Copyright2000 by the Genetics Society of America
A Screen for Dominant Modifiers of
ro
Dom, a Mutation That Disrupts
Morphogenetic Furrow Progression in Drosophila, Identifies Groucho
and Hairless as Regulators of
atonal
Expression
Franc¸oise Chanut,* Alvin Luk
†,1and Ulrike Heberlein*
,‡*Department of Anatomy,†Gallo Center and‡Program in Neuroscience and Developmental Biology, University of California, San Francisco, California 94143
Manuscript received May 3, 2000 Accepted for publication July 24, 2000
ABSTRACT
roDom is a dominant allele of rough (ro) that results in reduced eye size due to premature arrest in
morphogenetic furrow (MF) progression. We found that theroDomstop-furrow phenotype was sensitive to
the dosage of genes known to affect retinal differentiation, in particular members of thehedgehog(hh) signaling cascade. We demonstrate that roDom interferes with Hh’s ability to induce the retina-specific
proneural geneatonal(ato) in the MF and that normal eye size can be restored by providing excess Ato protein. We usedroDomas a sensitive genetic background in which to identify mutations that affecthhsignal
transduction or regulation ofatoexpression. In addition to mutations in several unknown loci, we recovered multiple alleles ofgroucho(gro) andHairless(H). Analysis of their phenotypes in somatic clones suggests that both normally act to restrict neuronal cell fate in the retina, although they control different aspects ofato’s complex expression pattern.
M
ORPHOGENESIS in higher animals requires the dentation, known as the morphogenetic furrow (MF), coordination of cell movements or cell shape in the retinal epithelium (Readyet al.1976;Tomlinsonchanges with the implementation of novel programs of 1985). In the MF, differentiation begins with the expres-gene expression (SmithandSchoenwolf1998;Leptin sion of the proneural gene atonal(ato), a member of 1999;NoselliandAgne`s1999). Retinal differentiation the basic-helix-loop-helix (b-HLH) gene family with high in Drosophila provides a striking example of an intricate similarity to members of theAchaete-Scutecomplex (
Jar-pattern of diverse cell fates that is achieved in a short manet al.1993, 1994).atofunction is required only for time over a large field of cells with minimum movement the differentiation of one of the eight photoreceptors or waste of cells and with remarkable accuracy. The that compose a mature ommatidium, photoreceptor R8 precision and rapidity of the process result from a com- (Jarmanet al. 1994). However, the R8 precursor acts plex network of long-range and short-range signaling as a founder cell that induces the progressive recruit-events that impose tight temporal and spatial regulation ment of all other ommatidial cell types (Tomlinson
to the differentiation process (Heberleinand Moses and Ready 1987). As a consequence, ato function is 1995;Albagli et al.1997;Freeman 1997). indispensable for retina formation, and adults homozy-Retinal differentiation in Drosophila begins during gous for loss-of-function ato alleles have very reduced the third larval instar, when a few cells near the center eyes devoid of photoreceptors (Jarmanet al.1994). of the posterior eye imaginal disc margin start clustering Progress of the MF is intimately linked to cross-regula-and expressing neuron-specific markers (Wolff and tion betweenatoand the segment polarity genehedgehog
Ready 1993). The anterior portion of the disc keeps (hh). Hh is produced by cells that differentiate, as an growing through random cell proliferation, while differ- indirect consequence ofatoexpression, behind the MF entiation sweeps across the disc as a wave, spreading (Heberleinet al.1993;Maet al.1993). When Hh signal-evenly to the lateral margins and eventually reaching ing is reduced during the third larval instar, using a the anterior margin over the course of 2 days. At the temperature-sensitivehhallele, or in somatic clones de-front of the wave, cells stop dividing and undergo con- void of the putative Hh receptor Smoothened (Smo; certed apical constrictions, which creates a narrow in- Alcedo et al.1996), atoexpression is abolished in MF cells (Domi´nguezandHafen1997;Struttand Mlod-zik1997;BorodandHeberlein1998). This is
accom-Corresponding author:Franc¸oise Chanut, Department of Anatomy, panied by the premature arrest or severe retardation of
S-1334, Box 0452, University of California, 513 Parnassus Ave., San
MF progression (Heberleinet al.1993;Maet al.1993;
Francisco, CA 94143. E-mail: [email protected]
Domi´nguez and Hafen 1997; Strutt and Mlodzik
1Present address:Tularik, Inc., 2 Corporate Dr., S. San Francisco, CA
94080. 1997;BorodandHeberlein1998). The anterior
ment of the MF therefore relies on the cells’ transition of the Rough protein in eye differentiation (Heberlein
from a state where they respond to the Hh signal and et al. 1991, 1993). Unlike loss-of-function ro alleles, expressatoto a state where they emit the Hh signal as which cause roughening of the eye surface without af-a consequence of differentiaf-ating. However, differentiaf-a- fecting eye size (Tomlinsonet al. 1988), roDomleads to
tion is not completely abolished within smo mutant reduced eye size due to premature arrest of MF progres-clones, indicating that Hh is not the sole activator of sion. We foundroDomto be sensitive to dosage manipula-atoexpression in the MF (Domı´nguezandHafen1997; tions of several genes involved in eye development. We
StruttandMlodzik1997). Rather, Hh may be viewed therefore used it as a sensitive background in which to as a permissive factor that facilitates the progress of identify new regulators of MF movement on the basis retinal differentiation by increasing the rate of Ato accu- of their ability to act as dominant modifiers of the
stop-mulation. furrow phenotype.
Like other proneural genes of the Drosophila periph- Some of the strongest genetic interactions were ob-eral nervous system,atofollows a dynamic transcription served with members of the hh pathway, includinghh
pattern. Its expression is first continuous and weak itself and its antagonist patched (ptc). Our screen also ahead of the MF, then increases rapidly and unevenly in yielded new mutations in genes of theNsignaling path-the MF, giving rise to an alternating pattern of strongly way, such asDl,groucho(gro), and Hairless(H), as well expressing and less strongly expressing cell clusters as uncovered theeyelid(eld) locus (Treismanet al.1997). (Jarmanet al.1995;Dokucuet al.1996;Sunet al.1998). Phenotypic characterization ofroDomshows that the
dif-At the posterior edge of the MF, groups of six to eight ferentiation arrest results primarily from decreasedato cells with high Ato protein levels progressively resolve expression. We demonstrate thatgroandHare normal into single, evenly spaced cells that maintain high levels regulators ofatoexpression in the MF.grorepressesato of Ato protein for an additional 6–8 hr and go on to expression behind the MF whereasHrepresses expres-differentiate as R8 photoreceptors (Jarmanet al.1995; sion anterior to the MF. Their role as suppressors of
Bakeret al. 1996). In addition tohh, signals from the roDomis discussed in the context of their relationship to Notch(N) andEpidermal growth factor receptor(EGFR) path- theN pathway.
