Effectiveness of parental provisioning under variable oceanographic conditions along the Western Australian coast in two shearwater species: the little shearwater Puffinus assimilis tunneyi and the wedge-tailed shearwater Puffinus pacificus
Full text
Figure




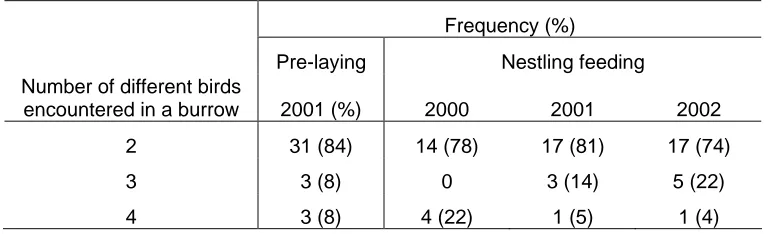
Outline
Related documents
In this paper, we argue that for such problems, Empirical Risk Minimization can be implemented using statistical counterparts of the risk based on much less terms (picked randomly
joining the centroid of the ligand ring and the metal atom. Non-H atoms are shown as 50% probability displacement ellipsoids and H atoms have been omitted for.
In our case of UC with oncocytic fea- tures, the Ki-67 labeling index was especially low in the tumor cells with oncocytic morpholo- gy corresponding to the observation that
ADEM: Acute disseminated encephalomyelitis; BBB: Blood-brain barrier; CCSVI: Chronic cerebrospinal venous insufficiency; CNS: Central nervous system; CSF: Cerebrospinal fluid;
Male notification rates were significantly higher for the 0 to 4 years old age group regardless of virus type or subtype; and higher for those aged 0 to 14 years and those 85 years
In this work we deal with the technical analysis of the villa Tugendhat interior furnishing, or more precisely with the structural analysis of its furniture joints. The goal of
Nutritional profit obtained from uptake of dissolved amino acids from natural sea water in an experimental flow-through
Relevant clinical subsets may be defined by different age of onset, different disease course by degree of severity, extrinsic (allergic sensitization detectable) and intrinsic
