Behavioural and morphological defences in a
fi
sh with a complex
antipredator phenotype
Van Wishingrad
a,*, Maud C. O. Ferrari
b, Douglas P. Chivers
a aDepartment of Biology, University of Saskatchewan, Saskatoon, SK, CanadabBiomedical Sciences, WCVM, University of Saskatchewan, Saskatoon, SK, Canada
a r t i c l e i n f o
Article history: Received 27 March 2014 Initial acceptance 9 May 2014 Final acceptance 25 June 2014 Available online 9 August 2014 MS. number: A14-00256R
Keywords: Acipenser fulvescens antipredator behaviour defensive morphology escape behaviour habitat use lake sturgeon phenotypic integration
A number of prey species rely on two primary adaptations, behavioural alterations and morphological defences, to reduce their probability of being killed by predators. There should be considerable selection for maintaining adaptive trait combinations between behavioural and morphological defences in natural populations. However, our understanding of how behaviour and morphology interact has been limited to somewhat simple systems. Here we investigate patterns of covariation between behaviour and morphology in juvenile lake sturgeon,Acipenser fulvescens, afish species with a complex antipredator phenotype. We found strong evidence for trait co-dependence between escape responses and body size, where largerfish were able to show stronger escape responses. Furthermore, we found that cover-seeking behaviour exhibited a complex multitiered relationship, representing a mixture of trait compensation and trait co-specialization that was dependent on specific combinations of mechanically independent traits. The smallestfish sought cover in rocky microhabitats more thanfish of any other size (i.e. trait compensation). However, the largestfish also increased the amount of time they spent in rocky microhabitats following exposure to risk, indicating that cover-seeking behaviour may co-specialize with the ability to show strong escape responses. Crypticfish of intermediate size did notflee from foraging grounds in response to risk (i.e. they showed trait compensation), whereasfish that lacked this pattern
fled. Our results demonstrate how behaviour can integrate with complex morphological variation to produce unique antipredator outcomes. As sturgeon morphology has been largely preserved since the lower Jurassic period (~200 million years ago), it may represent a highly successful morphological trait complex to which behavioural responses arefinely tuned. Insight into the extent to which behaviour is linked to morphology is important in order to understand the limitations or reaches of adaptive phenotypic plasticity and the evolution of integrated phenotypes.
©2014 The Association for the Study of Animal Behaviour. Published by Elsevier Ltd. All rights reserved.
Predation is a strong selective force shaping the lives of prey animals. Not only can predators reduce prey numbers through direct consumptive effects, they cause nonconsumptive effects influencing an animal's morphology, life history and behaviour, including when, where and how they forage and reproduce (Preisser, Bolnick,&Benard, 2005). An animal's success in avoiding predators is an integration of multiple defences; therefore, there should be considerable selection for maintaining adaptive trait combinations in natural populations (Sinervo&Svensson, 2002).
Perhaps the most well-known example of correlational selec-tion between antipredator behaviour and morphology is that of garter snake (Thamnophis ordinoides) escape behaviour and the pattern on their back (Brodie, 1989, 1992). When fleeing in a
straight line in the wild, snakes with striped back patterns have significantly higher survival rates than barred snakes, but when
fleeing while reversing, barred snakes have significantly higher survival rates than striped snakes (Brodie, 1992). In this case, spe-cific body patterns and behaviours that increased survival led to genetic correlations between morphology and antipredator behaviour (Brodie, 1989).
Dewitt, Sih, and Hucko (1999)proposed a framework for un-derstanding how antipredator traits interact. Mechanically inde-pendent traits can interact in an additive way (trait co-specialization), one trait can compensate for another (trait compensation), or traits may need to be combined in order to be efficient (trait complementation), andfinally, mechanically linked traits may rely on each other to function (trait co-dependence). In this influential paper,Dewitt et al. (1999)provided evidence of trait compensation and co-specialization in a freshwater snail, which sparked a great deal of research exploring how correlated traits
*Correspondence: V. Wishingrad, Department of Biology, University of
Sas-katchewan, 112 Science Place, Saskatoon, SK S7N 5E2, Canada. E-mail address:[email protected](V. Wishingrad).
Contents lists available atScienceDirect
Animal Behaviour
j o u r n a l h o me p a g e : w w w . e l s e v i e r . c o m / l o ca t e / a n b e h a v
http://dx.doi.org/10.1016/j.anbehav.2014.07.006
evolve. Examining how correlated traits evolve may help us un-derstand thefitness consequences of behaviour and the evolution of integrated phenotypes.
