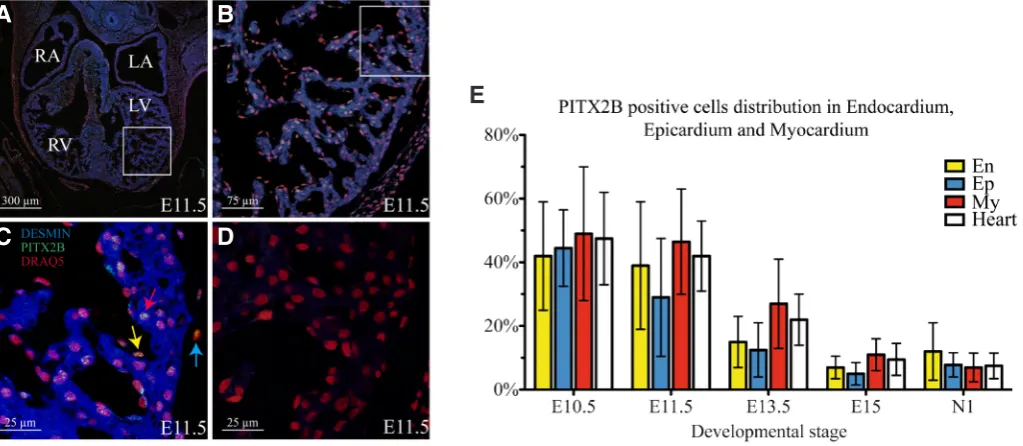
Co-immunolocalization experiments with a myocardial marker (DESMIN) indicated that PITX2B protein is present not only in myocardial cells but also in cells located in endocardium and epicardium (Fig. 3A-E). In addition, myocardial, endocardial and epicardial quantitative analyses showed that the proportion of PITX2B positive cells also decreases during development in all three tissue layers, showing a maximal number of positive cells in early stages of development and decreasing as development progresses (Fig. 3E).
Changes in PITX2B protein distribution within different cardiac compartments during development
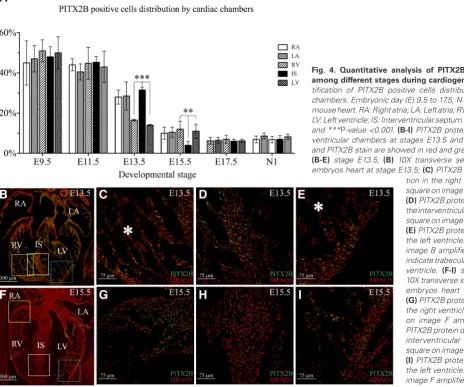
Previous experiments have clearly determined that Pitx2c mRNA is asymmetrically distributed in heart, and is restricted to specific heart compartments as development progresses (Campione et al., 1999; Campione et al., 2001; Franco et al., 2003). Thus, in order to determine the distribution of PITX2B positive cells among different cardiac regions during heart development, we performed quantitative analyses of PITX2B positive cells within the different cardiac compartments from E9.5 to neonatal stages (Fig. 4 A-I). In agreement with the data presented above, the number of Pitx2b positive cells in the different heart regions is higher during earlier stages and subsequently decreases with further development. However, this pattern differs depending on the specific heart areas. Thus, while left and right atria show a constant and homogeneous decrease, the ventricular chambers display some regional varia-tions. At stage E13.5 the proportion of Pitx2b positive cells within ventricular chambers is around 14~16%, but around 28~31% in
the atrial chambers and interventricular septum. Notably these results indicate that the decrease in Pitx2b positive cells at E13.5
Fig. 2. PITX2B protein expression analysis during heart morphogenesis. (A) Quantification of PITX2B positive cells in hearts from different stages dur-ing cardiogenesis. Embryonic day (E) 9.5 to 17.5; N1: 1 day neonate mouse heart.
(B-M) Nuclear immunohistochemical stain obtained in transverse heart sections from mouse embryos in different stages of development by using specific antiPITX2B antibody. Nucleus and PITX2B stain are showed in red and green respectively.
