SKELETAL STRAIN PATTERNS IN RELATION TO EXERCISE
TRAINING DURING GROWTH
ANDREW A. BIEWENER ANDJOHN E. A. BERTRAM*
Department of Organismal Biology and Anatomy, The University of Chicago, 1025 East 57th Street, Chicago, IL 60637, USA
Accepted 19 July 1993
Summary
Bones are believed to alter their shape in response to changes in tissue strains produced by physical activity and the goal of this study is to examine whether modeling responses of a growing bone to changes in physical exercise are adjusted to maintain a uniform distribution of functional strains. We test this idea by comparing in vivo strains recorded in the tibiotarsus of white leghorn chicks during ‘intensive’ treadmill exercise (60% of maximum speed, carrying a weight equal to 20% body weight on the trunk: 60%/L) with strains that had been recorded previously during ‘moderate’ treadmill exercise (35% of maximum speed, unloaded: 35%/UNL) at similar bone sites. Our hypothesis is that modeling adjustments of bones subjected to the intensive load-carrying exercise should re-establish strains recorded in the bones subjected to moderate exercise. At each exercise level, the animals were exercised for 5 days per week (2500 loading cycles per day) from 2 to 12 weeks of age. As in the moderate exercise group studied earlier, strains measured at six functionally equivalent sites on the tibiotarsus of the 60%/L group were consistently maintained during growth from 4 to 12 weeks of age. In addition, the pattern of strain recorded at these sites was uniformly maintained over the full range of speeds recorded (from 0.48 to 2.70 ms21at 12 weeks of age). Peak strains measured at 4 weeks of age in the load-carrying exercise group were initially elevated by 57% overall compared with peak strains recorded in the moderate exercise group. At 8 weeks of age, strain levels in the 60%/L group differed by only 4% overall compared with those recorded in the 35%/UNL group. The nature of strain (tensile versus compressive) and the orientation of principal strain at corresponding sites were also similar in the two groups. At 12 weeks of age, however, bone strain levels in the 60%/L group were again elevated (47% overall) compared with those recorded in the 35%/UNL group, although the general pattern and orientation of strains remained similar. This finding suggests a transient modeling response of the bone to the onset of exercise training, which was lost during subsequent growth, possibly because the normal pattern of functional strain was not altered significantly by the faster load-carrying exercise.
*Present address: Department of Anatomy, New York College of Veterinary Medicine, Cornell University, Ithaca, NY 14805, USA.
Introduction
It is commonly accepted that bones continually adjust their shape and mass in response to changes in mechanical loading patterns during an animal’s lifetime. The extent to which a bone can alter its form and the extent to which growing and mature bones have similar adaptive capacities, however, remain unclear (Bertram and Swartz, 1991; Biewener and Bertram, 1992). In addition, the precise structural objective of adaptive modeling and the mechanical stimuli that may initiate it are poorly understood. It seems likely that functional adaptation must depend on certain features of a bone’s loading history (Carter et al. 1981; Carter, 1984, 1987), being mediated in some fashion by the cyclic strains developed in the bone associated with its use (Hert et al. 1971; Lanyon, 1984; Rubin, 1984). Rubin and Lanyon (1984) have shown that surprisingly few (>36cyclesday21), high-magnitude (2000me, strain3106) loading cycles may be
sufficient to elicit a modeling response of a bone. Given that local tissue deformation is the direct, or indirect, agent of this response, it would seem likely that the functional integrity of each bone element in the skeleton could best be achieved and maintained by matching the bone’s form and mass to patterns of strain experienced during normal activity. The consistent pattern of surface midshaft strains recorded in the long bones of a number of species over a range of speed and change of gait supports this view (Lanyon and Baggott, 1976; Rubin and Lanyon, 1982; Biewener et al. 1983, 1988; Biewener and Taylor, 1986).
The purpose of this study is to test further the hypothesis that, in response to changes in physical activity, skeletal modeling maintains a uniform distribution of strains at functionally equivalent sites on a growing bone. We found previously (Biewener
et al. 1986) that the in vivo pattern of surface strains in the chick tibiotarsus engendered
by moderate exercise remained unchanged as the animals grew from 4 to 17 weeks of age (corresponding to a 10-fold increase in mass and a 3.3-fold increase in length of the bone). Though differing among sites, the magnitude, nature (tensile versus compressive) and orientation of principal strains at each site remained largely unaltered during this period of growth. These findings are consistent with those of Lanyon and his colleagues involving studies of older pigs (Goodship et al. 1979) and skeletally mature sheep (Lanyon et al. 1982), in which experimental removal of the ulnar diaphysis stimulated adaptive modeling of the radius to compensate for loss of the ulna in each species. Initially elevated strains at the radial diaphysis returned to normal levels within a 3-month period in the juvenile pigs and within a 1-year period in the mature sheep. A recent study of the tarsometatarsus in skeletally mature roosters (Loitz and Zernicke, 1992), however, suggests that adaptation of mature bone may not be as sensitive to exercise-induced changes in peak strain as that observed in younger, growing bone.
