Mutations in valosin-containing protein (VCP) decrease ADP/
ATP translocation across the mitochondrial membrane
and impair energy metabolism in human neurons
Received for publication, October 11, 2016, and in revised form, March 29, 2017 Published, Papers in Press, March 30, 2017, DOI 10.1074/jbc.M116.762898
X Marthe H. R. Ludtmann‡1, Charles Arber‡1, Fernando Bartolome§¶1, Macarena de Vicente§¶,XElisavet Preza‡, X Eva Carro§¶, Henry Houlden储,X Sonia Gandhi**,X Selina Wray‡, andX Andrey Y. Abramov‡2
From the‡Department of Molecular Neuroscience, UCL Institute of Neurology, London WC1N 3BG, United Kingdom, the §Neurodegenerative Disorders Group, Research Institute Hospital 12 de Octubre (i⫹12), Madrid 28041, Spain, the储Institute of
Neurology, MRC Centre for Neuromuscular Diseases, London WC1N 3BG, United Kingdom, the**Sobell Department of Motor Neuroscience and Movement Disorders, UCL Institute of Neurology, London WC1N 3BG, United Kingdom, and the¶Biomedical Research Networking Center on Neurodegenerative Diseases (CIBERNED), Madrid 28041, Spain
Edited by Roger J. Colbran
Mutations in the gene encoding valosin-containing protein (VCP) lead to multisystem proteinopathies including fronto-temporal dementia. We have previously shown that patient-derived VCP mutant fibroblasts exhibit lower mitochondrial membrane potential, uncoupled respiration, and reduced ATP levels. This study addresses the underlying basis for mitochon-drial uncoupling using VCP knockdown neuroblastoma cell lines, induced pluripotent stem cells (iPSCs), and iPSC-derived cortical neurons from patients with pathogenic mutations in VCP. Using fluorescent live cell imaging and respiration analysis we demonstrate aVCPmutation/knockdown-induced dysregu-lation in the adenine nucleotide translocase, which results in a slower rate of ADP or ATP translocation across the drial membranes. This deregulation can explain the mitochon-drial uncoupling and lower ATP levels in VCP mutation-bearing neurons via reduced ADP availability for ATP synthesis. This study provides evidence for a role of adenine nucleotide trans-locase in the mechanism underlying altered mitochondrial function in VCP-related degeneration, and this new insight may inform efforts to better understand and manage neurodegen-erative disease and other proteinopathies.
Valosin-containing protein (VCP)3 (alias p97/TERA) is a ubiquitously expressed member of the type II AAA⫹ATPase family. Mutations in VCPhave been found in patients with inclusion body myopathy with early-onset Paget disease and frontotemporal dementia as well as familial amyotrophic lateral
sclerosis (ALS). VCP has been linked to a variety of cellular functions including ubiquitin-proteasome system, protein deg-radation at the outer mitochondrial membrane, and autophagy but the underlying molecular mechanism in disease is yet to be fully understood.
Clues to the molecular mechanism associated with VCP mutations have come from model organisms that possess high inter-species homologies in the VCP protein. In particular, mutant mouse knock-in models mirror age-depandant motor-neuron dysfunction (1, 2). Abnormalities have been found in mitochondria cristae structures suggesting mitochondrial pathology inVCPmutation bearing cells (3). Our recent work has provided an insight into the observed mitochondrial pathologies through the utilization of a human dopaminergic neuroblastoma VCP knockdown cell line (SH-SY5Y), fibro-blasts from patients carrying pathogenic mutations in theVCP gene andVCPknockdown mouse neurons, where we revealed an uncoupling of respiration from oxidative phosphorylation. Furthermore, it was shown that uncoupling leads to a lower mitochondrial membrane potential (indicator of cell health), higher respiration, and lower ATP levels in these cells (4). We hypothesized that the VCP mutation-induced uncoupling event may occur due to one of two reasons: 1) uncoupling pro-teins (UCPs) may alter the mitochondrial proton gradient, thus uncoupling respiration from oxidative phosphorylation. 2) Dysfunctional adenine nucleotide translocase (ANT1, SLC25A4), which is responsible for the simultaneous ADP import and ATP export across the mitochondrial inner membrane, leading to reduced ADP substrate for oxidative phosphorylation.