(B-C) stage E9.5; D-E) stage E11.5; (F-G) stage E13.5; (H-I) stage E15.5; (J-K)
is larger in the left and right ventricles as compared with atrial and interventricular septum compartments. However at E15.5 we observe a drastic decrease in the number of Pitx2b positive cells in the interventricular septum (only 4%) compared with a less extensive decrease in atrial (≈10%) and ventricular chambers (≈11~12%). Contrary to this, at E17.5 and neonatal stages,
com-parable proportions of Pitx2b positive nuclei (≈5~6%) are shown
in all cardiac structures (Fig. 4I). Overall, these findings indicate that during the developmental window comprising interventricular septum formation (E11.5-E15.5) (Miquerol et al., 2012) PITX2B positive cells displayed a dynamic distribution within this ventricular compartment. At E13.5, we found that a major proportion of PITX2B ventricular positive nuclei are located in the interventricular septum and these cells shift to minimal PITX2B expression at E15.5 when ventricular septation is fully completed (Miquerol et al., 2012).
In addition, and based on recent reports that pointed out a pos-sible participation of PITX2 in the development of atrioventricular (AV) canal (Tessari et al., 2008), we investigated whether PITX2B could be involved in AV canal formation by analysing the number of PITX2B positive cells in those structures. At stages E9.5 and E11.5 the proportion of PITX2B positive mesenchymal cells in AV cushions is clearly low, around 4% (Fig. 5 A,B,E-H). Neverthe-less, at stage E13.5, when the mural and septal portions of valve primordia can be observed (Lincoln et al., 2004), differences in the structures derived from AV canal cushions were detected (Fig. 5 C,D,I-K). Thus, while mural portions of tricuspid valves (TV), close to the right ventricular wall on Fig. 5J, presented a proportion of PITX2B positive cells similar to that estimated for whole heart at this stage (around 23%), septal portions of valves displayed a value of PITX2B positive cells at around 3% (Fig. 5 C,J). However, for the mitral valve (MV) analogues, neither presented a significantly
high proportion of PITX2B positive cells, at around 3~7% for both
mural and septal portions (Fig. 5 D,K). This PITX2B expression profile suggests a role for PITX2B in the establishment of different components of the tricuspid valve during development, and this is the only sign of asymmetry for PITX2B protein distribution that we have detected during heart development.
Discussion
It is well established that Pitx2 plays a crucial role in the es-tablishment of vertebrate left-right axis determination, acting as a molecular transducer of embryonic left–right signalling at the organ level during early stages of development, including the heart (Franco et al., 2003). Previous studies have shown that, although Pitx2a and Pitx2b isoforms are symmetrically co-expressed with Pitx2c in certain regions of the embryo, only Pitx2c is asymmetri-cally expressed in the lateral plate mesoderm (LPM), heart and gut (Kitamura et al., 1999; Liu et al., 2001; Schweickert et al., 2000). The importance of Pitx2 in heart development gathered strength when, concurrently, Pitx2 specific null mice mutants (Pitx2abc -/- (Gage et al., 1999), Pitx2ab -/- (Liu et al., 2001) and Pitx2c -/- (Liu et al., 2002)) were developed. Pitx2 mutant mice for all three isoforms (dabcnull) display several cardiac malformations such as
right sino-atrial isomerism and abnormalities of atrioventricular and ventriculoarterial connections. Since similar cardiac malformations have been described in Pitx2c mutant mice while no cardiac anoma-lies have been detected in Pitx2a/b mutant mice, the relevance of Pitx2c isoform in the heart has been extensively accepted (Liu et al., 2001; Liu et al., 2002). However, several sets of controversial data support a putative role for Pitx2a/b in this process. First, Kioussi et al., have suggested a role for PITX2A during outflow
Fig. 3. Quantitative analysis of PITX2B positive cells in endocardium, myocardium and epicardium.(A-D) Nuclear and cytoplasm immunohisto-chemical stain obtained in transverse sections of mouse embryos at stage E11.5 by using specific antiPITX2B and antiDESMIN antibodies respectively.