achieved this both by increasing the speed of exercise and by having the animals carry loads proportional to their weight on their backs. While increases in running speed and body weight (1.2 times in the present case) each contribute to an overall increase in the magnitude of load that a bone must support, neither is likely to alter significantly the general distribution of strain within a bone (Rubin and Lanyon, 1982; Biewener et al. 1983; Biewener and Taylor, 1986). Consequently, the increase in exercise intensity achieved using this approach was designed to increase strain magnitude without disrupting the normal distribution of strain in the bone. We reasoned that elevated strain levels produced by the increase in exercise intensity would elicit an adaptive modeling response, beyond that of normal bone growth, to return strain levels near to those recorded previously in animals that had experienced only moderate treadmill exercise while growing. A non-invasive exercise approach such as this has the advantage of subjecting the bone to an elevated, but physiologically normal, strain pattern. Measurements of in vivo strain on the growing chick tibiotarsus during heavy exercise are reported here and compared with measurements of bone strain made previously in the moderate exercise group at the same functional sites. Changes in bone cortical geometry and shape elicited by the exercise regimens are reported elsewhere (Biewener and Bertram, 1993).
Materials and methods
The training and experimental protocols, the surgical methods and bone strain recording procedures used in the present experiments and described below are similar to those used in Biewener et al. (1986). Because growth slows considerably from 12 to 17 weeks of age, we limited our comparison of in vivo strain recordings (and associated modeling responses of the bone) to chicks aged 4–12 weeks.
Animals and exercise training
Fifteen male white leghorn chicks were divided randomly into three exercise groups (five animals per group) and exercised at 60% of their maximum running speed, carrying an additional 20% of their body weight (60%/L) in weights distributed by a cloth cummerbund about their trunk. By exercising the animals at a percentage of their maximum speed, the animals maintained a uniform exercise level as they grew to a larger size. At all ages, exercise speed (0.90 ms21at 4 weeks; 1.29ms21at 8 weeks; 1.54ms21
at 12 weeks) corresponded to a constant stride frequency of 2.96±0.13strides s21(N=15).
Load-carrying was used to increase further the magnitude of bone loading. The weights that the animals carried were adjusted every few days to account for increases in body weight as the animals grew. The weight-packs were designed to fit over the cloth harness that the animals wore later during the recordings of in vivo strain.
observed earlier in the moderate exercise, non-load-carrying group (35%/UNL; Biewener et al. 1986). As in our earlier study, exercise duration was set at 15min corresponding to about 2500 loading cycles per day at the faster speed. At all other times, the animals’ activity was restricted to sedentary weight support by housing them in confined conditions in 0.5 m30.5m cages (e.g. three animals weighing 1.0kg each were housed per cage at 12 weeks of age).
Surgery and in vivo bone strain recordings
[image:4.612.139.323.371.590.2]Each group of five animals was exercised to 4, 8 or 12 weeks of age. At each of these ages, three animals (the other animals were used for structural and histological analyses of bone modeling changes; Biewener and Bertram, 1993) underwent aseptic surgery to attach strain gauges on the proximal-medial (PM), medial-midshaft (MM), cranial-midshaft (CRM) and caudal-cranial-midshaft (CAM) cortices of the left tibiotarsus and on the cranial-distal (CRD) and caudal-distal (CAD) cortices of the right tibiotarsus (Fig. 1). Only one gauge could be attached medially at the bone’s proximal level because of attachments of anterior and posterior muscles and the fibula laterally. All bone sites were surgically exposed from the medial side of the limb. At each site, the overlying musculature was retracted away from the bone and a small patch (6mm36 mm) of the periosteum was cut out and removed using a scalpel. The underlying mineralized surface of the bone was then lightly scraped with a small periosteal elevator, defatted and dried by applying ether to the bone surface using a cotton-tipped applicator. After ensuring a dry,
clean surface, the strain gauge was bonded with a self-catalyzing cyanoacrylate adhesive with pressure applied to the gauge for 90s. The strain recording sites were defined with respect to a fixed percentage of the bone’s length (proximal 30%, midshaft 50%, distal 70%) and were considered to be functionally equivalent locations on the bone, consistent with the constant linear relationship of muscle and ligamentous attachment sites to a bone during its growth (Grant et al. 1980).
The lead wires from the strain gauges (36 gauge, etched Teflon-insulated: Micromeasurements) were sutured to surrounding fascia for strain relief and passed subcutaneously to an opening in the animal’s skin situated over its back, between the wings. The lead wires were then soldered to a multipin plastic connector (Amphenol, 222 series) and all wounds were sutured close. The connector plug was securely wrapped with an elastic bandage (Vet-Wrap) and held in place within a cloth cummerbund wrapped over the animal’s back and under its wings.