ways have been implicated in the early stages of ato
induction (BakerandYu1997;Domi´nguezandHafen
1997;Domı´nguezet al.1998;Spenceret al.1998;Sun MATERIALS AND METHODS
et al.1998). atowas also shown to activate its own
tran-Mutagenesis and mutant recovery:Males from an isogenic
scription (Sun et al. 1998). While the exact sequence
w⫺strain were fed ethyl methanesulfonate (EMS) at a
concen-in which these activation mechanisms act has not been
tration of 25–35 mmin 10% sucrose or exposed toⵑ4000 rad
fully established, together they account for the rapid of X-ray irradiation. They were mated to virgin females of the accumulation and early patterning of Ato protein in the T(2;3) roDom/TM3,Sbgenotype. The strategy to recover mutants
MF. Behind the MF, downregulation of atowithin the is illustrated in Figure 1. TheroDomtranslocation acts as a double
balancer for second and third chromosomes. Hence the
mod-intermediate clusters is attributed mainly to lateral
inhi-ifiers can be followed easily by their ability to segregate away
bition mediated by the receptor Notch (N) and its
li-from theroDom-bearing chromosome.
gand Delta (Dl;BakerandZitron1995), in
combina-Fly stocks and culture:Fly stocks are listed in Table 1. The
tion with the adhesion molecule Scabrous (Sca;Leeet roDomphenotype is more pronounced at 25⬚than at 18⬚. The al. 1996). Spacing of the R8 precursors may further screen and all subsequent crosses were performed at 25⬚ in
noncrowded conditions on standard fly medium.
depend on the patterned activation of the EGFRand/
Induction of somatic clones:Somatic clones were induced
orRas/Rafsignaling pathways (Domı´nguezet al.1998;
using the yeast Flip recombinase (FLP) and target sequence Kumaret al.1998;Spenceret al.1998;Greenwoodand
(FRT) system (GolicandLindquist1989;Golic1991).gro347,
Struhl1999). The homeodomain protein Rough (Ro; H299, andHE31mutations were recombined onto a third
chro-Tomlinsonet al.1988;Heberleinet al.1991), an inhibi- mosome carrying an FRT at position 82B (Xu and Rubin tor ofatotranscription expressed at the posterior edge 1993). To obtain marked clones in adults, the following crosses were performed:y, w, hs-FLP122;FRT(82B)P[w⫹]90Efemales⫻
of the MF, ensures the individualization of single ato
-w/Y;FRT(82B),Horgro/TM3,Sbmales. Progeny were grown
expressing cells from an equivalence group including
at 25⬚ and submitted to a 1-hr heat shock at 38.5⬚, once at
the R8, R2, and R5 photoreceptor precursors (Dokucu
the end of first larval instar (48 hr after egg laying) and once
et al.1996). again at the end of second larval instar (72 hr after egg laying). Given the variety of signals that converge in the MF, The presence of mutant clones was inferred from unpig-a genetic screen bunpig-ased on the disruption of furrow move- mented patches of retinal tissue in an otherwise normal,w⫹
adult eye.
ment should provide insights into many interesting
pat-To obtain marked clones in larvae, the following crosses
terning and differentiation processes. Here we report
were performed:y, w, hs-FLP122;FRT(82B),arm-lacZfemales⫻
the results of a genetic screen we undertook to identify
w/Y;FRT(82B),Horgro/TM3,Sbmales. The same heat-shock
new genes that promote or prevent progress of the MF. regimen as above was used to induce somatic recombination. We used a dominantro allele,roDom, that had been
iso-Thearm-lacZreporter (Vincentet al.1994) expresses -galacto-sidase ubiquitously in larval tissues. The presence of mutant
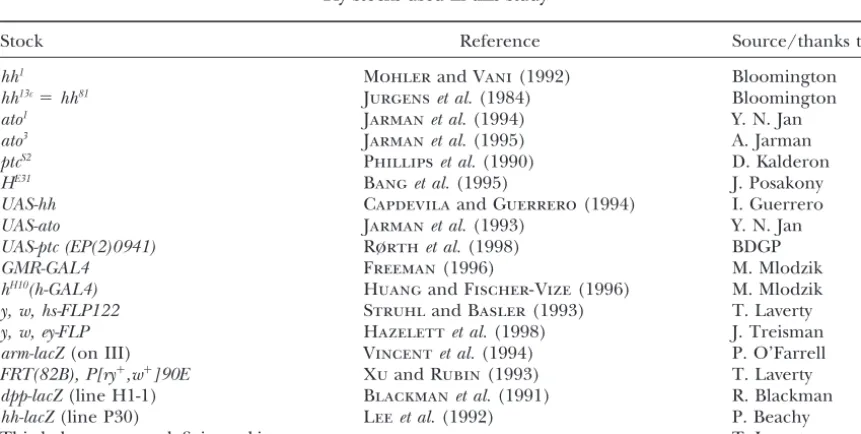
Figure 1.—A schematic outline of the genetic screen used to isolate modifiers ofroDomon chromosomes
II and III. M, modifier.
clones was inferred from the loss of-galactosidase activity in Immunohistochemistry and microscopy:Primary antibodies were kindly provided by the following people and were used patches of eye-disc cells.
The frequency of eye clones was greatly increased when we at the following dilutions: Rabbit-anti-Atonal (Andrew Jarman; preabsorbed, 1:1000 final); Mouse-anti-Rough (Gerry Rubin; used a construct that places the FLP recombinase under the
control of an eye-specific enhancer element from theeyeless 1:100); Rat-anti-ELAV (Gerry Rubin; 1:5), Mouse-anti-Hairy (Nadean Brown; 1:5); Mouse-anti-Boss (Larry Zipursky; 1:5). gene, designed by B. Dickson (ey-FLP;Hazelettet al.1998).
Crosses were performed as described above, replacinghs-FLP Dissection, fixation, antibody detection, and-galactosidase activity staining of eye-imaginal discs were performed as pre-withey-FLP, also on the X chromosome. No heat induction
was necessary asey-FLPis expressed constitutively in the eye viously described (Kimmelet al.1990;Treismanet al.1995). Confocal images were obtained on a Bio-Rad (Hercules, CA) disc from second instar on.
TABLE 1 Fly stocks used in this study
Stock Reference Source/thanks to
hh1 MohlerandVani(1992) Bloomington
hh13c⫽hh81 Jurgenset al.(1984) Bloomington
ato1 Jarmanet al.(1994) Y. N. Jan
ato3 Jarmanet al.(1995) A. Jarman
ptcS2 Phillipset al.(1990) D. Kalderon
HE31 Banget al.(1995) J. Posakony
UAS-hh CapdevilaandGuerrero(1994) I. Guerrero
UAS-ato Jarmanet al.(1993) Y. N. Jan
UAS-ptc (EP(2)0941) Rørthet al.(1998) BDGP
GMR-GAL4 Freeman(1996) M. Mlodzik
hH10(h-GAL4) HuangandFischer-Vize(1996) M. Mlodzik
y, w, hs-FLP122 StruhlandBasler(1993) T. Laverty
y, w, ey-FLP Hazelettet al.(1998) J. Treisman
arm-lacZ(on III) Vincentet al.(1994) P. O’Farrell
FRT(82B), P[ry⫹,w⫹]90E XuandRubin(1993) T. Laverty
dpp-lacZ(line H1-1) Blackmanet al.(1991) R. Blackman
hh-lacZ(line P30) Leeet al.(1992) P. Beachy
Figure2.—(A–F) The stop-furrow phenotype inroDomand hh1(see alsoHeberleinet al. 1993;
ChanutandHeberlein1997). In all panels, pos-terior is to the right. (A–C) Scanning electron microscopy (SEM) images of the left eye of
roDom/⫹ (A), wild-type (B), andhh1homozygous
(C) adult flies. Ommatidia are restricted to the posterior eye portion of roDom and hh1 mutants.