(A) 10X transverse section of mouse embryos heart at stage E11.5. (B) White square on image A amplified 40X. (C) White square on image B ampli-fied 120X. Nucleus, PITX2B and DESMIN stain are showed in red, green and blue respectively. Yellow, red and blue arrows indicate PITX2B positive nucleus from endocardium, myocardium and epicardium respectively (D) Negative control. (E) Quantificationof PITX2B positive cells in endocardium, epicardium and myocardium during cardiogenesis. Embryonic day (E) 10.5 to 15.5; N1: 1 day neonate mouse heart; En, endocardium; Ep, epicardium; My, myocardium; WH, Whole heart. RA: Right atria; LA: Left atria; RV: Right ventricle; LV: Left ventricle; IS: Interventricular septum.
B
C
D
A
tract (OFT) development (Kioussi et al., 2002). Second, Hjalt et al., (Hjalt et al., 2000), by using antibodies recognizing all PITX2 protein isoforms, detected the presence of PITX2 protein in the right atrial and ventricular wall at stage E12.5 in mouse, contradicting the Pitx2c mRNA expression pattern previously described (Cam-pione et al., 1999; Logan et al., 1998; Ryan et al., 1998; Yoshioka et al., 1998). Third, analysis of the fate of Pitx2 daughter cells in mutant embryos that only express Pitx2a and Pitx2b isoforms (Pitx2 δabccreneo;δc and Rosa26 reporter trans-heterozygotes) (Liu et al., 2002) revealed that fewer lacZ-positive cells were found in the right ventricular and inner curvature myocardium of Pitx2 mutant embryos at both 14.5 dpc and 18.5 dpc. And fourth, comparative anatomical analysis of hearts from Pitx2 specific null mutant mice (Pitx2abc -/- (Gage et al., 1999), Pitx2ab -/- (Liu et al., 2001) and Pitx2c -/- (Liu et al., 2002)) leads to speculate that PITX2A and/or PITX2B could play a role in the development of the OFT (Franco et al., 2003).
Here, we report for the first time that Pitx2a and Pitx2b isoforms are co-expressed with Pitx2c isoform during heart development. In agreement with previous works, our qRT-PCR analysis showed that Pitx2c is largely the most abundant isoform during heart
de-velopment, maintaining low expression levels in the adult heart. Interestingly, in terms of relative quantification of mRNA, while Pitx2a expression is only detected at early stages, Pitx2b displays a similar expression profile to that for Pitx2c, suggesting relevance for Pix2b during cardiogenesis. Since Pitx2a and Pitx2b share the same promoter and the mature Pitx2a and Pitx2b mRNAs are origi-nated from a common RNA molecule that undergoes a process of alternative splicing (AS), we could speculate that posttrancriptional regulation mechanisms could balance the amount of Pitx2a and Pitx2b isoforms. In fact, recent findings indicate that AS is controlled by the presence of post-transcriptional RNA operons or regulons (Keene, 2007). Pitx2a and Pitx2b expression is significantly lower than Pitx2c expression and current qRT-PCR techniques are largely more sensitive than any ISH previously used to analyse Pitx2 isoform expression (Campione et al., 1999; Schweickert et al., 2000). Therefore, it is not surprising that Pitx2a and Pitx2b expression have not been detected before in the developing heart. Although Pitx2, especially Pitx2c, mRNA expression patterns during cardiogenesis are apparently well known (Franco et al., 2003), PITX2 protein distribution in heart development is still poorly understood and seems to be contradictory (Hjalt et al.,
tion in the right ventricle. Yellow square on image B amplified 40X;
(D) PITX2B protein distribution in the interventricular septum. White square on image B amplified 40X;
(E) PITX2B protein distribution in the left ventricle. Blue square on image B amplified 40X. Asterisk indicate trabecular region on each ventricle. (F-I) stage E15.5; (F)
10X transverse section of mouse embryos heart at stage E15.5;
(G) PITX2B protein distribution in the right ventricle. Yellow square on image F amplified 40X; (H)
PITX2B protein distribution in the interventricular septum. White square on image F amplified 40X;
(I) PITX2B protein distribution in the left ventricle. Blue square on image F amplified 40X.