Because of the small size of the bone at 4 weeks of age, strain recordings were made using single-element strain gauges (type FLE-05, Tokyo Sokki Kenkyujo). These strain recordings were made with the gauges aligned along the bone’s longitudinal axis. At 8 and 12 weeks of age, strain recordings were made using rectangular rosette strain gauges (type FRA-1) at three of the bone sites in each animal. Single-element gauges were used at the other three sites. Sites of rosette strain recordings were alternated to determine the orientation and magnitude of principal strains in four of the six sites overall (excluding the MM and CAD sites). The longitudinal strains recorded at the proximal-medial (PM), cranial-midshaft (CRM), caudal-midshaft (CAM) and cranial-distal (CRD) sites were used to estimate the maximum (absolute magnitude) principal strains at these sites at 4 weeks of age, based on the axis of maximum principal strain determined from rosette strain gauge recordings made at 8 and 12 weeks of age. This procedure assumes a uniaxial planar state of strain at the bone’s surface to construct a Mohr’s circle of strain at each site (Dally and Riley, 1978) based on the magnitude of shear strain determined from the rosette strain recordings at the later ages. The principal strains are then computed on the basis of the magnitude of the longitudinal strain recorded from the single-element gauge made at 4 weeks of age and its angle to the assumed principal strain axes. Estimates of maximum principal strain determined from longitudinal strains measured at 4 weeks of age averaged +18% at the PM site (corresponding to a mean 22˚ deviation from the principal strain axis, see Table 2), +48% at the CRM site (mean 39˚ deviation), +28% at the CAM site (mean 30˚ deviation) and +23% at the CRD site (mean 26˚ deviation). Because this procedure oversimplifies the true state of strain by assuming a uniaxial planar state of strain at the bone’s surface, interpretations based on comparisons of corrected single-element strain recordings at 4 weeks of age should be treated with some caution.
during the swing phase of the limb. These levels were consistent with those recorded while the animal was resting with its limb held in a relaxed state. Following the bone strain recordings, the animals were immediately killed and their bones removed for structural analyses of bone modeling changes (Biewener and Bertram, 1993).
Least-squares regressions of the data (based on mean maximum strain levels within each individual animal at each site) were carried out to test for significant shifts in strain magnitude versus age at each site. Additional comparisons of strain magnitude and orientation between groups were performed using Student’s t-tests.
Results
Strain pattern versus growth
Strains recorded at each of the six sites when the animals ran at 60% of their maximum speed, carrying an additional 20% body weight load, increased slightly with age (Fig. 2). At no site did the magnitude of strain decrease at 8 or 12 weeks compared with levels measured at 4 weeks of age. Only at the CAM site was the increase in strain with age significant (Table 1). At two sites (MM and CRD), no change in strain magnitude was evident. At all sites, the nature (tensile versus compressive) of strain remained unchanged. Consequently, the overall pattern of locomotor strains recorded at the six sites changed very little from 4 to 12 weeks of age. Though more variable, the distribution of strain recorded in the intensive, load-carrying exercise animals was similar to that observed previously in animals exercised at 35% of their maximum speed, unloaded (35%/UNL; Biewener et al. 1986).
[image:6.612.58.433.514.600.2]In addition to the uniform distribution of strain magnitude and sign, the orientation of principal strains recorded at three of four (rosette strain gauge) sites also remained similar among animals at 8 and 12 weeks of age (Table 2). Although the orientation of principal strain differed significantly at the proximal-medial (PM) site, rosette strain recordings were obtained from only one animal at each of these ages, rendering this finding of questionable significance. In no case did carrying a load substantially alter the orientation of peak principal strain compared with when the same animals ran unloaded (Table 2).
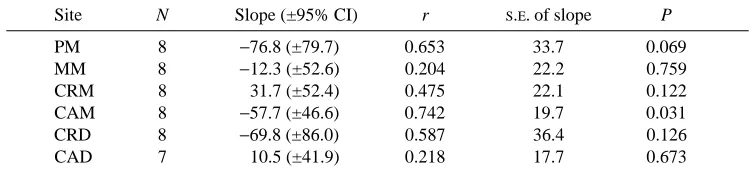
Table 1. Least-squares regression statistics for maximum principal strain versus age
(weeks) at six recording sites on the tibiotarsus
Site N Slope (±95% CI) r S.E. of slope P
PM 8 −76.8 (±79.7) 0.653 33.7 0.069
MM 8 −12.3 (±52.6) 0.204 22.2 0.759
CRM 8 31.7 (±52.4) 0.475 22.1 0.122
CAM 8 −57.7 (±46.6) 0.742 19.7 0.031
CRD 8 −69.8 (±86.0) 0.587 36.4 0.126
CAD 7 10.5 (±41.9) 0.218 17.7 0.673
For all but the CAM site, the slope of the regression is not significantly different from zero, indicating no age-related change in the magnitude or nature of strain at each of the other sites.
The orientation of principal strain determined at each site was also generally consistent with that observed previously in the moderate exercise group (Biewener et al. 1986). Angles of principal strain relative to the bone’s longitudinal axis at the midshaft (CRM 246˚ and CAM +35˚ at 12 weeks) indicate a significant component of torsional loading during the support phase of the stride.
Load-carrying and strain magnitude
Carrying a load equal to 20% of their body weight resulted in an overall 15% increase in strain magnitude at 8 and 12 weeks of age, compared with when the animals ran unloaded at the same speed (Table 3). The increase was variable among sites, however, ranging at 8 weeks from as high as +32% at the CRD site to as low as +5% at the MM site. Because strain levels recorded at the CAD site were low under all exercise conditions, the potential for error associated with establishing a zero strain level (when balancing the bridge circuits, see Materials and methods) relative to the peak strain recorded is high. Consequently, this site was excluded from comparisons between the two exercise groups and loaded versus unloaded exercise conditions. (The possibility that the low compressive strains recorded at the CAD site were due to gauge loosening was excluded by direct post-mortem loading of the bone to produce artificially high strains at this site.)
