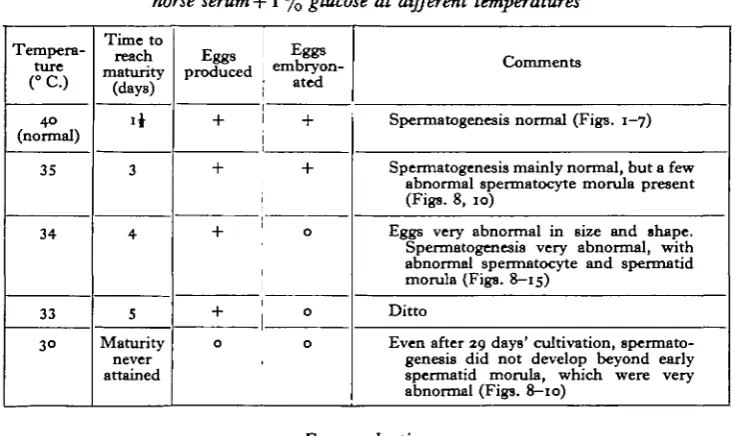
Studies on Tapeworm Physiology
6
0
0
Full text
Figure
Related documents
Full text
Figure
Related documents