Fig. 4. Quantitative analysis of PITX2B positive cells among different stages during cardiogenesis.(A) Quan-tification of PITX2B positive cells distribution by cardiac chambers. Embryonic day (E) 9.5 to 17.5; N1, 1 day neonate mouse heart. RA: Right atria; LA: Left atria; RV: Right ventricle; LV: Left ventricle; IS: Interventricular septum. **P-value <0.01 and ***P-value <0.001. (B-I) PITX2B protein distribution in ventricular chambers at stages E13.5 and E15.5. Nucleus and PITX2B stain are showed in red and green respectively.
(B-E) stage E13.5; (B) 10X transverse section of mouse embryos heart at stage E13.5; (C) PITX2B protein
distribu-G
B
C
D
E
F
H
I
2000). Here, we report by the firs time the expression pattern of a specific PITX2 protein isoform during heart development. Our results show how PITX2B protein is differentially expressed within cardiac compartments during heart development. Thus, in contrast to Pitx2c (Campione et al., 1999; Schweickert et al., 2000), PITX2B distribution is ubiquitous in the atrial chambers, suggesting that PITX2B is not mainly involved in left-right atrial identity. However, within ventricular compartments, PITX2B protein progressively accumulates in the interventricular septum between E11.5 and E13.5 coinciding with the developmental window in which ven-tricular septation takes place (Miquerol et al., 2012). Liu et al., previously showed the presence of Pitx2-daugthers cells in the interventricular septum and atrioventricular cushions (Liu et al., 2002). As suggested by the authors, one interpretation of these data is that Pitx2c daughter cells can expand to populate other hearts regions. Nevertheless, the contribution of cell populations expressing Pitx2a or Pitx2b isoforms in those cardiac tissues cannot be ruled out. In addition, it has been previously shown that Pitx2abc null mice exhibit ventricular septation defects (Gage et al., 1999) whereas Pitx2c mutants did not (Liu et al., 2002). These data suggest that the isoforms PITX2A and PITX2B could have a redundant function with PITX2C during heart development (Liu et al., 2001). In addition, we cannot rule out the possibility that dif-ferent PITX2 isoforms could be coexpressed in the same cardiac cells, according to the fact that different isoforms of PITX2 can act together forming homo and heterodimers leading to transcriptional synergism (Cox et al., 2002; Saadi et al., 2003).
Current studies suggest that Pitx2-mediated signalling during cardiogenesis is conducted within three different cell types: the
myocardium, the cardiac neural crest (CNC) cells, and the pha-ryngeal arch mesenchyme (Campione et al., 1999; Chinchilla et al., 2011; Franco et al., 2003; Kioussi et al., 2002; Liu et al., 2002; Tessari et al., 2008), minimizing the possible role that Pitx2 a, b or c, may have during development of epicardial and endocardial derivatives. In addition, it was previously reported only that Pitx2c expression is never detected in endocardium (Liu et al., 2002). Notably, we reported here the presence of PITX2B in epicardium and endocardium during heart development, opening new puta-tive roles for this gene during heart organogenesis. Concerning the possible role of Pitx2 in myocardium, Tessari et al., (Tessari et al., 2008) reported how myocardial Pitx2 expression delineates the remodelling of the left atrioventricular canal, contributing to atrio-ventricular septation. Interestingly our results show that in the atrioventricular canal compartment PITX2B protein is mainly present in the mural primordium of the mitral valve during heart development, suggesting a role of PITX2B in the maturation of the right portion of the atrioventricular canal. Although Liu et al., (Liu et al., 2002) speculated that Pitx2 daughter cells from myocardium invade the AV cushions and valves, in mice it has been also proposed that the leaflets and tendinous cords of the tricuspid valve are all generated from mesenchyme, derived from the endocardium with no substantial contribution from cells of the myocardial linage (de Lange et al., 2004). In whatever manner this cells arrives to the AV cushions, we cannot determine the source of cells that participate in this process since we have detected the presence of PITX2B in endocardium, myocardium and epicardium. Finally, we must stress that this PITX2B expression pattern in the atrioventricular canal constitute the only sign of asymmetry for PITX2B distribution
Fig. 5. Quantitative analysis of PITX2B positive cells in the AV canal structures. Quantificationof PITX2B positive cells in the AV canal structures at stage 9.5 (A), stage 11.5 (B) and stage 13.5, (C) Tricuspid valve primor-dium (D) Mitral valve primordium).