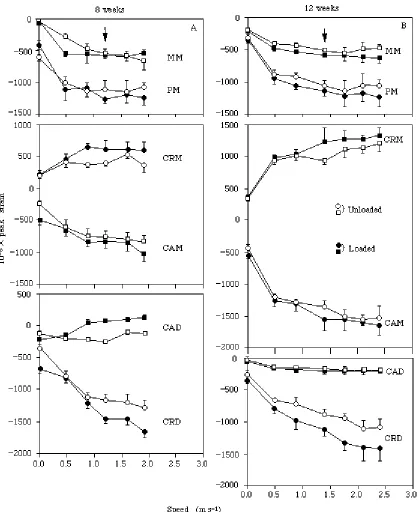
Bone strain patterns versus speed
[image:8.612.64.432.130.257.2]Although the pooled data show no significant differences in strain magnitude associated with load-carrying at the exercise speed, differences in strain magnitude were generally significant when compared at a given speed within a given animal (Fig. 3). As a result, the elevation of strain magnitude associated with load-carrying at most sites was consistent over each animal’s measured range of running speed (Fig. 3). At no site Table 2. Orientation (u) of maximum principal strain versus age at those sites for which
rosette strain recordings were made during loaded versus unloaded exercise at 60% maximum speed (positive denotes a proximo-medial orientation for the CRM and CRD
sites, proximo-lateral for the CAM site and proximo-caudal for the PM site) u(degrees)
Age PM CRM CAM CAD
8 weeks
Unloaded +30±5 −30±13 +25±3 −28±15
Loaded +33±7 −33±15 +24±5 −30±16
(1, 11) (2, 19) (2, 19) (3, 28)
12 weeks
Unloaded +9±3 −40+3 +33±5 −23±10
Loaded +10±3 −46±4 +35±7 −21±9
(1, 8) (2, 17) (2, 17) (3, 26)
Values represent the mean ± S.D.
(excluding the CAD site) did the tensile or compressive nature of strain change as a result of carrying a load. Overall, the pattern of locomotor strain recorded at the six sites on the tibiotarsus remained remarkably consistent as the animals increased speed, both when they were unloaded and when they carried a load.
Strain magnitude versus age and exercise level
[image:10.612.61.433.104.256.2]At 4 weeks of age, peak strains recorded at the intensive exercise level (60%/L) were initially 57±14% greater than those recorded when the same three animals ran at the moderate exercise level (35%/UNL) (Fig. 4A and Table 4). Except for the CAD site, the Table 3. Peak principal strains (meor ×106) recorded when the animals ran unloaded
versus loaded at 60% of their maximum running speed
Bone recording sites
Age PM MM CRM CAM CRD CAD
8 weeks
60%/unloaded −654±342 −838±174 +511±87 −954±230 −1206±294 −204±53 60%/loaded −763±367 −877±196 +589±153 −1044±261 −1593±301 −128±83
Ratio 1.17 1.05 1.15 1.09 1.32 0.63
(mean) (1.16)†
12 weeks
60%/unloaded −1113±228 −860±273 +884±116 −1251±166 −1217±279 −181±38 60%/loaded −1245±230 −926±303 +987±250 −1430±168 −1442±298 −203±38
Ratio 1.12 1.08 1.12 1.14 1.18 1.12
(mean) (1.13)†
†Values for the CAD site were excluded in calculating an overall mean strain ratio, as the potential for error in establishing the percentage change in strain at this site is high owing to the error inherent in establishing a zero strain level during the limb cycle relative to the low strains recorded at this site.
Table 4. Strains (meor ×106) recorded at six sites on the tibiotarsus of three chicks at 4 weeks of age while exercising at the moderate (35%/UNL) and intensive load-carrying
(60%/L) exercise conditions
Bone sites
PM MM CRM CAM CRD CAD Mean
35%/UNL −410±201 −522±69 +465±95 −612±64 −472±52 −142±97 60%/L −606±248 −931±136 +685±152 −895±134 −774±96 −148±42
Ratio 1.48 1.78 1.47 1.46 1.64 1.04† 1.57
Values are the pooled means ± S.D. (total number of strides >24).
The ratio indicates the initial strain differential established at each site by the load-carrying exercise regimen compared with the moderate exercise regimen.
[image:10.612.60.432.505.580.2]difference in strain magnitude was fairly consistent among sites. This 57% increase in strain magnitude provides an estimate of the mechanical stimulus that the growing tibiotarsi were exposed to at the onset of the 60%/L exercise training, assuming that the adaptive response of the bone was minimal at 4 weeks of age. Such an increase, however, is well within the normal physiological loading range of the bone, in which increases in strain magnitude of 389±49% from rest and 63±24% from the slowest walking speed (0.48 ms21 at 12 weeks) to the fastest running speed (2.70 ms21 at 12 weeks) were
observed. These percentage increases in strain magnitude represent the average increases obtained at all recording sites (excluding the CAD site) for all animals while running unloaded (Fig. 3).