Other studies have investigated the effects of VCP on cellular health in patient fibroblasts, SH-SY5Y, as well as myoblasts (5) and this present study is the first to employ a disease-relevant neuronal model. In this current study, we set out to generate a human neuronal model ofVCPmutations to explain the under-lying mechanism of mitochondrial uncoupling inVCPmutant cells. To produce a human neuronal cell model, expressing mutant VCP at physiological levels, we generated iPSCs from patient fibroblasts and subsequently differentiated the cells to cortical neurons to study mitochondrial function. Live cell imaging techniques and a combination of VCP knockdown This work was supported by the National Institute for Health Research (NIHR)
University College London Hospitals (UCLH) Biomedical Research Centre (BRC), a Wellcome Trust grant, and the NIHR Queen Square Dementia Bio-medical Research Unit UCL/UCLH. The authors declare that they have no conflicts of interest with the contents of this article.
Author’s Choice—Final version free via Creative Commons CC-BY license.
1These authors should be considered first co-authors.
2To whom correspondence should be addressed. Tel.: 44-2034484062;
E-mail: [email protected].
3The abbreviations used are: VCP, valosin-containing protein; ANT, adenine
nucleotide translocase; UCPs, uncoupling proteins; KD, knockdown; ETC, electron transport chain; CATR, carboxyatractyloside; FTD, frontotemporal dementia; iPSC, induced pluripotent stem cell; SCR, scrambled; RCR, respi-ratory control rate; qPCR, quantitative PCR; BKA, bongkrekic acid.
ARTICLE
Author’s Choice
J. Biol. Chem.(2017) 292(21) 8907–8917
8907
at ROYAL VET COLL./LIBRARIAN on July 24, 2018
http://www.jbc.org/
SH-SY5Y cell lines as well as iPSCs and iPSC-derived neurons enabled us to establish that a reduced activity of ANT1 leads to the uncoupling of mitochondria inVCPmutation bearing cells.
Results
Uncoupling of VCP mutation bearing cells is not caused by UCPs
Our previous study provided evidence of lower ATP levels as well as uncoupled mitochondria (4). To investigate this further, we assessed mitochondrial coupling in stableVCPknockdown (KD) and scrambled (SCR) SH-SY5Y neuroblastoma cell lines. TheVCPKD mimics the dominant-negative effect displayed by pathogenic loss-of-functionVCPmutations. As previously shown by Bartolomeet al.(4), VCPKD cells (clones KD#1, KD#2 and KD#3) displayed a significantly lower respiratory control rate (RCR; an indicator of coupling of oxidative phos-phorylation and respiration) when compared with SCR cells (Fig. 1 A;nⱖ3 experiments;p⬍0.01). To assess whether the mitochondrial uncoupling in SH-SY5YVCPKD cells is caused by UCPs, permeabilized cells were pre-exposed for 1 min to the UCP inhibitors GDP (100M) or Genipin (50M) and the cou-pling of oxidative phosphorylation and respiration was assessed (Fig. 1A). Application of GDP and Genipin did not alter the RCR of SCR orVCPKD cells indicating that UCPs are not the underlying reason for the mitochondrial uncoupling in VCP mutants (nⱖ3 experiments). In support of this finding, West-ern blotting for UCP2 showed equal levels in control andVCP KD cells (Fig. 1B). Protein levels of mitochondrial electron
transport chain (ETC) complexes were also found to be unchanged inVCPKD cells when compared with control sug-gesting that the mitochondrial phenotype is not caused by an alteration in ETC complex expression (Fig. 1D).