AVCC Atrio-ventricular canal cushions.
(E-K) PITX2B protein distribution in the AV canal structures from E9.5 to E13.5. Nucleus and PITX2B stain are showed in red and green respectively.
(E) 10X transverse section of mouse embryos heart at stage E9.5. (F)
White square on image E amplified 40X. (G) 10X transverse section of mouse embryos heart at stage E11.5.
(H) White square on image G ampli-fied 40X. (I) 10X transverse section of mouse embryos heart at stage E13.5. (J) PITX2B protein distribution in Tricuspid Valve primordia. White square on image I amplified 40X. (K)
PITX2B protein distribution in Mitral Valve primordia. Blue square on image I amplified 40X. AVCC: Atrio-ventricular canal cushions; TVP: Tricuspid Valve primordia; MVP: Mitral Valve primordia; MP: Mural portion; SP: Septal portion; RA: Right atria; LA: Left atria; RV: Right ventricle; LV: Left ventricle; IS: Interventricular septum. White arrows indicate PITX2B positive cells. ***P-value <0.001.
G
E
F
H
I
J
K
B
C
D
during cardiogenesis.
In summary, we report that Pitx2a and Pitx2b isoforms are co-expressed with Pitx2c isoform during heart development. Interest-ingly, in terms of relative quantification of mRNA, Pitx2b and Pitx2c isoforms display similar expression profiles during cardiogenesis, decreasing with further development but maintaining its expression until adult stages. Moreover, a detailed analysis of PITX2B protein during cardiac development shows that PITX2B is dynamically expressed in the developing ventricular septum and asymmetri-cally expressed in the tricuspid valves primordia. This suggests a putative role for PITX2B during ventricular septation as well as in the maturation of the right portion of the atrioventricular canal.
Materials and Methods
Tissue preparation
Balb/c female mice were sacrificed and whole hearts ranging from embryonic day (E) 9.5 to E17.5 were isolated. The day of vaginal plug was taken as E0.5. Adult and neonatal hearts (1 and 15 days) were also obtained. Hearts for RNA isolation and immunohistochemistry experiments were processed as previously described (Dominguez et al., 2005)
Quantitative real time PCR (q-PCR)
Total RNA was isolated and reverse transcribed from pooled hearts at each stage (n = 5 per stage), ranging from E9.5 to E17.5 embryos, and from neonate and adult mice (n = 3 per stage) as previously described (Dominguez et al., 2005). Finally, each experimental point is represented by the average of five different pools of hearts. The primers used to detect mouse Pitx2a (NM_001042504.2), Pitx2b (NM_011098.4), Pitx2c (NM_001042502.2) and b-actin (NM_007393) gene expression (Table 1) were specifically designed by using Primer3v.0.4.0 program (available on line http://frodo.wi.mit.edu/).
Real-time PCR was performed within an iCycler PCR thermocycler (Bio-Rad, Spain) and SYBR Green detection system. Reactions were performed in 96-well plates with optical sealing tape (Bio-Rad) in 20 ml total volume containing SYBR Green Mix (Bio-Rad) and cDNA corresponding to 50 ng of total RNA. Mouse b-actin was used in parallel for each run as internal control. Amplification conditions were: 95 °C for 5 min; 45 cycles of 95 °C for 30 s, 60 °C for 30 s, 72 °C for 30 s; and 72 °C for 10 min. The relative expression level of each Pitx2 isoform was calculated through Livak analysis method (Livak, Schmittgen, 2001).
Immunohistochemistry
Serial tissue sections from littermates were cut for parallel analysis. The distribution of PITX2B and DESMIN proteins were analysed by im-munohistochemistry and confocal microscopy as previously described (Dominguez et al., 2005). Rabbit polyclonal antibody against mouse PITX2B (Hernández-Torres et al., 2008) and mouse monoclonal antibody against mouse DESMIN (Sigma, Spain) were incubated overnight at room temperature with a 1:50 dilution. Following rinsing, sections were incubated
for 5 h with antirabbit Cy3 or antimouse Cy2 secondary antibodies (Jackson Labs, USA) diluted in TBSA-BSAT at 1:200 or 1:100 respectively. Nuclear staining was performed using DRAQ-5™ (Red Fluorescent Cell-Permeable DNA probe, Biostatus Limited).