(D–F) Immunohistochemical staining of third in-star eye discs with an antibody against ELAV. In wild type (E), immature clusters (black arrow) form at the anterior front of differentiation and progressively grow into mature ommatidia (white arrow). InroDom/⫹(D) andhh1(F) discs, mature
clusters (white arrows) are found all the way to the front of the differentiation wave, indicating that MF progression has stopped prematurely. (G–I)roDomis sensitive torodosage. Light
micros-copy images of the left eye of a homozygote (roDom/ roDom; G), heterozygote (roDom/⫹; H), and
hemizy-gote (roDom/roX63; I) female. ( J–L)roexpression as
detected with an antibody directed against the Ro protein in wild-type ( J) androDommutant discs (K
and L). ( J) In wild type, Ro is found in most cells along the posterior edge of the MF (white arrowhead) and in a subset of photoreceptors in developing ommatidia (Kimmelet al.1990). (K) In early third instarroDom/⫹mutant discs, Ro
ex-pression is similar to wild type, but in older discs (L), Ro disappears from the front of differentia-tion (arrowhead) as a consequence of furrow ar-rest and cell maturation. Although quantitative comparisons are difficult from one disc to the other, Ro expression in developing clusters consis-tently appeared slightly stronger in mutant vs.
wild type.
MRC 1024 microscope and Nomarsky images on a Zeiss eyes made of fewer than 100 ommatidia, compared to
(Thornwood, NY) Axioskop II. 700–800 in wild type (Figure 2, A and B). Ommatidia
Adult heads were embedded in Durcupan resin (Fluka,
display essentially normal organization and structure
Milwaukee, WI) for sectioning as previously described
(Tom-(not shown), but are confined to the posterior third of
linson and Ready 1987). Two-micrometer retinal sections
were mounted in DPX mountant (Fluka) and observed by the eye, while the anterior part of the eye is occupied
phase microscopy on a Zeiss AxioskopII. Images were captured by a few unpatterned pigment cells and by head cuticle. with a Spot2 (Technical Instruments, San Francisco) camera
As previously shown (Heberlein et al. 1993), the
re-and processed using Photoshop software. Scanning electron
duced eye size inroDomresults from the premature arrest
microscopy of adult heads was performed as previously
de-scribed (Kimmelet al.1990). of retinal differentiation during the third larval instar.
roDom mutant discs stained to reveal expression of
neu-ronal markers contain mature photoreceptor clusters
RESULTS
all the way to the anterior front of differentiation (Fig-ure 2D), instead of the smooth gradient of maturing
roDomis a gain-of-function stop-furrow mutation:Flies
2E). Ommatidial clusters in the posterior disc region alleles ofhedgehog, eyelid, Delta, groucho, andHairless: roDom heterozygous females were crossed to wild-type
therefore finish their maturation, but the induction of
new clusters at the front edge is interrupted: this is the males that had been mutagenized with EMS or X ray (see materials and methods). F1 individuals whose
hallmark of a stop-furrow mutation.
roDom is an X-ray-induced translocation of the tip of eyes were larger or smaller thanroDomheterozygotes were
crossed to balancer stocks. Interactions withroDomwere
the right arm of chromosome III onto the base of the
left arm of chromosome II. One of these breakpoints retested in their progeny, and the modifier mutations allocated to the second or third chromosome (Figure is close to therolocus (97D5) and molecular mapping
revealed a genomic rearrangement 5⬘to theroughgene, 1). Stocks were established with the appropriate bal-ancer chromosomes and assessed for lethality or visible although the coding region appeared intact (U.
Heberlein,unpublished observations). Several obser- homozygous phenotypes. Out of 130,000 F1 progeny
scored, we recovered 103 modifiers, including 53 en-vations suggest that the stop-furrow phenotype results
from a gain-of-function mutation of the rough locus. hancers and 50 suppressors (Table 2).
To address the specificity of the modifiers’ effect on First, a differentiation defect very similar toroDomis
ob-served upon overexpressing therogene during the third roDom, we took advantage of a novel mutation recovered
in our screen. This mutation,2033, displays a dominant larval instar under the control of a heat-shock promoter
(Kimmelet al.1990). In contrast, mutations that reduce stop-furrow phenotype very similar to roDomandhh1,
al-though it maps to a distinct location (K. WooandU.
or abolishro function are recessive and do not lead to
reduced eye size. Instead, they cause roughening of the Heberlein,unpublished results). A large subset ofroDom
modifiers was found to modify2033as well. These muta-eye, due to photoreceptor mis-specification (
Tomlin-son et al. 1988; Heberlein et al. 1991). Second, the tions might therefore represent genes with a general role in furrow progression, rather than genes that inter-dominantroDomstop-furrow phenotype proved reversible
by EMS mutagenesis and all revertants failed to comple- act specifically withroor theroDomallele.
Although most modifier stocks carried a lethal muta-ment loss-of-functionroalleles (not shown). Third,
re-moving the wild-type copy ofro in aroDommutant back- tion, only 15 lethal complementation groups were
found, representing a total of 42 independent muta-ground partially suppressed the reduced eye phenotype
(compare Figure 2, H and I), whereas two copies of tions. This indicates that our mutagenesis was not satu-rating. Most single hits were not characterized further,
roDomled to strong enhancement (Figure 2G). Eye-disc
antibody staining detected the Ro protein in the same unless their interaction withroDomwas particularly strong.