Individual differences in morphology can influence how in-dividuals within a species behave. For example,Pettersson, Nilsson, and Br€onmark (2000)found that crucian carp,Carassius carassius, from populations that coexist with predators, and, as a conse-quence, have developed deep bodies, show weaker fright re-sponses. Likewise, deep-bodied goldfish,Carassius auratus, show a weaker response to predator odours than do shallow-bodied con-specifics (i.e. trait compensation;Chivers, Zhao,&Ferrari, 2007). Trait compensation is one way that animals can achieve compara-ble total fitness while expressing antipredator behaviours to different degrees (Dewitt et al., 1999). Thus, antipredator behaviour, in and of itself, may not be a strong surrogate of truefitness because resource allocation away from antipredator behaviour and towards morphological defences may ultimately have equivalent fitness consequences (Lind&Cresswell, 2005).
Differences in morphology between species may also correlate with differences in antipredator behaviour.Abrahams (1995)found that brook stickleback, Culea inconstans, which have a series of spines on their dorsal surface, show little behavioural modification in response to a predator, while unarmoured fathead minnows,
Pimephales promelas, respond significantly more. Similarly,Rundle and Bronmark (2001)€ found evidence of species-specific patterns of trait compensation in predator defence adaptations across several species of pulmonate snails. These studies provide evidence that morphological differences between species are associated with predictable differences in antipredator behaviour. However, behaviour of distantly related species used in these types of comparative studies may be the result of differences in life history or body shape, and thus, may obscure the true relationship be-tween morphology and behaviour (Mikolajewski & Johansson, 2004). To help address this problem,Mikolajewski and Johansson (2004) investigated the relationship between behaviour and morphological defence (abdominal spine length) in larvae of three dragonfly species within the genusLeucorrhinia. They found that the three species showed a combination of trait compensation and co-specialization, where the species with spines of intermediate length (Leucorrhinia dubia), rather than the species with the longest spines (Leucorrhinia albifrons), displayed the weakest antipredator response. These results suggest thatL. albifronsmaximizes overall defence, L. dubiaattempts to save on the cost of morphological defence by being more active, while the species with the shortest spines (Leucorrhinia rubicunda) compensates for limited morpho-logical defence by displaying the strongest antipredator behaviour. One potential confounding factor, however, could be differences in predator abundance between lakes from which dragonfly larvae were collected, which may have driven the results to a certain extent (Mikolajewski&Johansson, 2004).
Understanding the interplay between morphology and behav-iour continues to be an exciting area of research. However, one major deficit in our understanding of how behaviour and morphology interact is that it has so far been limited to somewhat simple systems, mostly having to do with various properties of shell morphology in snails, body size or armour infish and spine length in dragonfly larvae. Furthermore, how behaviour changes alongside the development of defensive morphological adaptations in the absence of potential confounding effects of predator exposure is not well understood. Finally, the relative degree to which several complementary morphological traits covary with antipredator behaviour has not been examined. Lake sturgeon, Acipenser ful-vescens, are an ideal species in which to investigate these questions. Antipredator behaviours include increased activity (an escape response) and increased residency in rocky habitats (cover seeking)
(Wishingrad, Chivers,&Ferrari, 2014). Antipredator morphological adaptations consist of five rows of scutes (armour/spines), a mottled (cryptic) body pattern that develops in juveniles and fast-growing bodies (Peterson, Vecsei,&Jennings, 2007). Furthermore, lake sturgeon are an ideal system in which to investigate behav-ioural changes during morphological development because they undergo a dramatic morphological change in a relatively short period. The extent of these morphological changes and the rate at which they proceed are so dramatic that it resembles a meta-morphosis (Peterson et al., 2007). Finally, since lake sturgeon have several different morphological defence traits, unravelling the de-gree to which antipredator behaviours covary across morphological variation may be possible.
In the present study we investigated how escape behaviour and cover seeking vary in response to changes in several independent morphological traits. We also investigated how these two anti-predator responses change as morphological development pro-ceeds. We predicted that, as these morphological defences developed, antipredator behaviour would decrease (i.e. trait compensation) because this is the most common way behavioural and morphological traits interact (Mikolajewski & Johansson, 2004). We also investigate the hierarchical structure of behav-ioural dependence on morphology (i.e. which morphological ad-aptations are most strongly linked to behaviour), but given the lack of available data, we are unable to make any reasonable a priori predictions about this outcome, so we approach this component in a purely exploratory way.
METHODS
Study Species
We obtained lake sturgeon from Sustainable Sturgeon Culture, Ontario, in May 2012 where they were spawned from wild broodstock endemic to the Rainy River, Ontario. They were housed in the RJF Smith Centre for Aquatic Ecology, held in a 640-litrefl ow-through tank and fed daily with liveArtemiaand crushed frozen Chironomidae. They were maintained in dechlorinated tap water at 16e18C under a 14:10 h light:dark cycle.