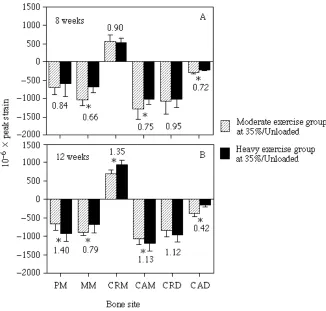
Following the initial increase in strain associated with the intensive exercise regime measured at 4 weeks, continued exercise to 8 weeks of age resulted in a substantial decrease in the difference of strain measured between the two exercise groups (Fig. 4B). At 8 weeks of age, strains in the 60%/L group averaged only 4% greater than strains recorded previously at the same sites in the 35%/UNL group. At two of the sites (MM and CAM), strain levels were actually lower in the 60%/L group than strains recorded at the same sites in the 35%/UNL group. In contrast, peak compressive strains recorded at the CRD site remained elevated in the 60%/L group. This was due in part to increased bending of the bone at its distal level during the intensive, load-carrying exercise compared with when the animals ran without carrying a load.
At 12 weeks of age (Fig. 4C), differences in strain level between the intensive and moderate exercise groups were greater at all sites compared with the differences established at 8 weeks, with an average differential of 47%, similar to that observed originally at 4 weeks of age. Once again, individual bone sites exhibited different levels of change.
A second comparison that tests whether the intensive, load-carrying exercise had an effect on the bone’s growth in relation to functional patterns of strain is shown in Fig. 5. If the 57% increase in strain measured at 4 weeks of age significantly affected the bone’s subsequent structural modeling response, as originally hypothesized, bone strains recorded in the 60%/L group would be expected to be significantly reduced compared with strains recorded in the 35%/UNL group when compared at the same exercise level (i.e. when both groups of animals were walking at 35% of their maximum speed, carrying no load). Fig. 5 shows that at 8 weeks of age this was the case, with strains in the heavy exercise group averaging 83±11% of those acting in the moderate exercise group (excluding the CAD site). At 12 weeks, however, strain magnitude was greater in the heavy exercise group at four of the same five sites. This change at the 35%/UNL exercise level matches the increased differential in strain observed between the two groups from 8 to 12 weeks when compared at their respective exercise levels (compare Fig. 4B,C with Fig. 5A,B).
Discussion
functional strain at equivalent sites within a growing bone. This hypothesis was based on an earlier study of strain patterns measured during moderate exercise (fast walk) in the chick tibiotarsus, in which we found that the distribution of locomotor strains was uniformly maintained from 4 to 17 weeks of age (Biewener et al. 1986). With an increase in exercise intensity, we anticipated that the structural properties of the bone’s cortex would be enhanced to achieve a distribution of functional strains similar to that measured in the moderately exercised animals. Although the initial 57% elevation of strain magnitude engendered by the faster load-carrying exercise at 4 weeks of age was abolished at 8 weeks of age, strains measured in the 60%/L group were again significantly higher than those recorded in the 35%/UNL group at 12 weeks of age. Despite these changes in strain magnitude, the relative magnitude and nature of strain
recorded in the strenuous exercise group were consistently maintained at both 8 and 12 weeks of age, being similar to the distribution of strain observed in the earlier moderate exercise group. Functional strain patterns recorded on the tibiotarsus were also consistent over a wide range of speed and gait. While this has been observed in the long bones of other species moving at different speeds and with different gaits (Rubin and Lanyon, 1982; Biewener et al. 1983; Biewener and Taylor, 1986), these observations have been limited to strain measurements at only two or three midshaft sites on a bone. Hence, our strain measurements at six sites on the chick tibiotarsus provide the strongest support yet for functional strain patterns being uniformly maintained over a range of locomotor activity.
The observed initial increase, re-establishment and subsequent increase in strain levels recorded in the load-carrying group compared with the moderate exercise group at 4, 8 and 12 weeks of age indicate a transient response of the strenuously exercised tibiotarsi to the onset of exercise and elevated loading that is consistent with changes in cortical geometry observed in these animals: cortical area (A) and second moment of area (I) were elevated by 26% and 40% compared with sedentary controls at 8 weeks of age, but were substantially diminished at 12 weeks of age (A 9 %, I 12%; Biewener and Bertram, 1993). The reason for the inconsistent nature of these changes in strain level and cortical bone geometry is unclear. A recent study of strains developed in the tarsometatarsus of skeletally mature chickens during treadmill exercise (Loitz and Zernicke, 1992) also reports unexpected changes in strain levels associated with exercise compared with values from sedentary control animals. Lanyon (1987) has suggested that bone remodeling is an error-driven process in which unusual strain conditions, even if infrequent, are most critical in eliciting an adaptive response. While this suggestion may explain the initial re-establishment of strains we observed between the two groups at 8 weeks of age following the onset of exercise and elevated functional strain, it is difficult to interpret the subsequent increase in strain as being due to a diminished growth stimulus of the faster, load-carrying exercise, because it became the more ‘usual’ loading regimen. Our hypothesis of adaptive modeling of growing bone in relation to functional strain pattern originates from a similar hypothesis proposed by Lanyon and his colleagues (Goodship et al. 1979; Lanyon et al. 1982) for young adult and mature bone. In their studies of pigs and sheep, experimental strain overload of the radius produced by surgical resection of the ulnar diaphysis elicited an adaptive modeling response of the radial diaphysis in each species, increasing cortical area and altering the shape of the bone’s cortex to re-establish ‘customary’ strains associated with walking. Skeletal adjustments of functional strain were achieved within 3 months in the young adult pigs and within 1 year in the older sheep. Although strains were not recorded at intervals during the adaptive process, bone modeling appeared to proceed progressively with an initial rapid periosteal deposition of woven bone that was subsequently consolidated at a slower rate into a dense compacta. No evidence of a transient response or a reversal in the remodeling process was observed.