Lack of ATP synthase substrate in VCP KD cells
Due to the difference in charge between ATP and ADP, changes in ANT activity can affect mitochondrial membrane potential and respiration. To test whether this is the underlying cause of the uncoupling event observed in VCPknockdown cells, we exposed SCR control andVCPKD cells to ANT inhib-itors carboxyatractyloside (CATR; 5 M) or bongkrekic acid
(BKA) (5 M) and RCR was assessed. Inhibition of ANT resulted in a significantly reduced RCR in SCR cells, which mir-rors the RCR observed in SH-SY5YVCPKD cells (Fig. 1A;p⬍ 0.01;nⱖ3 experiments). Furthermore, the lower RCR in the ANT inhibitor-treated SCR cell suggests that inhibition of ANT alone is able to uncouple mitochondria. There was no addi-tional reduction in RCR inVCPKD cells exposed to ANT inhib-itors suggesting that ANT is already inhibited and substrate (ADP) supply is reduced (Fig. 1A). To exclude a reduced ANT1 expression being the underlying reason, a Western blot analysis probing for ANT1 was undertaken. No changes in total ANT1 protein expression were found inVCPKD cells (Fig. 1B).
A proton leak through the ATP synthase could be the under-lying reason for uncoupled mitochondria inVCPKD cells (6). Therefore, the irreversible ATP synthase inhibitor oligomycin (2 g/ml) was applied (Fig. 1C). Oligomycin induced much
A
C
D
B
0.1 0.2 0.3 0.4 0.5 0.6 0.7 0.8 0.9 1.0
* *
KD#3 KD#2 KD#1 VB
a
sal
/V
Oligom
ycin
SCR
* 0.0
0.5 1.0 1.5 2.0 2.5
**
RCR (V3/V4)
SCR KD#1 KD#2 KD#3
**
* **
Vehicle +G
DP
+Genipin +C
ATR +BKA
Figure 1.A,RCR is the ratio of state 3 respiration (ADP stimulated) to state 4 respiration (no ADP present), and is considered an indication of the degree of coupling of mitochondrial respiratory chain activity to oxidative phosphorylation. RCR of mitochondria in permeabilized SHY5Y control andVCPknockdown cells (KD#1, KD#2, KD#3) in the presence of vehicle control, uncoupling protein inhibitors (GDP;Genipin), or ANT inhibitors (CATR;BKA).B,SHY5Y SCR and KD cells were probed for VCP, ANT1, UCP2, and-actin by Western blotting (cropped).C,quantification ofVBasal/Voligomycinin permeabilized SHY5Y control and VCP
knockdown cells as an alternative indicator of coupling of mitochondrial respiratory chain activity to oxidative phosphorylation.D,full-length representative Western blot analysis of whole cell lysates from stable SCR SH-SY5Y cells and stableVCPknockdown SH-SY5Y cells probing for mitochondrial complexes and -actin. *, p⬍0.05; **,p⬍0.01; ***,p⬍0.001.
at ROYAL VET COLL./LIBRARIAN on July 24, 2018
http://www.jbc.org/
smaller inhibition of respiration inVCPKD cells providing evi-dence that the uncoupling of these mitochondria is not caused by a proton leak in these cells. These results suggest that sub-strate supply in the form of ADP is limited in VCP KD cells leading to mitochondrial uncoupling.
Generation of VCP mutant iPSCs and cortical neurons
To further our understanding of mutant-VCP disease mech-anism, we employed a more physiologically relevant model in the form of patient-derived iPSCs and iPSC-derived neurons.