Experimental Set-up
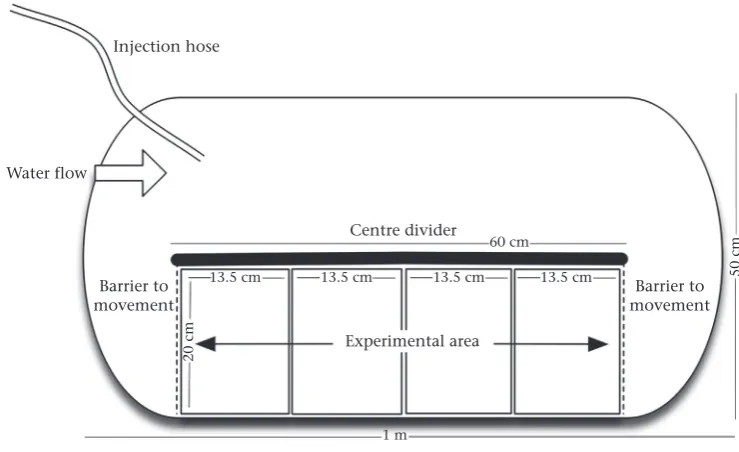
Trials took place in 100-litre polyurethane D-ended mesocosms measuring 1 m long50 cm wide30 cm high (Fig. 1).
Each mesocosm was divided in half longitudinally with an opaque acrylic panel measuring 60 cm long to allow the circular
flow of water with a submersible powerhead (15 litres/min), which provided aflow of 2e3 cm/s. A rectangular section (5520 cm) of the long edge of the mesocosm was segregated using vinyl-coated
height of 5.5 cm over the substrate boxes to control for use of the water column during trials. The experimental area comprised the rectangular section of the mesocosm containing four microhabitat boxes, two of each type, between the mesh barriers.
Stimulus Preparation
Antipredator responses were elicited with alarm cues (~conspecific damage-released cues) that increase the perceived level of danger in the environment where they are released (Chivers&Smith, 1998; Ferrari, Wisenden,&Chivers, 2010). We usedfive donor sturgeon to prepare the alarm cue solution the day before the trials were conducted. Sturgeon were euthanized by cervical dislocation, followed by a concussive blow to the head, then placed in chilled dechlorinated water and homogenized using a Polytron homogenizer (Brinkman Instruments, Rexdale, ON, Canada). Then, the solution wasfiltered through glass wool and diluted to a concentration of 25 cm2of skin/litre. Solutions were frozen in 50 ml aliquots until used. We could not anaesthetize the sturgeon prior to euthanizing them because of the potential contamination of the alarm cue solution with the anaesthetic. However, our euthanasia method was rapid and resulted in mini-mal suffering.
Experimental Protocol
Trials took place in June and July 2012 when thefish were be-tween 42 and 69 days posthatch, and again in September and October 2012 when the fish were between 130 and 148 days posthatch. Eighty trials were run between June and July and be-tween September and October (20 alarm cue/20 water control in each group). Fish were arbitrarily selected from a holding tank and threefish were placed in the experimental area of each tank and left to acclimate for 48 h prior to the onset of trials. Trials were conducted between 2300 hours and 0200 hours. Trials were con-ducted at night because this is the time whenfish forage most actively and are therefore susceptible to detection by predators. Incident light at an approximate intensity of 0.25 lx (equivalent to the light of a full moon on a clear night) provided a level of illu-mination that would likely be encountered during at least some
nights in the wild. A small amount of crushed bloodworm was added to each mesocosm 1 h prior to behavioural observations. Introducing food to the mesocosms at this time guaranteed that the
fish would forage actively at the onset of trials so trade-offs be-tween foraging and antipredator behaviour could be assessed. All trials were recorded using overhead cameras mounted with infrared lights. Before the onset of trials, 60 ml of water from each mesocosm was withdrawn through a 2 m long injection hose and discarded (to remove stagnant water from the injection hose), then 60 ml of water from each mesocosm was withdrawn and retained (toflush the stimuli completely into the mesocosm after injection). Trials began with a 7 min pre-injection observation period (to measure baseline habitat selection and foraging activity level), followed by a 1 min injection period of either 10 ml of alarm cue solution or 10 ml of dechlorinated tap water (control), followed immediately by 60 ml of retained water, andfinally completed with a 7 min post-injection observation period. Alarm cue trials were conducted to examine how lake sturgeon respond to risk, while water control trials were conducted to verify that the observed behavioural responses were due to risk cues rather than the in-jection per se. The experimenter was blind to cue treatment (water versus alarm cue) during injections. We introduced alarm cue at a concentration of 1 mm2of damaged skin per litre in the experi-mental active space (Wisenden, 2008) to maintain a constant level of absolute risk. At the completion of trials, focal fish whose behaviour was scored were photographed for morphometric analysis. After photographs were taken, all fish were placed in a separate holding tank and not used for subsequent trials to main-tain statistical independence among trials.