Quantification of nucleus (total and PITX2B positive) was done as Car-mona et al., (CarCar-mona et al., 2007) indicated. This method renders images devoid of both, extranuclear staining and background, thus emphasizing the nuclear signal. Images used in this work correspond to frontal heart sections in witch Left Atria, Right Atria, Interventricular Septum (when it was present), Left Ventricle and Right Ventricle structures were clearly recognizable. Each structure was isolated as an individual image for its individual analisys; the total number of nucleus as well as the number of PITX2B positive nucleus were measured. Finally, the proportion of PITX2B positive nucleus was calculated by dividing the number of PITX2B posi-tive nucleus by the total number of nucleus counted on each image. This ratio was finally expressed as a percentage. Each experimental point is represented by the average of the analysis of five different heart images for each embryo with a total number of three embryos at each stage.
Acknowledgements
We thank Chris Doherty for reading the manuscript and the “Centro de Intrumentación Científico-Técnico (CICT), Universidad de Jaén” for the confocal microscopy help. This work was supported by grants of Span-ish Ministry of Economy and Competitiveness, MINECO, and FEDER (BFU2013-48643-C3-2-P to F.N.) and Junta de Andalucía (BIO258, PI10-CVI6521 to F.N.).
References
ADAMS R, PORRAS A, ALONSO G, JONES M, VINTERSTEN K (2000). Essential role of p38a MAP kinase in placental but not embryonic cardiovascular
develop-ment. Mol Cell. 6:109-116
CAMPIONE M, ROS MA, ICARDO JM, PIEDRA E, CHRISTOFFELS VM, SCH-WEICKERT A, BLUM M, FRANCO D, MOORMAN AF (2001). Pitx2 expression defines a left cardiac lineage of cells: evidence for atrial and ventricular molecular isomerism in the iv/iv mice. Dev Biol 231: 252–264.
CAMPIONE M, STEINBEISSER H, SCHWEICKERT A, DEISSLER K, VAN BEBBER F, LOWE LA, NOWOTSCHIN S, VIEBAHN C, HAFFTER P, KUEHN MR, BLUM M (1999). The homeobox gene Pitx2: mediator of asymmetric left-right signaling in vertebrate heart and gut looping. Development 126: 1225–1234.
CARMONA R, MACÍAS D, GUADIX JA, PORTILLO V, PÉREZ-POMARES JM, MUÑOZ-CHÁPULI R (2007). A simple technique of image analysis for specific nuclear immunolocalization of proteins. J Microsc 225: 96–99.
CHINCHILLA A, DAIMI H, LOZANO-VELASCO E, DOMINGUEZ JN, CABALLERO R, DELPÓN E, TAMARGO J, CINCA J, HOVE-MADSEN L, ARÁNEGA AE, FRANCO D (2011). PITX2 insufficiency leads to atrial electrical and structural remodeling linked to arrhythmogenesis. Circ Cardiovasc Genet 4: 269–279.
CHINCHILLA A, LOZANO E, DAIMI H, ESTEBAN FJ, CRIST C, ARANEGA AE, FRANCO D (2011). MicroRNA profiling during mouse ventricular maturation: a role for miR-27 modulating Mef2c expression. Cardiovasc Res 89: 98–108. COX CJ, ESPINOZA HM, MCWILLIAMS B, CHAPPELL K, MORTON L, HJALT TA,
SEMINA EV, AMENDT BA (2002). Differential regulation of gene expression by PITX2 isoforms. J Biol Chem 277: 25001–25010.
CUNNINGHAM ET, ELIOTT D, MILLER NR, MAUMENEE IH, GREEN WR (1998). Familial Axenfeld-Rieger anomaly, atrial septal defect, and sensorineural hearing loss: a possible new genetic syndrome. Arch Ophthalmol 116: 78–82. DE LANGE FJ, MOORMAN AFM, ANDERSON RH, MÄNNER J, SOUFAN AT, DE
GIER-DE VRIES C, SCHNEIDER MD, WEBB S, VAN DEN HOFF MJB, CHRIST-OFFELS VM (2004). Lineage and morphogenetic analysis of the cardiac valves.