Fibroblasts from two patients carrying the R155C (most prevalentVCPmutation) and R191Q mutations were repro-grammed to iPSCs using episomal reprogramming factors (7). These patients both exhibited autosomal dominant inclusion body myopathy and frontotemporal dementia (FTD). The resultant iPSC clones exhibited similar expression of pluripo-tency markers to control hESCs at the protein (Fig. 2A) and RNA level (Fig. 2B). Reprogrammed cells exhibited a stable karyotype with unaltered g-banding (data not shown) and DNA sequencing confirmed that these stem cell lines retained the VCPmutation (Fig. 2D). The potential of the cells to differen-tiate into three germ layers was confirmed (Fig. 2E). Two clones from each patient were taken forward for further experiments and compared with two control iPSC lines and one control hESC line.
iPSCs were differentiated into cortical neurons, which are affected in VCP-linked FTD, as described previously (8). After 80 days of neurogenesis, control andVCPmutant lines gener-ated similar proportions of deep (TBR1), middle (CTIP2), and upper (SATB2) cortical neurons (Fig. 2,A,ii, andF). The faith-ful silencing of pluripotency marker gene expression reinforces the footprint free reprogramming method (Fig. 2C). SOX2 expression remains high; representing the neural stem cells that persist within the culture (9). The fact thatVCPmutant iPSCs can generate cortical neurons at a similar efficiency to control lines hints at a normal capacity ofVCPmutant cells for neuro-genic differentiation and supports disease associated to aging rather than developmental abnormalities. The glutamatergic identity was confirmed by loading neurons with the cytosolic calcium indicator Fura-2 and application of a physiological concentration of glutamate (1M), which resulted in a transient
intracellular calcium signal (Fig. 2G). Furthermore, a mature glutamatergic neuronal phenotype was validated via punctate vGLUT1 staining and expression of the post-synaptic marker PSD95 (Fig. 2H). Together, these results demonstrate an effec-tive ability of the cells to differentiate into cortical neurons and represent an appropriate model to studyVCPmutations in the cell type affected in FTD.
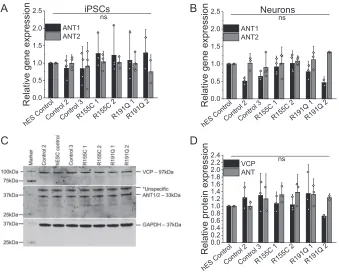
VCP and ANT expression is not affected in patient-derived neurons
To further investigate the restricted ATP synthase substrate supply through ANT, we employed patient-derived iPSCs and cortical neurons bearing VCP mutations. First, we assessed
ANT1/2gene expression on the mRNA level in iPSCs and
iPSC-derived neurons where no significant changes could be found in R155C or R191Q mutation-bearing cells (Fig. 3,AandB). Pro-tein expression levels of VCP andANT in the iPSC-derived
mutant neurons were also not changed when compared with control neurons (Fig. 3,CandD); reinforcing the dominant-negative theory thatVCPmutations interfere with VCP hex-amer function without affecting expression levels (10).
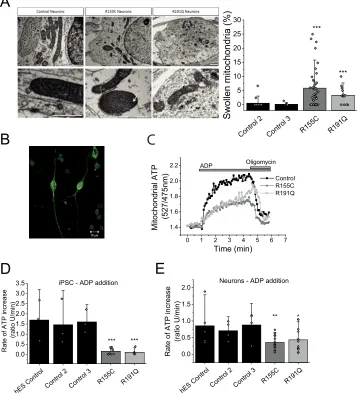
Mitochondrial swelling in iPSC-derived neurons bearing a VCP mutation
Mitochondrial morphology was assessed in iPSC-derived neurons by EM (representative images in Fig. 4A, i). We observed that the number of swollen mitochondria in VCP mutant R115C neurons and R191Q was significantly higher (p⬍0.001) when compared with control neurons (Fig. 4A,ii). This was confirmed in three independent neuronal inductions and not seen in three parallel control cultures. Additionally, swollen mitochondria displayed cristae that were disordered and interdigitation was disrupted (Fig 4A, i, white arrows). These observations in mitochondrial morphology further sup-port that mitochondrial health is likely to be affected inVCP mutant iPSC-derived neurons. It should be noted that abnor-mal mitochondrial morphology has been witnessed in other
VCP mutant models, such as R155H homozygous and
heterozygous mice (3, 11).