Video Analysis
All videos were scored using JWatcher (version 1.0;Blumstein, Daniel,&Evans, 2006). Activity level was measured by the num-ber of times a randomly selected focalfish passed over a super-imposed 1013.5 cm grid, where the side of each square was equivalent to approximately three tofive body lengths of thefish. The grid pattern corresponded to transitions between each microhabitat division and a horizontal line dividing each micro-habitat in half. Thus, activity level could be scored independently of
Injection hose
Water flow
Barrier to movement
Barrier to movement Centre divider
Experimental area 60 cm
13.5 cm 13.5 cm 13.5 cm
1 m
13.5 cm 50 cm
20 cm
transitions between microhabitats as thefish could either increase activity within one microhabitat, or alternatively, move between microhabitats. Rocky microhabitat occupancy was measured as the proportion of total time the focal fish spent in the rocky micro-habitat. The behaviour of the other twofish was not scored: they were added to provide a realistic social context during the exper-iment as juvenile lake sturgeon are often found in groups (Peterson et al., 2007). The experimenter was blind to cue treatment (water versus alarm cue) during video scoring.
Morphometric Analysis
Dorsal and lateral photographs were taken of each focalfish using a digital camera (Panasonic DMC-FZ28; ~50 pixels/mm on image sensor). Fish were out of the water for no more than 30 s while photographs were taken, which is much shorter than a 2e3 min air exposure that is considered a‘brief’air exposure for lake sturgeon (Baker, Peake,&Kieffer, 2008). Photographs were scored using ImageJ (version 1.45s;Schneider, Rasband,&Eliceiri, 2012). We measured fish length, width and height to estimate body size, and measured the height of each dorsal scute to estimate scute size. All measurements were made to the nearest 0.1 mm; measurements at this scale were highly repeatable (r>0.9). Mottling was absent in the young group and present in the older group. Photographs of representativefish are provided inFig. 2and descriptive statistics for morphometric data are presented in
Table 1.
Statistical Analysis
We analysed our data using R (version 2.15.2;R Development Core Team, 2012). Escape responses were defined as the relative change in line crosses ((postpre)/(pre)). The escape response distribution was logeþ1 transformed prior to analysis to achieve
normality. This transformation was chosen because the distribution was positively skewed, and the minimum value was0.75. Cover-seeking behaviour was defined as the inverse relative change in proportion of time spent in sand microhabitats ((postpre)/(pre)). The distribution of cover-seeking behaviour was normally distrib-uted and not transformed prior to analysis.
We used principal components analysis (PCA) to reduce dimensionality in the body size variables to generate a‘body size’
composite variable from the length, width and height measure-ments (Jolliffe, 2005). Similarly, we calculated the average dorsal scute height to generate a single‘scute size’variable to be used in
the analysis. We used Pearson's test for significant correlations to investigate whether escape responses and cover use in response to alarm cues were significantly correlated. We used regression trees to investigate how each antipredator behaviour (escape behaviour and cover seeking) covaried with different morphologies using the
‘rpart’package (Therneau, Atkinson, &Ripley, 2012). Regression tree analysis produces a predictive tree-like diagram based on recursive partitioning of variance in the response data and gives an indication of the influence of the explanatory variables at each split (node) in the tree (De'ath&Fabricius, 2000; Legendre&Legendre, 2012). The dependent variable was either escape response or cover seeking, and the independent variables were body size, scute size (average of all dorsal scutes) and mottling (present or absent). We used cross-validation to select tree size by selecting trees with the minimum cross-validated standard error (De'ath & Fabricius, 2000).
RESULTS
Behavioural Correlations
Escape responses and cover seeking were not significantly correlated (Pearson correlation:r38¼0.26,P¼0.1), so we analysed
each antipredator behaviour separately to investigate how morphological traits contributed to each response.
Body Size PCA
PC axis 1 (i.e.‘body size’) explained 99.69% of the variation in body size measurements. As would be expected, body size was
Figure 2.Lateral images of lake sturgeon showing the range in size, scute
develop-ment and change in colour pattern over the course of the study. The smallfish (above)
is 45 days old; the largefish (below) is 148 days old.