Circ Res 95: 645–654.
DOMINGUEZ JN, NAVARRO F, FRANCO D, THOMPSON RP, ARÁNEGA AE (2005). Temporal and spatial expression pattern of beta1 sodium channel subunit during heart development. Cardiovasc Res 65: 842–850.
FRANCO D, CAMPIONE M (2003). The role of Pitx2 during cardiac development. Linking left-right signaling and congenital heart diseases. Trends Cardiovasc
Oligonucleotides Sequence Amplicon Size (bp)
β-actin F
β-actin R 5´-CCAGAGGCATACAGGGAC-3´ 5´-TGAGGAGCACCCTGTGCT-3´ 144 Pitx2a F
Pitx2a R 5´ GAGAGCAGCAGACAGAAAC 3´ 5´ ATCTTTCTCTAATTGCACGC 3' 270 Pitx2b F
Pitx2b R 5' GGTGCAGTTCACGGACTCTC 3' 5' TGTCTGGGTAGCGGTTTCTC 3' 233 Pitx2c F
Pitx2c R 5' CCTCACCCTTCTGTCACCAT 3' 5' GCCCACATCCTCATTCTTTC 3' 179 TABLE 1
HERNÁNDEZ-TORRES F, PEDRAJAS JR, ARÁNEGA AE, NAVARRO F (2008). Expression in bacteria of small and specific protein domains of two transcription factor isoforms, purification and monospecific polyclonal antibodies generation, by a two-step affinity chromatography procedure. Protein Expr Purif 60: 151–156. HJALT TA, SEMINA EV, AMENDT BA, MURRAY JC (2000). The Pitx2 protein in
mouse development. Dev Dyn 218: 195–200.
KEENE JD (2007). RNA regulons: coordination of post-transcriptional events. Nat
Rev Genet 8: 533–543.
KIOUSSI C, BRIATA P, BAEK SH, ROSE DW, HAMBLET NS, HERMAN T, OHGI KA, LIN C, GLEIBERMAN A, WANG J, BRAULT V, RUIZ-LOZANO P, NGUYEN HD, KEMLER R, GLASS CK, WYNSHAW-BORIS A, ROSENFELD MG (2002). Identification of a Wnt/Dvl/beta-Catenin --> Pitx2 pathway mediating cell-type-specific proliferation during development. Cell 111: 673–685.
KIRCHHOF P, KAHR PC, KAESE S, PICCINI I, VOKSHI I, SCHELD H-H, ROTERING H, FORTMUELLER L, LAAKMANN S, VERHEULE S, SCHOTTEN U, FABRITZ L, BROWN NA (2011). PITX2c is expressed in the adult left atrium, and reducing Pitx2c expression promotes atrial fibrillation inducibility and complex changes in gene expression. Circ Cardiovasc Genet 4: 123–133.
KITAMURA K, MIURA H, MIYAGAWA-TOMITA S, YANAZAWA M, KATOH-FUKUI Y, SUZUKI R, OHUCHI H, SUEHIRO A, MOTEGI Y, NAKAHARA Y, KONDO S, YOKOYAMA M (1999). Mouse Pitx2 deficiency leads to anomalies of the ventral body wall, heart, extra- and periocular mesoderm and right pulmonary isomerism.
Development 126: 5749–5758.
LINCOLN J, ALFIERI CM, YUTZEY KE (2004). Development of heart valve leaflets and supporting apparatus in chicken and mouse embryos. Dev Dyn 230: 239–250. LIU C, LIU W, LU MF, BROWN NA, MARTIN JF (2001). Regulation of left-right
asym-metry by thresholds of Pitx2c activity. Development 128: 2039–2048.
LIU C, LIU W, PALIE J, LU MF, BROWN NA, MARTIN JF (2002). Pitx2c patterns anterior myocardium and aortic arch vessels and is required for local cell move-ment into atrioventricular cushions. Developmove-ment 129: 5081–5091.