Complementation groups were mapped by meiotic re-cells in wild type androDom, though at slightly elevated
levels in the mutant (Figure 2, J–L). On the basis of combination or by lethal complementation against a third chromosome deficiency collection [Berkeley Dro-these genetic and molecular criteria, we believe that the
roDommutation leads to a slight overexpression ofro. sophila Genome Project (BDGP); Table 1]. Allelism to
known loci was tested, and we identified four new alleles Loss-of-function mutations inhh, such as the
eye-spe-cific allele hh1, display a stop-furrow phenotype very ofDland one ofglass(gla) that acted as strong enhancers
ofroDomand four new alleles ofH, six ofgro, and two of
similar to roDom (Chanut and Heberlein 1997). This
suggested thatroDomwould provide a good genetic back- hhthat acted as strong suppressors. One of the novel
loci identified in this screen iseyelid (eld;Treisman et
ground in which to isolate new mutations in the hh
signaling cascade. Genetic interactions were systemati- al.1997), which is allelic to thetrithoraxgroup geneosa
and encodes a nuclear protein that affects transcription cally tested betweenroDomand known regulators of eye
development and furrow movement. Strong interac- by participating in a chromatin remodeling complex that also includesbrahma(Collinset al.1999;Va´zquez
tions were indeed observed with various members of thehhsignaling pathway (see below). In addition,roDom
et al. 1999). Among the other novel mutations, some modified weakly the wing phenotype of the N gain-proved highly sensitive to gene dosage alterations in
the Notch and EGFR signaling pathways (not shown). of-function alleleAbruptex(Ax) and ofHheterozygotes, suggesting an involvement in theN signaling pathway Interactions with mutations of the cell-cycle regulator
string, members of the winglesssignaling pathway, and (not shown). For most of them, however, the only de-tectable interactions were withroDomand2033.Examples
members of the retinal fate specification network were
also detected, although they were more modest. No of strong suppressors of roDom recovered in our screen
are shown in Figure 6. They include onegro and one interaction with mutants of the dpp signaling cascade
was observed. Hallele, mutant 519, a viable EMS-induced mutation on 3R, and mutant1480, a member of a lethal comple-Because roDom was sensitive to mutations in several
pathways regulating eye differentiation, a screen for mentation group also mapped to 3R.
The hh paradox: Among our strongest suppressors dominant modifiers of the stop-furrow phenotype was
carried out with the aim of isolating novel members of were two new alleles of hh(Table 2). The direction of this interaction was perplexing: sincehhis required for these pathways or novel regulators of furrow progression.
TABLE 2 Modifiers ofroDom
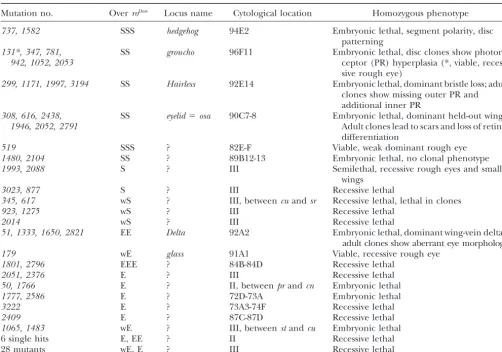
Mutation no. OverroDom Locus name Cytological location Homozygous phenotype
737, 1582 SSS hedgehog 94E2 Embryonic lethal, segment polarity, disc patterning
131*, 347, 781, SS groucho 96F11 Embryonic lethal, disc clones show
photore-942, 1052, 2053 ceptor (PR) hyperplasia (*, viable,
reces-sive rough eye)
299, 1171, 1997, 3194 SS Hairless 92E14 Embryonic lethal, dominant bristle loss; adult clones show missing outer PR and additional inner PR
308, 616, 2438, SS eyelid⫽osa 90C7-8 Embryonic lethal, dominant held-out wings.
1946, 2052, 2791 Adult clones lead to scars and loss of retinal
differentiation
519 SSS ? 82E-F Viable, weak dominant rough eye
1480, 2104 SS ? 89B12-13 Embryonic lethal, no clonal phenotype
1993, 2088 S ? III Semilethal, recessive rough eyes and small
wings
3023, 877 S ? III Recessive lethal
345, 617 wS ? III, betweencuandsr Recessive lethal, lethal in clones
923, 1275 wS ? III Recessive lethal
2014 wS ? III Recessive lethal
51, 1333, 1650, 2821 EE Delta 92A2 Embryonic lethal, dominant wing-vein deltas, adult clones show aberrant eye morphology
179 wE glass 91A1 Viable, recessive rough eye
1801, 2796 EEE ? 84B-84D Recessive lethal
2051, 2376 E ? III Recessive lethal
50, 1766 E ? II, betweenprandcn Embryonic lethal
1777, 2586 E ? 72D-73A Embryonic lethal
3222 E ? 73A3-74F Recessive lethal
2409 E ? 87C-87D Recessive lethal
1065, 1483 wE ? III, betweenstandcu Embryonic lethal
6 single hits E, EE ? II Recessive lethal
28 mutants wE, E ? III Recessive lethal
Only a subset of the 103 modifiers originally recovered is represented in the table, including new alleles of known loci, members of novel lethal complementation groups (more than one hit), and single hits displaying strong interaction withroDomand/or with 2033, an unrelated stop-furrow mutation (K. WooandU. Heberlein,unpublished results). Novel loci were mapped based on their lethal phenotype.eld(discovered in this screen;Treismanet al.1997) and1480were mapped by lethal complementation against a collection of lethalP-element insertions (obtained from Todd Laverty and the BDGP). Other lethals were mapped by meiotic recombination using visible markers or by lethal complementation against the third chromosome deficiency kit (see materials and methods).519is a viable, strong suppressor ofroDom.The suppression was mapped by meiotic recombination
relative to a series ofP-element insertions on 3R (obtained from Todd Laverty). S, suppressor; E, enhancer; w, weak.
we had expectedhhmutations to enhance theroDomphe- duction ofsmo had no effect (not shown), but several
alleles ofCubitus interruptus(Ci), which encodes a tran-notype. Nevertheless, strong suppression was observed
with allhhloss-of-function alleles tested, including our scription factor that modulates Hh signaling (Orenic
et al.1990), acted as strong enhancers (Table 2). AsCi
two new alleles (presumed hypomorphs or nulls, on the
basis of their recessive lethality; Table 2), the eye-specific gives rise to several protein products that can act as mediators or repressors of Hh signaling (Aza-Blancet
hypomorph hh1 and the presumed null hh13c (Figure
3C). Interestingly, loss-of-function mutations inpatched al. 1997; Hepker et al. 1997), the direction of these interactions is difficult to interpret.
(ptc) led to strong enhancement of the stop-furrow
phe-notype (Figure 3D).ptcencodes an integral transmem- roDomprevents expression ofhhtarget genes in the MF:
To understand better how roDom interferes with furrow
brane protein that prevents Smo from transducing the
Hh signal (AlcedoandNoll1997) and has been shown progression, we examined the expression of various markers of differentiation and furrow progression in to antagonize hh function in retinal differentiation
(Chanutand Heberlein1995; Ma and Moses1995; mutant third instar discs. hh transcription was moni-tored using alacZreporter construct inserted at thehh
Wehrliand Tomlinson1995). Its effect on roDom, the
reverse ofhh’s effect, therefore confirmed thatroDomwas locus (Leeet al.1992) and was found identical in
wild-type androDommutant discs (Figure 4, A and B).
a reliable sensor of the integrity of the hh signaling
restores wild-type eye size in hh1 homozygotes (not
shown), it resulted in a strong enhancement of theroDom
stop-furrow phenotype (Figure 5C). Therefore, in aroDom
mutant background, excess Hh signaling can further inhibit differentiation, rather than promote it. Further-more, the stop-furrow phenotype was substantially sup-pressed when excess Ptc was provided ahead of the MF, using a UAS-ptc line (Rørthet al. 1998) and a GAL4
driver under the control of thehairypromoter (h-GAL4;
HuangandFischer-Vize1996). Excess Ptc in anterior cells is expected to reduce their ability to respond to Hh as the MF advances. The fact that it can alleviate furrow arrest suggests that inroDommutants, furrow cells
have an abnormal response to Hh signaling. They may activate an inhibitor that interferes with expression of Hh’s transcriptional targets, likedppand ato. Alterna-tively, they may have an increased sensitivity to Hh sig-naling that makes them adopt fates normally encoun-tered further posteriorly, closer to the source of Hh, where neitherdppnoratoare normally expressed.