Table 1
Morphometrics of lake sturgeon used in the study
Morphometrics (mean±SD, mm)
Length Width Height Scutes
Young, mottling absent
(42e69 dph)
40.9±6.8 4.1±0.7 4.0±0.7 0.7±0.2
Older, mottling present
(130e148 dph)
67.0±6.7 8.7±1.0 7.1±0.8 1.2±0.2
dph: days posthatch. Sturgeon are divided into two groups: youngfish (which lack
mottled coloration) and older fish (with mottled body patterns). Length was
measured as total length (tip of rostrum to tip of tail). Width and height were
measured immediately anterior to pectoralfins. Scute size is the average height of
all dorsal scutes.N¼40 per group.
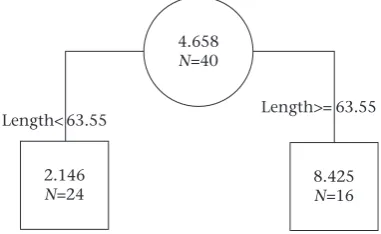
4.658 N=40
2.146 N=24
Length< 63.55 Length>= 63.55
8.425 N=16
Figure 3.Regression tree showing the relationship between escape behaviour
(increased activity) relative to the three morphological variables (fish length, scute
size, body pattern) based on 40 trials. Fish length was the only variable that covaried with escape behaviour. Values at each node indicate the mean proportion change in activity level following exposure to alarm cues, where higher values indicate stronger
escape responses. Values along each branch indicate the thresholdfish length (mm)
positively correlated with length, width and height (allr38>0.95, P<0.001). We therefore used length instead of body size in the subsequent analysis to simplify the interpretation of our results.
Regression Trees
We present the regression tree results where length was used as a predictor variable. In all cases the results obtained using length were identical to those obtained using ‘body size’ (i.e. cross-validation selected the same number of groups). We elected to use length in the analysis because the regression trees would identify specific threshold values, which are easier to interpret as a length than as a body size score from three morphological vari-ables. In addition, the use of length instead of body size allows for an easier comparison between other studies and systems. Thus, in all cases, length (in mm), mottling (present/absent) and scute size (average of all dorsal scutes) were used as predictor variables in the following regression trees.
The escape response regression tree is presented inFig. 3. Fish length was the only morphological variable that predicted escape responses, where larger fish showed a stronger response than smallerfish (average percentage increase in activity of 843% versus 215%).
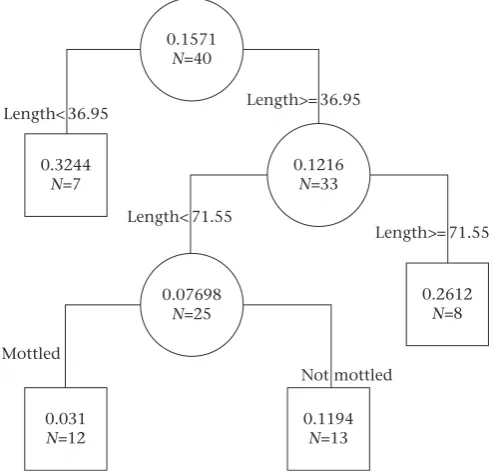
The cover-seeking regression tree is presented in Fig. 4. The smallestfish in the study increased the percentage of time spent in rocky areas the most (~32% increase) after exposure to alarm cues relative to the prestimulus baseline. The largestfish in the study also increased the percentage of time they spent in rocky areas (~26% increase), but less so than the smallestfish. Mottledfish of intermediate size showed a small increase in the percentage of time spent in rocky areas (~3.1%) relative to otherfish of equivalent size
that lacked the mottled body pattern (increase of ~12%) after exposure to alarm cues.
Antipredator Behaviour between Stages and Treatments
Table 2compares the responses of sturgeon to alarm cues at two stages of development. Overall, largerfish (all from the older group) showed stronger escape behaviours in response to alarm cues than smallerfish (which included all those from the younger group and four from the older group). On the other hand, largerfish spent less time in rocky microhabitats in response to alarm cues than smaller
fish. Antipredator responses to water injections were within 1 SE of zero in all treatment combinations (except for one case in which the response was weakly negative), verifying that the observed anti-predator responses were due to alarm cues.