LIVAK K, SCHMITTGEN T (2001). Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 25: 402–402408. LOGAN M, PAGÁN-WESTPHAL SM, SMITH DM, PAGANESSI L, TABIN CJ (1998).
anomaly in Rieger Syndrome: heterogeneity or contiguity? Acta Ophthalmol
Scand 76: 509–512.
MIQUEROL L, KELLY RG (2012). Organogenesis of the vertebrate heart. WIREs
Dev Biol 2: 17–29.
RYAN AK, BLUMBERG B, RODRIGUEZ-ESTEBAN C, YONEI-TAMURA S, TAMURA K, TSUKUI T, LA PEÑA DE J, SABBAGH W, GREENWALD J, CHOE S, NORRIS DP, ROBERTSON EJ, EVANS RM, ROSENFELD MG, IZPISÚA BELMONTE JC (1998). Pitx2 determines left-right asymmetry of internal organs in vertebrates.
Nature 394: 545–551.
SAADI I, KUBURAS A, ENGLE JJ, RUSSO AF (2003). Dominant negative dimeriza-tion of a mutant homeodomain protein in Axenfeld-Rieger syndrome. Mol Cell
Biol 23: 1968–1982.
SCHWEICKERT A, CAMPIONE1 M, STEINBEISSER H, BLUM M (2000). Pitx2 isoforms: involvement of Pitx2c but not Pitx2a or Pitx2b in vertebrate left-right asymmetry. Mech Dev 90: 41–51.
SEMINA EV, REITER R, LEYSENS NJ, ALWARD WL, SMALL KW, DATSON NA, SIEGEL-BARTELT J, BIERKE-NELSON D, BITOUN P, ZABEL BU, CAREY JC, MURRAY JC (1996). Cloning and characterization of a novel bicoid-related homeobox transcription factor gene, RIEG, involved in Rieger syndrome. Nat
Genet 14: 392–399.
TESSARI A, PIETROBON M, NOTTE A, CIFELLI G, GAGE PJ, SCHNEIDER MD, LEMBO G, CAMPIONE M (2008). Myocardial Pitx2 differentially regulates the left atrial identity and ventricular asymmetric remodeling programs. Circ Res 102: 813–822.
TSAI JC, GRAJEWSKI AL (1994). Cardiac valvular disease and Axenfeld-Rieger syndrome. Am J Ophthalmol 118: 255–256.
WANG J, KLYSIK E, SOOD S, JOHNSON RL, WEHRENS XHT, MARTIN JF (2010). Pitx2 prevents susceptibility to atrial arrhythmias by inhibiting left-sided pacemaker specification. Proc Natl Acad Sci USA 107: 9753–9758.
Genome-wide gene expression analysis in mouse embryonic stem cells Juan Sainz, Fernando García-Alcalde, Armando Blanco and Ángel Concha Int. J. Dev. Biol. (2011) 55: 995-1006
http://dx.doi.org/10.1387/ijdb.103123js
Building the vertebrate heart - an evolutionary approach to cardiac development
5 yr ISI Impact Factor (2013) = 2.879 José M. Pérez-Pomares, Juan M. González-Rosa and Ramón Muñoz-Chápuli
Int. J. Dev. Biol. (2009) 53: 1427-1443 http://dx.doi.org/10.1387/ijdb.072409jp
Heart formation and left-right asymmetry in separated right and left embryos of a newt Kazuhiro Takano, Yuzuru Ito, Shuichi Obata, Tsutomu Oinuma, Shinji Komazaki, Hiroaki Nakamura and Makoto Asashima
Int. J. Dev. Biol. (2007) 51: 265-272 http://dx.doi.org/10.1387/ijdb.072270kt
Xenopus nodal related-1 is indispensable only for left-right axis determination Ryuji Toyoizumi, Tsuyoshi Ogasawara, Shigeo Takeuchi and Kazue Mogi
Int. J. Dev. Biol. (2005) 49: 923-938 http://dx.doi.org/10.1387/ijdb.052008rt
A P19Cl6 GFP reporter line to quantify cardiomyocyte differentiation of stem cells Jennifer C Moore, Rene Spijker, Anton C Martens, Teun de Boer, Martin B Rook, Marcel A G van der Heyden, Leon G Tertoolen and Christine L Mummery