et al. 1986). In response to long-term immoblization, Uhthoff and Jaworski (1978)
observed biphasic and dissimilar modeling responses in the long bones of dogs, in which resorption occurred mainly on trabecular and periosteal surfaces in young adult dogs but almost exclusively on the endosteal surface of older dogs. In a treadmill exercise study similar to our own, Matsuda et al. (1986) found that cortical growth was inhibited in the chick tarsometatarsus at 8 weeks of age following 5 weeks of exercise at 70% of maximum aerobic capacity, but was significantly enhanced when measured at 12 weeks of age. These morphological studies and our present in vivo strain study indicate, therefore, that growing and adult skeletons may respond differently to mechanical stimuli at different stages of growth and aging. To determine whether adaptive modeling is developmentally constrained, particularly in a rapidly growing bone such as the chick tibiotarsus, requires that parallel mechanical loading or exercise studies of mature versus growing bone be carried out in the same species.
Another possible explanation for the transient nature of the response that we observe in the present study is that the elevated strains produced by the load-carrying exercise were insufficient, either in intensity or duration, to maintain an adaptive response of the tibiotarsus during growth. Frost (1983, 1987) has proposed a strain stimulus threshold in the range of 1500–3000mefor bone tissue. Only when strain levels exceed this threshold would cells be signaled to initiate adaptive modeling of the bone. Consequently, as no site on the chick tibiotarsus experienced strain levels above 2000me (with most sites being below 1500me), adaptive modeling would not be expected to occur. This seems unlikely for three reasons. First, the 57% increase in strain achieved in the present experiments is greater than the magnitude of ‘strain overload’ achieved in the ulnar osteotomy study of the sheep radius (Lanyon et al. 1982) and comparable to that observed in the study of growing pigs (Goodship et al. 1979). In both experiments, significant adaptive modeling occurred to restore functional strain levels to near normal, even though the change in strain was well within the normal physiological range of each bone. Second, as we note above, cortical bone growth was significantly enhanced at 8 weeks of age in the 60%/L exercise group (Biewener and Bertram, 1993), correlating with the match in strain magnitude that we found between the 60%/L and 35%/UNL groups at this age. Finally, although the duration of exercise in our study was brief, the number of loading cycles experienced by the bone during this period (roughly 2500 cycles) is well beyond a threshold (36 cycles) shown by Rubin and Lanyon (1984) to be sufficient to elicit adaptive modeling in rooster ulnae. Nonetheless, the influence of additional (though low-magnitude) loading events associated with the transport and housing of the animals that might diminish the effect of the treadmill exercise cannot be ruled out.
percentage of bone length and cortical quadrant, allows us to demonstrate a consistently maintained pattern of strain during several weeks of growth, small differences in the location of strain recordings among animals can introduce significant variation in the strain established at a given site. Although we attempted to minimize this source of variation, it is unavoidable that this contributed to the overall variation that we found between the two groups. Consequently, our ability to assess exercise-induced skeletal adaptation in the context of changes in functional strain pattern based on in vivo surface strain measurements is limited to a certain extent by these factors.
Having noted these caveats, we believe that an attractive explanation for our results remains: adaptive modeling may be most sensitive to disruption of the normal
distribution or pattern of functional strain within a bone, rather than to an elevation of
strain magnitude per se . An important difference in the nature of the strain ‘overload’ produced in the experiments of Lanyon et al. (1982) and Goodship et al. (1979) compared with our present treadmill exercise study of the growing chick tibiotarsus is that, in terms of the relative magnitude, orientation and nature of strain recorded among the differing bone sites, the overall distribution of strain in the 60%/L exercise group was quite similar to that previously observed in the 35%/UNL group. In the experiments noted above, the normal distribution of locomotor strain within the radial midshaft was disrupted in both the pigs and sheep when the ulnar diaphysis had been removed. The possibility that disruption of a bone’s functional strain distribution underlies observed modeling responses has also been noted recently in controlled loading studies of the radius in roosters (Gross and Rubin, 1992).