DISCUSSION
We found strong evidence for trait co-dependence between escape responses and body size. It is easy to predict that the rela-tionship between body size and escape responses would be inversely related (trait compensation), because increased body size may make sturgeon less vulnerable to predators and allow them to reduce their investment in costly antipredator behaviour (Peterson et al., 2007). However, the relationship we found suggests that the magnitude of escape responses is mechanistically linked to body size. As sturgeon grow larger, their tailfin area increases, their accessory fins grow larger and their swimming musculature be-comes more highly developed, all of which allow sturgeon to show stronger and more sustained escape responses. Indeed, swimming endurance in lake sturgeon increases with body length (Peake, Beamish, McKinley, Scruton,&Katopodis, 1997) and, in a closely related species, the white sturgeon,Acipenser transmontanus, small juveniles (<82 mm tail length, TL) show lower escape speeds than medium (82e92 mm TL) and large (>93 mm TL) juveniles (Boysen &Hoover, 2009). Furthermore, our results suggesting that escape responses and body size are co-dependent are corroborated by what we know of the evolution of actinopterygian morphology. Specifically, high-power caudal propulsion is thought to have evolved due to selective pressure from pursuit predators (Webb, 1982). Trait co-dependence between escape behaviour and body size may extend across taxonomic boundaries as well. For example, larger (and older) hellbenders, Cryptobranchus alleganiensis, in-crease activity in response to predators (as an escape response), while smaller (and younger) individuals typically display predator avoidance behaviour instead (Crane&Mathis, 2013).
Cover-seeking behaviour showed a complex multitiered rela-tionship that was dependent on specific morphological combina-tions. Similar to our results for escape responses, body size was the
0.1571 N=40
0.07698 N=25
0.2612 N=8
0.031 N=12
0.1194 N=13 0.3244
N=7
0.1216 N=33 Length< 36.95
Length< 71.55
Mottled
Not mottled Length>= 36.95
Length>= 71.55
Figure 4.Regression tree showing the relationship between cover-seeking behaviour
(increased time in rocky microhabitats) relative to three morphological variables (fish
length, scute size, body pattern) based on 40 trials. Fish length and body pattern were the only two variables that covaried with cover-seeking behaviour. Values at each node indicate the mean proportion increase in time spent in rocky areas following exposure to alarm cues, where higher values indicate stronger cover-seeking behaviour. Values
along the upper branches indicate the thresholdfish length (mm) that define the
different behavioural groups. The lowermost branches indicate the presence or absence of mottled body pattern.
Table 2
Descriptive statistics for escape responses and cover-seeking antipredator behav-iours of young (lacking mottled body coloration) and older (mottled) lake sturgeon
Escape response
(mean±SE)
Cover seeking
(mean±SE)
Alarm cue
(N¼20)
Control
(N¼20)
Alarm cue
(N¼20)
Control
(N¼20)
Young, mottling absent
(42e69 dph)
1.96±0.92 0.12±0.13 0.19±0.04 0.11±0.07
Older, mottling present
(130e148 dph)
7.35±2.42 0.08±0.26 0.12±0.04 0.01±0.02
dph: days posthatch. Units are the relative change in behaviour (relative to the
prestimulus baseline) and were calculated using the formula ((postpre)/(pre)).
primary morphological trait linked to cover use, but in the opposite direction. The smallest fish in the study occupied rocky micro-habitats more than any other size class, indicating that occupying protective rocky microhabitats may compensate for small body size. This result matches our predicted hypothesis concerning the relationship between body size and cover-seeking behaviour. However, the largestfish in the study also increased the amount of time spent in rocky areas, although not to the same degree as the smallestfish. This might be evidence of trait co-specialization. The largestfish tested here may be able to manoeuvre though rocky microhabitats quickly and swiftly and be able to use rocky features to their advantage when faced with a threat, further increasing their probability of survival, but we presently have no data to support this hypothesis. Finally, sturgeon of intermediate size did not increase the amount of time they spent in rocky areas if they were mottled, whereas sturgeon that lacked this cryptic colour pattern did increase the amount of time spent in rocky areas. The former case may represent a form of trait compensation, allowing mottled sturgeon to remain in foraging grounds even in high-risk situations. The latter case may also represent a case of trait compensation, albeit of a different sort. Here, sturgeon that lacked the mottled body pattern appeared to compensate by seeking cover in rocky areas. In this case, selection appears to have favoured specific behaviours that are dependent on the presence or absence of cryptic coloration. This is not surprising given that several pre-vious studies have found that predators can impose correlational selection on prey colour pattern and behaviour (Brodie, 1989; Forsman&Appelqvist, 1998).