Lower ATP synthase activity in VCP mutation bearing cells Oxygen consumption measurements are an indirect assess-ment of ATP synthase efficiency rather than ATP levels. To measure mitochondrial ATP levels in real-time, a FRET-based ATP probe was employed to allow assessment of ATP level changes (12). Patient-derived iPSCs and neurons were trans-fected with this probe where the fluorescent signal reflects mitochondrial ATP levels (Fig. 4B). The transfected cells were then permeabilized (40 M digitonin to permeabilize the
plasma membrane) in pseudo-intracellular solution and ATP synthase activity was assessed in the presence of mitochondrial substrates (5 mMmalate and 5 mMglutamate) and ADP (500 nmol) (13). The applied ADP is translocated across the mito-chondrial membranes via ANT where it is converted to ATP by the ATP synthase. ATP binds to the probe, which allows fluo-rescent live time recordings (Fig. 4C). All measurements were carried out in 3 control cell lines and two patient cell lines bearing either R155H or R191Q mutations (pool of two clones each) and statistical analysis was carried out on the pool of all control cell linesversusmutant cell lines.
iPSCs of affected patients had a significantly lower rate of ATP synthesis when compared with control (Fig. 4D). The rate of ATP increase in R155C (n⫽10) and R191Q (n⫽6) was significantly slower (p ⬍ 0.001) when compared with the pooled unaffected iPSC controls (n⫽9; Fig. 4D). The rate was also significantly lower in neurons bearing the R155C mutation (n⫽17;p⬍0.001; Fig. 4D) and R191Q mutation (n⫽12;p⬍ 0.05) when compared with control iPSC-derived neurons (n⫽ 13).
These results provide evidence that the ATP synthase inVCP mutant cells does not convert ADP to ATP as efficiently when compared with control cells. However, this approach does not distinguish between ATP synthase malfunction or a lower sub-strate (ADP) supply as the underlying basis for the difference in rate in ATP synthesis between control andVCPmutant cells.
at ROYAL VET COLL./LIBRARIAN on July 24, 2018
http://www.jbc.org/
A ii ii
hESC con
trol
Fibrobl ast
R155C 1
R155C 2
R191Q 1
R191 Q 2
0.0 0.2 0.4 0.6 0.8 1.0 1.2 1.4 1.6 1.8
Relative gene expression
OCT4 NANOG SOX2 DNMT3b
hESC cont
rol
hESC cont
rol Cont
rol 2 Cont
rol 3 R155CR191
Q 0.0
0.4 0.8 1.2 1.6iPSC
R
e
la
ti
v
e
gene ex
pr
es
s
ion
OCT4 SOX2 NANOG DNMT3b
Neurons
B C
D EE
0 1 2 3 4 5 6 7 0.8
1.0 1.2 1.4 1.6 1.8
Fu
ra
-2
ra
tio
Time (min)
1µM Glutamate
0 20 40 60 80 100
R191Q 2 R191Q
1 R155C
2 R155C 1 Cont
rol 3 Cont
rol 2
P
e
rc
ent
age of
tot
al
cel
ls TBR1/DAPI
CTIP2/DAPI SATB2/DAPI
hES Cont rol
F G
H
at ROYAL VET COLL./LIBRARIAN on July 24, 2018
http://www.jbc.org/
ADP/ATP translocation is inhibited in VCP mutation bearing cells
The translocase activity of ANT can be reversed through the application of high concentrations of ATP to permeabilized cells, which enables the import of ATP across the mitochon-drial membranes. The translocated ATP is then bound by the mitochondrial ATP probe and the kinetic fluorescent measure-ments are a direct assessment of ATP translocation by ANT. It should be noted that this process is independent of the ATP synthase or ETC substrates and merely a measurement of ATP uptake, hence ANT activity (Fig. 5A). This method allowed investigation whether a lower substrate supply in the form of ADP is the reason for uncoupling.