It is interesting to note that at the one site (CRD) in which strains remained consistently elevated (from 49 to 70%) at all three ages examined, disruption of the normal (unloaded) strain gradient across the distal cortex was greatest (Table 5). Correspondingly, the most pronounced increase in periosteal bone formation occurred at the distal level of the bone at 8 weeks of age (Biewener and Bertram, 1992). In contrast, differences in bending-induced strain relative to axially compressive strain (eb:ea) between the two groups were
considerably less at the midshaft, where adaptive modeling was less extensive. Notably, the largest change in midshaft eb:ea occurred at 8 weeks, when the geometry of the
[image:16.612.59.435.539.626.2]midshaft cortex also exhibited the greatest response to increased mechanical loading
Table 5. Distribution of strain due to bending (eb) versus axial compression (ea) at the
midshaft and distal levels of the tibiotarsus for the moderate (35%/UNL) and intensive load-carrying (60%/L) exercise conditions
eb:ea
Age Midshaft Distal
(weeks) 35%/UNL 60%/L D% 35 %/UNL 60%/L D%
4 7.33 7.52 3 0.54 0.86 59
8 2.58 4.23 64 0.56 1.08 93
12 4.75 5.45 15 0.40 1.10 175
(Biewener and Bertram, 1993). In their study of rooster ulnae subjected to controlled loading, Rubin and Lanyon (1985) have argued that sufficiently large strains are also critical to the maintenance of skeletal mass. However, their artificially imposed loading regime resulted in a diaphyseal cortical strain distribution that differed considerably from that observed during normal wingbeat activity. Hence, the observed adjustments of skeletal mass were probably affected by changes in both strain magnitude and strain distribution, as Gross and Rubin (1992) have noted recently.
The fact that in vivo strain levels can vary considerably, but in a regular pattern, among differing sites within a bone suggests that a uniform strain threshold does not exist for all regions of a bone. It would seem that the threshold for adaptive modeling, if it is sensitive to strain magnitude, must vary among sites within the bone and, thus, is more likely to involve an assessment of the distribution of strain throughout a larger volume of the bone. In this context, remodeling algorithms involving site-specific strain thresholds in relation to measured or estimated functional strains have been adopted in an attempt to model the adaptive response of bone to mechanical stimuli with some limited success (Cowin et al. 1985). The observation that normal functional strain distributions at a given cortical level (in most cases measured at the bone’s midshaft) are maintained consistently over a range of speed and change of gait (Rubin and Lanyon, 1982; Biewener et al. 1983, 1988; Biewener and Taylor, 1986; and see Fig. 3) also supports the view that the distribution of strain within the bone is a critical component underlying adaptive bone modeling.
Implicit in our use of in vivo strain gauge recordings to ‘map’ the functional distribution of strain within a bone and to characterize its loading history for a given exercise regimen is the assumption that a uniaxial (planar) state of strain exists at each of the recording sites on the bone’s surface. However, given that the actual state of strain within a local region of the bone (both at the bone’s surface and deeper within its cortex) is the result of multiaxial loading, the objective of strain-mediated bone modeling is more likely to involve a three-dimensional assessment of strain at a local cellular level. In addition, by focusing on peak strain magnitude, we ignore the time-varying nature of functional loading patterns. Using two-dimensional finite element models, Carter and his colleagues (Carter, 1987; Carter et al. 1987; Carter and Wong, 1988) have shown that the decomposition of strain energy density into hydrostratic versus shear strain densities predicts well the spatial and temporal ossification patterns within a fetal bone, in which regions of high shear strain stimulate the differentiation of cartilage into bone. In that strain energy density can be generalized throughout the whole of a bone (in constrast to the site-specific nature of strain magnitude), it is an attractive candidate for the load-related variable to which bone cells respond. A recent study that attempted to correlate patterns of strain energy density with a bone’s modeling response, however, found the correlation to be weak (Brown et al. 1990).
over a range of speed and change of gait, as well as during growth, would suggest that functional adaptation is unlikely to be a major factor affecting bone mass and shape under most circumstances of normal activity during an animal’s lifetime.
We thank M. Lee and P. Ricketts for their assistance in these experiments. This work was supported by grants awarded to AAB by the Whitaker Foundation (grant no. 635214) and the NIH (AR 39828).
References
BERTRAM, J. E. A. ANDSWARTZ, S. M. (1991). The ‘Law of Bone Transformation’: a case of crying Wolff? Biol. Rev. 66, 245–273.
BIEWENER, A. A. ANDBERTRAM, J. E. A.(1992). Mechanical loading and bone growth in vivo. In Bone: A Treatise, vol. 7 (ed. B. K. Hall), pp. 1–36. Boca Raton: CRC Press.
BIEWENER, A. A. ANDBERTRAM, J. E. A. (1993). Structural of growing bone to exercise and disuse. J. appl. Physiol. (in press).
BIEWENER, A. A., SWARTZ, S. M. ANDBERTRAM, J. E. A.(1986). Bone modeling during growth: dynamic strain equilibrium in the chick tibiotarsus. Calc. Tiss. Int. 39, 390–395.
BIEWENER, A. A. ANDTAYLOR, C. R. (1986). Bone strain: a determinant of gait and speed? J. exp. Biol. 123, 383–400.