One very surprising outcome of this study was that scute size did not covary with sturgeon behaviour. Scutes are energetically costly to produce and increase energetic costs when swimming due to increased drag (Peterson et al., 2007). We therefore predicted that this cost would be offset by a reduction in antipredator behaviour (i.e. trait compensation). However, it may be that at this stage of development sturgeon scutes are not large enough or strong enough to provide an effective defence against predators, which would prevent behavioural changes in response to scute development from evolving in sturgeon during this life stage. As sturgeon mature and their scutes grow larger and stronger, any behavioural effects due to scutes may become evident. On the other hand, it may be that scutes simply co-specialize with antipredator behaviour in sturgeon to maximize survival. Further study is clearly needed to resolve this issue. In a related species, Gadomski and Parsley (2005)found that predation on white sturgeon by chan-nel catfish,Ictalurus punctatus, and northern pikeminnow, Ptycho-cheilus oregonensis, decreases as a function of sturgeon size, and is nearly absent once white sturgeon reach approximately 130 mm in length, while predation on alternative prey (Chinook salmon,
Oncorhynchus tshawytscha, or coho salmon,Oncorhynchus kisutch) of comparable size remains much higher. This suggests unique sturgeon morphology, such as scute development, influences pre-dation risk.
Together, our results suggest that behaviour may covary most strongly with body size, followed by cryptic coloration, which in turn covaries with behaviour more than defensive morphological structures (scutes) at this stage of development. In addition, we found that body size may be co-dependent and compensatory, and perhaps even co-specialized as well, across different behaviours and stages of morphogenesis. Indeed, selection acts on total or-ganism performance, and traits (and interactions between traits) need not be limited to a single mechanism so long as there is adaptive value to the decision rule directing and adjusting behaviour.
Evaluating the adaptive value of these behaviouremorphology combinations directly would require an empirical test of their
survival value in natural populations. However, all sturgeon species are now listed as endangered in part of their ranges, and most in all of their ranges (Billard & Lecointre, 2001; Birstein, Bemis, & Waldman, 1997). In addition, sturgeon are more critically endan-gered than any other animal family on the planet today (IUCN, 2010), precluding our ability to test survival value of behav-iouremorphology trait combinations at this time. If and when sturgeon populations recover, testing the adaptive value of these integrated traits is an obvious next step.
Frequency-dependent biotic interactions, such as preda-toreprey relationships, are often regarded as strong enough to produce and maintain adaptive genetic correlations amongfi tness-related traits (Sinervo&Svensson, 2002). As sturgeon morphology has been largely preserved since the lower Jurassic period (~200 million years ago), it may represent a highly successful morphological trait complex to which behavioural responses are
finely tuned. This study sheds light on how behaviour interacts with morphology in a dynamic multitrait system. Insight into the extent to which behaviour covaries with morphological develop-ment is important in order to understand the limitations or reaches of adaptive phenotypic plasticity and the evolution of integrated phenotypes. Continuing to understand the manner by which se-lection acts on integrated phenotypes should prove to be an exciting area of future research.
Acknowledgments
We thank the Natural Sciences and Engineering Research Council of Canada, the Canadian Wildlife Federation and the Uni-versity of Saskatchewan Department of Biology forfinancial sup-port. We thank J. Hunter and Sustainable Sturgeon Culture for providingfish used in this study and J. Sloychuk for help withfish care. We thank J. Johnstone for valuable statistical advice and two anonymous referees for advice on an earlier draft of this manu-script. All research herein was conducted under animal use protocol 20120013 issued by the University of Saskatchewan.
References
Abrahams, M. V. (1995). The interaction between antipredator behaviour and antipredator morphology: experiments with fathead minnows and brook sticklebacks.Canadian Journal of Zoology, 73, 2209e2215.
Baker, D. W., Peake, S. J., & Kieffer, J. D. (2008). The effect of capture, handling, and tagging on hematological variables in wild adult lake sturgeon.North American Journal of Fisheries Management, 28, 296e300.
Billard, R., & Lecointre, G. (2001). Biology and conservation of sturgeon and pad-dlefish.Reviews in Fish Biology and Fisheries, 10, 355e392.
Birstein, V. J., Bemis, W. E., & Waldman, J. R. (1997). The threatened status of Aci-penseriform species: a summary.Environmental Biology of Fishes, 48, 427e435.
Blumstein, D. T., Daniel, J. C., & Evans, C. S. (2006).JWatcher(Version 1.0)http://
www.jwatcher.ucla.edu/.
Boysen, K. A., & Hoover, J. J. (2009). Swimming performance of juvenile white sturgeon (Acipenser transmontanus): training and the probability of entrain-ment due to dredging.Journal of Applied Ichthyology, 25, 54e59.
Brodie, E. D., III (1989). Genetic correlations between morphology and antipredator behaviour in natural populations of the garter snakeThamnophis ordinoides. Nature, 342, 542e543.