Application of ATP (500 nmol) to permeabilized cells re-vealed a significantly slower ATP uptake into mitochondria of R155C iPSCs (n⫽7;p⬍0.001) as well as R191Q iPSCs (n⫽14; p⬍0.01) when compared with control cell lines (n⫽8; Fig. 5,
AandB). This observation was reproduced in iPSC neurons
where ATP uptake into the mitochondria was significantly reduced in R155C neurons (p⬍0.001) and R191Q when com-pared with control neurons (n⫽8; Fig. 5C).
As a proof of principle, permeabilized control iPSCs were exposed to ANT inhibitors BKA or CATR for 30 min and the rate of ATP uptake was recorded (Fig. 5,DandE). Mitochon-dria pre-exposed to either ANT inhibitor displayed a signifi-cantly reduced ATP uptake (CATRn ⫽4; BKAn⫽ 5;p⬍ 0.001), mirroring the uptake observed in untreated patient iPSC (n⫽8), when compared with control (n⫽8). The rate of ATP uptake in iPSC-derived control neurons exposed to CATR (p⬍0.01) and BKA (p⬍0.05) was also significantly lower when compared with control neurons (n⫽3; Fig. 5E). These results provide evidence that the mitochondrial uncoupling and lower ATP levels inVCPmutation bearing cells are caused by a lack of substrate (ADP) being translocated from the cytoplasm across the mitochondrial membranes.
Discussion
We have previously shown that fibroblasts from patients harboring mutations in theVCP gene display mitochondrial uncoupling, seen by increased respiration, reduced oxidative phosphorylation, and lower ATP levels (4). In the present study we focused on the mechanism of this mitochondrial
uncou-Figure 2.A,immunocytochemistry showing three control iPSCs and two patient-derivedVCPmutant iPSCs lines (2 clones each). All stem cells show expression
of pluripotency markers OCT4 and SSEA4 in stem cell conditions and have characteristic colony morphology. By day 80 of differentiation,-III-tubulin positive neurons are formed expressing markers of deep (TBR1), middle (CTIP2), and upper (SATB2) cortical neurons. Size bar⫽100m.B,qPCR analysis of four pluripotent stem cell markers in VCP patient-derived iPSCs and control fibroblasts, normalized to three housekeeping genes.C,qPCR analysis to represent faithful silencing of pluripotency genes in neuronal cultures, relative to hESC controls.D,DNA sequencing of iPSC-derived neuron genomic DNA showing mutated nucleotides corresponding to amino acid residues 155 and 191 (arrows).E,monolayer directed differentiation, showing the ability of newly derived iPSC lines to enter the endoderm and mesoderm differentiation pathways.Size bar⫽50m.F,automated counting of mature neuronal markers displays no difference in the ability of control and VCP iPSCs to form deep (TBR1), middle (CTIP2), and upper (SATB2) layer cortical neurons.G,representative traces of Fura-2-loaded iPSC-derived neurons showing a transient cytosolic calcium response to physiological concentrations of glutamate. Representative image depicts iPSC-derived neurons loaded with Fura-2.H,iPSC-derived neurons express markers of glutamatergic neurons (vGLUT1) and functional synapses (PSD95).Size bar⫽20m.