roDomis rescued by excess Ato protein:To determine
whether decreasedatoexpression was sufficient to
ex-Figure 3.—roDom is modified by mutations in ato and in
plain furrow arrest inroDom, we forced expression of a
components of thehhsignaling pathway. SEMs ofroDom/⫹(A);
UAS-atoconstruct (Jarmanet al.1993) anterior to the
roDom/ato1 (B); roDom/hh13c (C); roDom/ptcS2 (D) adult eyes. The
MF with theh-GAL4driver. While this had no effect in
reduction of eye size inroDom/⫹is enhanced by loss ofatoand
a wild-type background (not shown), it led to nearly
ptcand suppressed by loss ofhh.In all panels, posterior is to
the right. complete rescue of eye size in the roDom background
(Figure 5E). Conversely, loss-of-function alleles of ato
acted as dominant enhancers ofroDom (Figure 3B). In
contrast,dppoverexpression had no effect onroDomeye
theTGF- homologdpp(Heberleinet al. 1993;Maet size (not shown). We conclude that theroDomphenotype al.1993) and the proneural geneato(Domi´nguezand results primarily from loss ofatoexpression in and ahead
Hafen1997; Borodand Heberlein1998), were also of the MF.
examined. Both are transcribed in an overlapping band groandHaffect different aspects ofatoexpression: of cells in the anterior portion of the MF (Figure 4, Several alleles of Hand gro were recovered as strong C and E), and expression of both genes was sharply suppressors of roDom (Table 2 and Figure 6, A and B).
decreased inroDomdiscs (Figure 4, D and F;Heberlein
While both genes act in theN signaling pathway, they
et al. 1993). Importantly, roDom did not cause reduced
function at different steps and have opposite effects: transcription of all genes in the eye-disc anterior por- H antagonizes signaling by preventing Suppressor of tion. For instance, expression ofhairy(h), an inhibitor Hairless [Su(H)] from entering the nucleus upon activa-of neural differentiation that slows the anterior progress tion of the receptor N (Schweisguth andPosakony of differentiation (Brownet al.1995), remained strong 1994). Gro mediates the inhibition of proneuronal gene and even appeared to expand ahead of the MF (Figure expression by acting as a corepressor with members of 4, I and J). Unlikedppandato,h is not thought to be the Enhancer of split [E(spl)] complex, whose transcrip-a direct ttranscrip-arget ofhh(GreenwoodandStruhl1999). tion is activated by Su(H) (Oellerset al.1994;
Heitz-We conclude that roDom affects furrow progression by
ler et al. 1996). To understand how loss-of-function interfering with Hh signaling and preventing the imple- mutations in both genes may lead toroDomsuppression,
mentation of furrow-specific cell fates. we analyzed the effect of mutant somatic clones on
hhsignaling inroDominhibits differentiation:roDomcould
retinal development. Mutant clones were induced using affect furrow cells in two ways: it could reduce their a heat-induced (hs-FLP) or eye-specific (ey-FLP) FLP re-ability to transduce the hh signal, or it could induce combinase construct and chromosomes carrying thegro
an aberrant response tohh signaling. To address this andHmutations and anFRTrecombination sequence question, we tested the effects of excess Hh and excess (XuandRubin1993). Clonal phenotypes were assessed Ptc on differentiation inroDom.
in adults or in third instar eye-antennal discs (see
mate-Excess Hh was provided using a UAS-hh construct rials and methods).
Figure4.—roDom interferes with expression ofhhtargets in the MF. (A and B)hhtranscription as revealed by-galactosidase
activity staining of third instar discs from wild-type (A) androDom/⫹(B) larvae carrying alacZreporter gene inserted at thehh
locus.hhis expressed in cells differentiating behind the MF (arrowhead) and in the ocellar region (arrow) at similar levels in
roDom/⫹ and wild type. (C and D) Double staining for neuron-specific nuclear antigen ELAV expression (brown) to reveal
differentiation and-galactosidase activity (blue) to reveal the MF (arrowheads) in wild-type (C) androDom/⫹mutant (D) third
instar discs carrying a dpp-lacZreporter gene. Expression of thedpp-lacZreporter is sharply reduced in the central portion of the MF in theroDom/⫹disc (arrow). As the disc ages, loss ofdppexpression affects the whole dorso-ventral span of the MF (not
shown). (E–J) Confocal miscroscopy images of the MF of third instar wild-type (E, G, I) androDom/⫹(F, H, J) discs stained to
reveal Ato in green and ELAV (E and F), Ro (G and H), or Hairy (I and J) in red. The anterior, MF-specific band of Ato expression is sharply reduced in the mutant relative to wild type. Ro and Ato maintain complementary expression patterns at the front of differentiation inroDom/⫹discs, although expression of both proteins is strongly reduced. Expression of Hairy anterior
to the MF is not abolished inroDom/⫹mutant discs but appears instead to expand further anteriorly than in wild type.
tidia appeared to have fused, and photoreceptor clusters tors. Boss is specific to R8 cells and appears in wild type as a regular array of single staining cells behind the MF were commonly found in groups of 20 or more, instead
of the normal complement of 8. Rhabdomeres were (Figure 7C;Caganet al.1992). In contrast,gromutant clones contained tight clusters of Boss-expressing cells, often deformed and elongated, and clusters of small
rhabdomeres, typical of inner photoreceptors R7 or R8, suggesting that adjacent R8 precursors can coexist in
gromutant tissue. Nevertheless, large gaps persisted be-were common.
In third instar eye discs, staining with an antibody tween these R8 clusters, indicating that other photore-ceptor types must also account for neuronal hyperplasia. directed against the neuronal nuclear protein ELAV
(RobinowandWhite1991) revealed a greater density Indeed, staining with Bar and Rough antibodies re-vealed abnormal numbers of R1-6 cells amonggro mu-of cells differentiating as neurons in gromutant tissue
compared to neighboring wild-type tissue (Figure 7B). tant clusters (not shown).