BIEWENER, A. A., THOMASON, J. ANDLANYON, L. E.(1983). Mechanics of locomotion and jumping in the forelimb of the horse ( Equus): in vivo stress developed in the radius and metacarpus. J. Zool., Lond. 201, 67–82.
BIEWENER, A. A., THOMASON, J. ANDLANYON, L. E.(1988). Mechanics of locomotion and jumping in the horse (Equus): in vivo stress in the tibia and metatarsus. J. Zool., Lond. 214, 547–565.
BROWN, T. D., PEDERSEN, D. R., GRAY, M. L., BRAND, R. A. AND RUBIN, C. T. (1990). Toward identification of mechanical parameters initiating periosteal remodeling: a combined experimental and analytic approach. J. Biomech. 23, 893–905.
CARTER, D. R. (1984). Mechanical loading histories and cortical bone remodeling. Calc. Tiss. Int. 36, S19–S24.
CARTER, D. R.(1987). Mechanical loading history and skeletal biology. J. Biomech. 20, 1095–1107. CARTER, D. R., HARRIS, W. H., VASU, R. ANDCALER, W. E. (1981). The mechanical and biological
response of cortical bone to in vivo strain histories. In Mechanical Properties of Bone (ed. S. C. Cowin). ASME Pub. AMD. 45, 81–92.
CARTER, D. R., ORR, T. E., FYHRIE, D. P. ANDSCHURMAN, D. J.(1987). Influence of mechanical stress on prenatal and postnatal skeletal development. Clin. Orthop. 219, 237–250.
CARTER, D. R. ANDWONG, M. (1988). The role of mechanical loading histories in the development of diarthrodial joints. J. orthop. Res. 6, 804–816.
COWIN, S. C., HART, R. T., BABER, J. R. ANDKOHN, D. H. (1985). Functional adaptation in long bones: establishing in vivo values for surface remodeling rate coefficients. J. Biomech. 18, 665–684. DALLY, J. ANDRILEY, W. F. (1978). Experimental Stress Analysis. 2nd edn. New York: McGraw-Hill. FROST, H. M. (1983). The minimum effective strain. A determinant of bone architecture. Clin. orthop.
rel. Res. 175, 286–292.
FROST, H. M.(1987). The mechanostat: a proposed pathogenic mechanism of osteoporoses and the bone mass effects of mechanical and nonmechanical agents. Bone and Mineral 2, 73–85.
GOODSHIP, A. E., LANYON, L. E. ANDMACFIE, H. (1979). Functional adaptation of bone to increased stress. J. Bone Jt Surg. 61A, 539–546.
GRANT, P. G., BUSHANG, P. H., DROLET, D. W. ANDPICKERELL, C. (1980). Invariance of the relative positions of structures attached to long bones during growth: cross-sectional and longitudinal studies. Acta. anat. 107, 26–34.
GROSS, T. S. ANDRUBIN, C. T. (1992). The correlation of bone tissue adaptation to specific mechanical stimuli. Trans. orthop. Res. Soc. 17, 96A.
JAWORSKI, Z. F. G. ANDUHTHOFF, H. K. (1986). Disuse osteoporosis: current status and problems. In Current Concepts of Bone Fragility (ed. H. K. Uhthoff and E. Stahl), pp. 181–194. New York: Springer-Verlag.
KIISKINEN, A. (1977). Physical training and connective tissues in young mice – physical properties of Achilles tendons and long bones. Growth 41, 123–137.
LANYON, L. E. (1984). Functional strain as a determinant of bone remodeling. Calc. Tiss. Int. 36, S56–S61.
LANYON, L. E. (1987). Functional strain in bone tissue. J. Biomech. 20, 1083–1093.
LANYON, L. E. ANDBAGGOTT, D. G. (1976). Mechanical function as an influence on the structure and form of bone. J. Bone Jt Surg. 58B, 436–443.
LANYON, L. E., GOODSHIP, A. E., PYE, C. J. ANDMACFIE, H. (1982). Mechanically adaptive bone remodelling. J. Biomech. 15, 141–154.
LOITZ, B. J. AND ZERNICKE, R. F. (1992). Strenuous exercise-induced remodeling of mature bone: relationships between in vivo strains and bone mechanics. J. exp. Biol. 170, 1–18.
MATSUDA, J. J., ZERNICKE, R. F., VAILAS, A. C., PEDRINI, V. A., PEDRINI-MILLE, A. ANDMAYNARD, J. A. (1986). Structural and mechanical adaptation of immature bone to strenuous exercise. J. appl. Physiol. 60, 2028–2034.
RUBIN, C. T. (1984). Skeletal strain and the functional significance of bone architecture. Calc. Tiss. Int. 36, S11–S18.
RUBIN, C. T. ANDLANYON, L. E. (1982). Limb mechanics as a function of speed and gait: a study of functional strains in the radius and tibia of horse and dog. J. exp. Biol. 101, 187–211.
RUBIN, C. T. AND LANYON, L. E. (1984). Regulation of bone formation by applied dynamic loads. J. Bone Jt Surg. 66A, 308–314.
RUBIN, C. T. ANDLANYON, L. E.(1985). Regulation of bone mass by mechanical strain magnitude. Calc. Tiss. Int. 37, 41–417.