Brodie, E. D., III (1992). Correlational selection for color pattern and antipredator behavior in the garter snakeThamnophis ordinoides.Evolution, 46, 1284e1298. Chivers, D. P., & Smith, R. J. F. (1998). Chemical alarm signaling in aquatic
preda-toreprey systems: a review and prospectus.Ecoscience, 5, 338e352. Chivers, D. P., Zhao, X., & Ferrari, M. C. O. (2007). Linking morphological and
behavioural defences: preyfish detect the morphology of conspecifics in the diet of their predators.Ethology, 113, 733e739.
Crane, A. L., & Mathis, A. (2013). Learning about danger by young hellbenders (Cryptobranchus alleganiensis): are antipredator strategies ontogenetically plastic?Amphibia-Reptilia, 34, 119e124.
De'ath, G., & Fabricius, K. E. (2000). Classification and regression trees: a powerful yet simple technique for ecological data analysis.Ecology, 81, 3178e3192. Dewitt, T. J., Sih, A., & Hucko, J. A. (1999). Trait compensation and cospecialization in
Ferrari, M. C. O., Wisenden, B. D., & Chivers, D. P. (2010). Chemical ecology of predatoreprey interactions in aquatic ecosystems: a review and prospectus. Canadian Journal of Zoology, 88, 698e724.
Forsman, A., & Appelqvist, S. (1998). Visual predators impose correlational selection on prey color pattern and behavior.Behavioral Ecology, 9, 409e413.
Gadomski, D. M., & Parsley, M. J. (2005). Laboratory studies on the vulnerability of young white sturgeon to predation.North American Journal of Fisheries Man-agement, 25, 667e674.
International Union for Conservation of Nature (IUCN). (2010).Sturgeon more
crit-ically endangered than any other group of species(18 March 2010, International
news release). Retrieved from:
http://www.iucn.org/?4928/Sturgeon-more-critically-endangered-than-any-other-group-of-species.
Jolliffe, I. (2005).Principal component analysis(2nd ed.). New York, NY: Springer. Legendre, P., & Legendre, L. (2012). Numerical ecology (3rd ed.). Oxford, U.K.:
Elsevier.
Lind, J., & Cresswell, W. (2005). Determining thefitness consequences of anti-predation behavior.Behavioral Ecology, 16, 945e956.
Mikolajewski, D. J., & Johansson, F. (2004). Morphological and behavioral defenses in dragonfly larvae: trait compensation and cospecialization.Behavioral Ecology, 15, 614e620.
Peake, S., Beamish, F. W. H., McKinley, R. S., Scruton, D. A., & Katopodis, C. (1997). Relating swimming performance of lake sturgeon, Acipenser fulvescens, to
fishway design. Canadian Journal of Fisheries and Aquatic Sciences, 54, 1361e1366.
Peterson, D. L., Vecsei, P., & Jennings, C. A. (2007). Ecology and biology of lake sturgeon, a synthesis of current knowledge of a threatened North American Acipenseridae.Reviews in Fish Biology and Fisheries, 17, 59e76.
Pettersson, L. B., Nilsson, P. A., & Br€onmark, C. (2000). Predator recognition and defense strategies in crucian carp,Carassius carassius.Oikos, 88, 200e212. Preisser, E. L., Bolnick, D. I., & Benard, M. F. (2005). Scared to death? The effects of
intimidation and consumption in predatoreprey interactions. Ecology, 86, 501e509.
R Development Core Team. (2012).R: A language and environment for statistical
computing. Vienna, Austria: R Foundation for Statistical Computing. http:// www.R-project.org.
Rundle, S. D., & Br€onmark, C. (2001). Inter- and intraspecific trait compensation of defence mechanisms in freshwater snails.Proceedings of the Royal Society B: Biological Sciences, 268, 1463e1468.
Schneider, C. A., Rasband, W. S., & Eliceiri, K. W. (2012). NIH Image to ImageJ: 25 years of image analysis.Nature Methods, 9, 671e675.
Sinervo, B., & Svensson, E. (2002). Correlational selection and the evolution of genomic architecture.Heredity, 89, 329e338.
Therneau, T., Atkinson, B., & Ripley, B. (2012).rpart: Recursive Partitioning(R package
Version 3.1e55)http://CRAN.R-project.org/package¼rpart.
Webb, P. W. (1982). Locomotor patterns in the evolution of actinopterygianfishes. American Zoologist, 22, 329e342.
Wisenden, B. D. (2008). Active space of chemical alarm cue in naturalfish pop-ulations.Behaviour, 145, 391e407.
Wishingrad, V., Chivers, D. P., & Ferrari, M. C. O. (2014). Relative cost/benefit
trade-off between cover-seeking and escape behaviour in an ancestral fish: the
importance of structural habitat heterogeneity.Ethology. Advance online