The existence of multiple Boss-expressing cells ingro
While individual clusters were still visible immediately
behind the MF, neuronal differentiation seemed contin- mutant tissue suggests a deregulation ofatoexpression, leading to excess R8 photoreceptor differentiation. This uous in more mature (i.e.,posterior) areas of the disc
(Figure 7B). Neuronal hyperplasia was restricted to the was confirmed by staining with the anti-Ato antibody.gro
mutant clones that straddled the MF showed a posterior mutant tissue and therefore appears to be a cell
photorecep-Figure5.—roDomis rescued by excessatoand
enhanced by excesshh.Light microscopy image of the left eye ofroDom/⫹females carrying various
combinations of GAL4 driver and UAS re-sponder transgenes. In all panels, posterior is to the right. (A and B) Left eye ofroDom/⫹females
carrying one copy of the driver construct GMR-GAL4(A) or hH10-GAL4 (B). The effect of the
drivers onroDom/⫹eye size is negligible (blurry
eye surface results from photographing heads under alcohol). (C)GMR-GAL4drives UAS-hh
expression in differentiating cells behind the MF. Excesshhexpression in its normal location reduces roDom/⫹ eye size. (D and E)hH10-GAL4
drives expression of UAS-ptc (D) and UAS-ato
(E) ahead of the MF. Both conditions result in robust increase of theroDom/⫹eye size.
pattern eventually took place further posteriorly, but it the anterior span of ato expression. Near the MF, the posterior edge ofatoexpression was often seen to bulge often left clusters ofato-expressing cells instead of the
single ato-expressing R8 precursors seen in wild type forward in mutant tissue. This suggests that the refine-ment of ato expression occurred prematurely in the (Figure 7E).
groclones were also occasionally accompanied by se- absence of H protein. Behind the MF, ato-expressing cells were irregularly spaced and sometimes seen in vere overgrowth. This was most obvious in older discs
clusters of two or three (Figure 8, D, G, and H). Whether and correlated with the presence of clones that reached
the effect ofHmutant tissue onatoexpression was cell the anterior and lateral disc margins (Figure 7, F and
autonomous or not could not clearly be established. G). Overgrowth was not usually associated with ectopic
Upregulation ofatoin anterior clones seemed confined differentiation, as shown by the absence of ectopic ELAV
to the boundaries of the clone (Figure 8, D and E). or Ato staining. In contrast, internal clones and clones
However, downregulation behind the MF sometimes touching the posterior margin rarely caused overgrowth
appeared stronger at the center of large clones than at and were always accompanied by neural hyperplasia.
their edges, suggesting some rescue from the neigh-The effect of H mutations was assessed in similar
boring wild-type tissue (Figure 8F). experiments. In adults, clones of ourH299allele led to
Our observations demonstrate a complex effect of mild roughening of the retinal surface. Sections
re-H on ato expression during retinal differentiation. H
vealed mutant ommatidia in regular arrays, but
incom-repressesatoexpression in territories far ahead of the plete, most often lacking one or two outer
photorecep-MF, while behind the MF it ensures the timely selection tors. Ommatidia with two to three inner photoreceptors
of regularly spaced R8 precursors, presumably by modu-were also frequent (Figure 8A). Stronger defects modu-were
lating N signaling. observed with the null allele HE31 (not shown),
sug-gesting thatH299is a hypomorph.
We used HE31, which also acts as a strong suppressor
DISCUSSION
ofroDom(not shown), to analyzeHfunction in disc clones.
Staining for ELAV revealed clusters with reduced num- Genetic screens based on dominant interactions with bers of differentiating cells compared with wild-type a gain-of-function mutation represent a powerful and clusters of similar age (Figure 8, B and C), as expected popular means of identifying components of various from the adult sections. Effects onatoexpression were developmental pathways in flies (Karimet al.1996;
tage ofroDom, a dominant stop-furrow mutation that lends
itself to both enhancement and suppression, to uncover novel components of retinal differentiation. We screened through 130,000 EMS- and X-ray-induced mutants and recovered 103 mutations that either suppressed or en-hanced theroDomreduced eye phenotype. We were
inter-ested in recovering dominant modifiers ofroDomthat were
recessive lethal, as they would likely represent genes that had not been identified in previous screens for defects in adult eye morphology. In particular, we expected mutations that affecthhsignaling, since Hh is required for normal furrow movement andhhmutations display strong genetic interactions withroDom
. How does roDom work? Loss-of-function
ro mutations cause eye roughness, due to mis-specification of photo-receptors R2 and R5, and the formation of ommatidia with more than one R8 photoreceptor (Heberlein et al.1991). Repression of R8 cell fate has been attributed to inhibition ofatoexpression by the Ro homeodomain protein (Dokucu et al. 1996). In support of this pro-posal, Rough and Atonal proteins appear in comple-mentary sets of cells behind the MF, andatoexpression is expanded behind the MF inro mutants (Kimmel et al.1990;Dokucu et al. 1996). Generalized expression ofro under a heat-shock promoter (hs-ro) leads to loss of atoexpression in the MF (Dokucuet al.1996) and eventually results in furrow arrest (Kimmelet al.1990). We find that furrow arrest inroDomis also accompanied
Figure6.—Restoration of eye size by various EMS-induced
by loss of atoexpression in the MF. By analogy to the
suppressors ofroDom recovered in this screen. SEMs of roDom/ hs-ro phenotype, we propose thatroDom leads to excess
H299(A);roDom/gro347(B);roDom/1480(C); androDom/519(D). In
Ro production, although that excess was not detectable all panels, posterior is to the right. by antibody staining.
roexpression at the posterior edge of the MF is under
the control of hh signaling (Domı´nguez 1999). The among other things, components ofhh signaling that affectrooratoexpression or partners of Ro in the inhibi-position ofro-expressing cells, adjacent tohh-expressing
cells, suggests that high levels of hh signaling are re- tion ofatotranscription.
Expression of dpp is also sharply decreased in roDom.
quired forroexpression. By comparison,ato, also a
tar-get ofhhsignaling in the MF (Domı´nguezandHafen Likeato,dppexpression could be inhibited byrodirectly. This may explain its sharp downregulation behind the 1997;BorodandHeberlein1998), is expressed further
away from thehhsource, suggesting a requirement for MF in wild type at the location where ro begins to be expressed. Alternatively, its decrease inroDomcould be a
lower levels of hhsignaling. We propose that the roDom
rearrangement sensitizes the ro gene to hh signaling, secondary consequence of decreasedatotranscription. In support of the latter, dpp transcription is sharply either by removing a negative regulatorycis-element or
by bringing in an additional hh-responsive enhancer reduced in the MF ofato1homozygous larvae (
Jarman
et al. 1995). Surprisingly, Hairy protein levels remain element. The resulting anterior expansion of the ro
expression domain would prevent ato expression and elevated ahead of the MF inroDom, although
hhas been shown to be a target of Dpp signaling (Greenwoodand ultimately cause differentiation arrest.
While this model cannot be proven at this point, it Struhl1999). The same is true inato1mutants (Jarman et al. 1995). This suggests that h is under the control provides a simple explanation for the surprising genetic
interactions between roDom andhh: if the rearrangedro of other, as yet unidentified, mechanisms that are not
dramatically impaired by the roDom mutation and the
gene is more sensitive to Hh, then increasinghh gene
dosage will cause more Ro production and accelerate accompanying loss of dpp transcription. In any case, effects ofroDomon dppandh expression are unlikely to
the differentiation arrest. On the other hand, reducing
Hh signaling, by removing one copy ofhhor by provid- explain the furrow arrest since the roDom phenotype is
not detectably affected by changes of h and dpp gene ing the inhibitor Ptc in excess, will diminish the amount
of Ro protein made and restore Ato accumulation. Mod- dosage.
In contrast,roDomis very sensitive to alterations ofato
Figure 7.—Effect of gro
somatic mutant clones on retinal differentiation. (A) Tangential section through an adult retina carryinggro347
clones. The blue arrow points to a typical wild-type omma-tidium with a single inner photoreceptor (small rhab-domere) surrounded by six outer photoreceptors (large rhabdomeres) and a crown of pigment cells. Mutant clones are marked by the ab-sence of red pigment (see materials and methods). Yellow arrows point to large mutant clones with fused ommatidia that lack pig-ment cells and contain an ex-cess of disorganized and misshapen inner and outer photoreceptors. (B–G) Third instar discs carrying gro347
clones and stained with anti-bodies (brown) against the neuronal marker ELAV (B, F), the R8-specific cell sur-face antigen Boss (C), or the retinal proneural protein Ato (D, E, G). Clones are marked by the absence of the cell-autonomous markerarm-lacZas evidenced by-galactosidase activity staining (blue). Red arrowheads indicate the approximate location of the MF. Posterior is to the right. (B)gro347clones behind
the MF fill up with ELAV-expressing cells (white arrows), indicating neuronal hyperplasia. (C) Boss-expressing cells form tight clusters ingro347clones behind the MF (white arrows), instead of the evenly spaced single cells seen in neighboring heterozygous
tissue (pink arrow) or in the homozygous wild-type twin-spot (blue arrow). (D) In mutant clones that span the MF, furrow-specific Ato expands further posteriorly (white arrow) than in neighboring wild-type or heterozygous tissue. (E) Behind the MF, Ato persists in cell clusters (white arrows), instead of resolving to single cells. (F and G) Mutant clones that reach the anterior and lateral disc margins are associated with severe overgrowth without ectopic differentiation, as evidenced by ELAV (F) or Ato (G) detection. Neuronal hyperplasia is observed in mutant clones located behind the MF (white arrows) or in the ocellar precursor region (red arrows). Black arrows point to epithelial folds indicative of overgrowth.
gene dosage, as it is enhanced by loss-of-function ato nezet al. 1997;Fisher and Caudy1998). Association of Gro with Hairy deserves to be envisaged here, since alleles and almost completely rescued when high levels
of ato expression are restored ahead of the MF. The Hairy has been implicated in inhibition of ato as well (Brownet al.1995). We find, however, thatgromutant
roDom phenotype therefore appears to result primarily
from inhibition ofatoexpression due to excess Ro pro- clones expand ato expression posterior to the MF, whereas h inhibits ato expression anterior to the MF tein. On the basis of this understanding, we have
ana-lyzed the role of two of the strongest suppressors isolated (Brown et al. 1995). Another intriguing possibility is that Gro associates with Ro to mediate inhibition ofato
in our screen, new alleles ofgroandH, onatoregulation
and furrow progression. expression behind the MF. Although we cannot com-pletely eliminate this hypothesis, we find it unlikely,
groucho: gro encodes a transcription inhibitor that
combines with b-HLH genes of the E(spl) complex to because complete loss-of-function phenotypes ofroand
groare different. While both of them lead to increased inhibit expression of proneural genes such as achaete
and scute (Heitzler et al. 1996). We find that in gro atoexpression and imperfect R8 resolution, this effect is much more extensive and long lasting ingromutant mutant clones, expression ofatopersists behind the MF
longer than in wild-type tissue. This is consistent with tissue (our data) than in ro mutant tissue (Kimmel et al.1990;Heberleinet al.1991;Dokucuet al.1996). In a role for Gro in the N signaling events that lead to the
refinement ofatoexpression behind the MF. addition, neuronal hyperplasia is not observed in ro
mutant tissue, which suggests that at least thisgro func-However, Gro is also known to form inhibitory
com-plexes with other transcription factors of the b-HLH tion must involve factors other than Ro. On the other hand, removal of E(spl) function results in neuronal class, such as Hairy, or of other classes, such as the
c-Rel homolog Dorsal or the homeodomain, segment hyperplasia and excess R8 development very similar to removal ofgrofunction (Treismanet al.1997;
Jime´-Figure 8.—Effect of H so-matic mutant clones on retinal differentiation. (A) Tangential section through an adult re-tina carrying anH299clone. The
clone is marked by the absence of red pigment (seematerials and methods). Mutant omma-tidia are arrayed regularly but contain an abnormal comple-ment of photoreceptors. Some have extra inner photorecep-tors and lack outer photore-ceptors (green circle); others lack inner photoreceptors (yel-low circle). (B–H) Third instar discs carryingHE31clones and
stained with antibodies (brown) against the neuronal marker ELAV (B, C, C⬘) or against Ato (D–H). Clones are marked by the absence of the cell-autono-mous clonal markerarm-lacZas revealed by-galactosidase ac-tivity staining (blue). Red ar-rowheads indicate the
approxi-mate location of the MF.
Posterior is to the right. (B, C, C⬘) Mutant ommatidia contain fewer ELAV-staining cells than wild-type ommatidia of the same stage, appear mis-ori-ented (C, close-up of area marked in B), and are inter-spersed with picnotic vesicles (black arrows) indicative of cell death (C⬘, close-up of another
HE31clone). (D–F) HE31clones
that straddle the MF are asso-ciated with accelerated resolu-tion of Ato expression behind the MF (D, blue arrow), expansion of Ato anterior to the MF (D and E, red arrows), or loss of Ato in the MF (F, black arrow). (G and H) Higher magnification views of HE31mutant clones near the MF, revealing uneven
Ato production in the MF and irregular spacing or abnormal clustering of Ato-expressing cells behind the MF.
gakis et al. 1998). Therefore, we favor the hypothesis receptors was observed with a hypomorphic gro allele in the absence of excess R8 differentiation (
Fischer-that Gro restrictsatoexpression by combining with
pro-teins of theE(spl)complex whose expression is induced Vizeet al.1992). The excess of all photoreceptor types observed with our strongergroallele may therefore re-by N signaling.
Even in the complete absence ofgro[orE(spl)] func- flect an involvement ofgroin the restriction of cell fates at each step of ommatidial formation.
tion, some refinement of ato expression still occurs,
which indicates that factors independent of N and Gro gro mutant clones can also induce extensive over-growth of head capsule and retinal tissues. In the wing, play important roles in patterning Ato behind the MF.
Candidates include Ro (Dokucuet al.1996), the EGFR groclones have been found to cause overgrowth via the induction of ectopichhexpression (de Celisand
Ruiz-inhibitor Argos (Spenceret al.1998), and Hh (
Domı´n-guez1999). Moreover, outer photoreceptors differenti- Go´ mez 1995). Hh is also a powerful inducer of over-growth in eye discs, when provided in excess or ectopi-ate in large excess between the R8 precursors and are
the main cause of neuronal hyperplasia (Treisman et cally (ChanutandHeberlein1995; Heberleinet al.
1995). However, overgrowth due to ectopichh
expres-al.1997;Ligoxygakiset al.1998). Neuronal hyperplasia
could occur as a direct consequence of the excess of sion is accompanied by ectopic and premature photore-ceptor differentiation, a phenotype we do not observe R8 precursors in gro[and E(spl)] mutant tissue, which
would, through the normal serial induction process, in overgrowngromutant tissue. It is therefore unlikely that gromutant clones cause ectopic hh expression in recruit an excess of neighboring cells into ommatidial
production, we would expect enhancement, rather than which act in opposite direction in the N pathway, is not unique. A similar situation was encountered in another suppression, of theroDomphenotype (see above). While
scenarios in which a slight increase in cell proliferation study where mutations in gro andH were both found to enhance the wing and bristle phenotypes associated allows the MF to progress further in roDom cannot be
eliminated, we find it more likely that gro mutations with loss-of-function mutations in EGFR (Price et al.
1997). Our observation that mutations in both genes suppressroDomby allowing Ato protein to persist longer
in the MF. elevate ato expression in the vicinity of the MF, but at different stages of the differentiation process, helps
Hairless: Hairless inhibits N signaling by preventing
Su(H), a transcription factor (SchweisguthandPosa- resolve this paradox. Our results also indicate that the exact timing (or location) ofatoexpression might not
kony1992;Schweisguth1995), from translocating to
the nucleus and activating transcription of N targets be crucial to MF progression, provided adequate levels are reached. This conclusion is supported by our finding such as theE(spl)complex genes (Banget al.1991;Bang
and Posakony 1992; Schweisguth and Posakony that Ato supplied anterior to its normal expression do-main, in thehexpression domain, restores normal eye 1994). In the absence of H, Su(H) is free to enter the
nucleus upon activation of N.Su(H)mutant clones lead size in aroDombackground. Whether proper R8 spacing
and ommatidial patterning can be achieved under these to expanded Ato expression behind the MF, consistent
with a role for Su(H) in the N-mediated lateral inhibi- conditions remains to be shown.
tion that leads to the refinement of ato expression We are grateful to Todd Laverty and other members of the fly (Ligoxygakiset al.1998). We find Hclones in which community for supplying fly stocks and antibodies. We thank the Electron Microscopy Lab at UC Berkeley for assistance with SEM. We
the refinement ofatoexpression to single cells appears
thank Dr. Gerry Rubin at UC Berkeley and Dr. Ivan Diamond at the
accelerated. This is consistent with a role for H as an
Ernest Gallo Clinic and Research Center for supporting us in the
inhibitor of N and Su(H) in lateral inhibition. Surpris- early steps of this project. We thank Katherine Woo for suggesting ingly, however, individual clusters of Ato-expressing theUAS-ptcrescue experiment and for assistance with microscopy and cells often persist in Hmutant tissue behind the MF, image processing. Finally, we are grateful to members of the Heberlein lab, past and present, for various contributions to this work, comments
instead of resolving to single R8 precursors; in adults
on the manuscript, and constant support. This work was supported
as well, mutant ommatidia often contain more than one
by grants from the National Institute of Health (EY-11410) and the
R8. This would suggest that at later stages H is required National Science Foundation (IBN-9604216) to U.H. to resolve ato expression to single R8 precursors, a role
that is not expected for an inhibitor of theNpathway.
AnteriorHmutant clones show precociousatoexpres- LITERATURE CITED sion anterior to the MF. This might explain the
pat-Albagli, O., M. P. LagetandF. Chanut,1997 Nearsighted
mor-terning defects behind the MF, if precocious and exces- phogenesis in the Drosophila retina. M S-Med. Sci.13:175–183.
Alcedo, J.,andM. Noll,1997 Hedgehog and its
Patched-smooth-sive accumulation of Ato protein in the MF interferes
ened receptor complex: a novel signalling mechanism at the cell
with the proper execution of lateral inhibition via N or
surface. Biol. Chem. Hoppe-Seyler378:583–590.
with downregulation by Ro. In this regard, we note that Alcedo, J., M. Ayzenzon, T. Von Ohlen, M. NollandJ. E. Hooper,
1996 The Drosophila smoothened gene encodes a seven-pass
excess Ato protein, as provided under heat-shock
con-membrane protein, a putative receptor for the Hedgehog signal.
trol, was found to perturb the resolution ofato
expres-Cell86:221–232.
sion to single R8 precursors (Dokucuet al.1996). Aza-Blanc, P., F.-A. Ramı´rez-Weber, M.-P. Laget, C. Schwartzand
T. B. Kornberg,1997 Proteolysis that is inhibited by Hedgehog
It has been suggested that earlyatoexpression, ahead
targets Cubitus interruptus protein to the nucleus and converts
of the MF, is in part the result of an as yet unsuspected
it to a repressor. Cell89:1043–1053.
“proneural” effect of N signaling (BakerandYu1997). Baker, N. E.,andS. Y. Yu,1997 Proneural function of neurogenic
genes in the developing Drosophila eye. Curr. Biol.7:122–132.
The anterior expansion ofatoexpression inHmutant
Baker, N. E.,andA. E. Zitron,1995 Drosophila eye development:
clones is consistent with this model, assuming that H
Notch and Delta amplify a neurogenic pattern conferred on the
would act as an inhibitor ofNthere as well. However, morphogenetic furrow by scabrous. Mech. Dev.49:173–189.
Baker, N. E., S. YuandD. Han,1996 Evolution of proneural atonal
the proneural function ofNmust not be mediated by
expression during distinct regulatory phases in the developing Su(H), since removal ofSu(H)function did not abolish
Drosophila eye. Curr. Biol.6:1290–1301.
ato expression ahead of the MF (Ligoxygakis et al. Bang, A. G.,andJ. W. Posakony,1992 TheDrosophilageneHairless encodes a novel basic protein that controls alternative cell fates
1998). Our results may indicate thatHantagonizes the
in adult sensory organ development. Genes Dev.6:1752–1769.
proneural function ofNvia a mechanism that does not
Bang, A. G., V. HartensteinandJ. W. Posakony,1991 Hairlessis
involveSu(H). Alternatively, the role of H on early ato required for the development of adult sensory organ precursor cells inDrosophila.Development111:89–104.
expression may be independent of N signaling.
Regard-Bang, A. G., A. M. BaileyandJ. W. Posakony,1995 Hairless
pro-less of the exact mechanism, the enhanced expression
motes stable commitment to the sensory organ precursor cell
ofatoahead of the MF inHmutants is likely to explain fate by negatively regulating the activity of the Notch signaling
pathway. Dev. Biol.172:479–494.
suppression of theroDomphenotype by counteracting the
Blackman, R. K., M. Sanicola, L. A. Raftery, T. GillevetandW. M.
effect of ectopic Ro onatoexpression in the MF.
Gelbart,1991 An extensive 3⬘cis-regulatory region directs the
Finding that similar levels of suppression can be imaginal disk expression of decapentaplegic, a member of the
TGF-family inDrosophila.Development111:657–666.