A
B
C
D
0.0 0.5 1.0 1.5 2.0 2.5
R
e
la
ti
ve gene expr
essi
on
R191Q 2
R191Q 1
R155C 2 R155C 1 Cont
rol 3 Cont
rol 2
hES Co ntro
l
ANT1 ANT2
ns Neurons
0.0 0.5 1.0 1.5 2.0 2.5
R
el
at
ive gene expr
essi
o
n
R191Q 2
R191Q 1 R155C 2 R155C 1 Cont
rol 3 Cont
rol 2
hES C ontro
l
ANT1 ANT2
ns iPSCs
0.0 0.2 0.4 0.6 0.8 1.0 1.2 1.4 1.6 1.8 2.0 2.2 2.4
R
el
a
ti
ve pr
ot
ei
n expr
essi
on
R19 1Q 2 R191Q
1
R155 C 2
R155C 1 Cont
rol 3 Cont
rol 2
hES Cont rol
VCP ANT
ns
Figure 3.Quantification ofANT1/2gene expression in iPSC (A) and iPSC-derived neurons (B) by qPCR. Number of repeats are indicated in the bar chart.C,
cropped Western blotting andD,densitometry analysis of VCP and ANT1/2 in iPSC-derived neurons. GAPDH was used as a loading control.
at ROYAL VET COLL./LIBRARIAN on July 24, 2018
http://www.jbc.org/
pling. The UCP family and ANT both play an important role in regulating the coupling of the mitochondrial respiratory chain to oxidative phosphorylation. UCPs are mitochondrial carriers functioning via dissipation of the proton gradient across the inner membrane (14). To date, five human UCP homologues have been identified in different tissues and all are located at the mitochondria. UCP1 has a well characterized thermogenic role in brown adipose tissue. UCP3 is mainly expressed in both brown adipose tissue and muscle, whereas UCP2, UCP4, and UCP5 are mainly expressed in the central nervous system and therefore play key roles in neurons. Although only the role of UCP1 has been well characterized, it is generally accepted that all five UCPs uncouple oxidative phosphorylation from ATP synthesis by dissipating the proton gradient across the inner membrane (14).
Mitochondrial respiration analysis and Western blotting of VCPKD cells established that uncoupling of respiration from oxidative phosphorylation is not caused by UCPs or a proton
leak through the ATP synthase. However, ANT inhibition in VCPKD did not lead to further uncoupling indicating that ANT could already be inhibited. Classical uncoupling is understood to be a reduction in coupling between the rate of electron trans-port in the respiratory chain (ADP independent; State 4) and ATP production (ADP-dependent; State 3) leading to a lower RCR. However, a reduced substrate supply (ADP) through lim-ited uptake via ANT will lead to state 3 being inhiblim-ited resulting also in a lower RCR. Both scenarios decrease ATP levels and will ultimately lead to a lower bioenergetic status of the cell.
ANT1 is a mitochondrial protein that is thought to have an intrinsic uncoupling property (15–17). Additionally, ANT1 has been associated with the mitochondrial permeability transition pore and believed to be involved in mitochondria-mediated apo-ptosis (18). This study provided evidence that reduced ANT activity can lead to mitochondrial uncoupling as shown by ANT inhibition in SCR cells. The observed mitochondrial deficiency in VCPKD cells is in agreement with our previous work (4)
A
Control 2 Con trol 3
R155C R191Q 0
5 10 15 20 25 30
***
Swollen mitochondria (%)
***
B
D
hES Con trol
Control 2 Control 3
R155C R191Q
0.0 0.5 1.0 1.5 2.0 2.5 3.0 3.5
*** iPSC - ADP addition
Rate of ATP increase
(ratio U/min)
***
0 1 2 3 4 5 6 7
1.4 1.6 1.8 2.0 2.2
Mitochondrial ATP
(527/475nm)
Time (min)
Control R155C R191Q
ADP Oligomycin
C
E
hES C
ontrol
Cont
rol 2
Cont
rol 3
R155C R191
Q 0.0
0.5 1.0 1.5 2.0
** Neurons - ADP addition
R
a
te of A
T
P
i
n
c
reas
e
(r
a
ti
o U
/m
in)
*
Figure 4.A,i, transmission electron microscopy images of mitochondria within control and mutant iPSC-derived neurons at day 80 of differentiation. Mutant
neurons consistently display a small proportion of swollen, electron dense mitochondria.Arrowsshow damaged cristae.Scale barrepresent 1m.ii, quanti-fication of swollen mitochondria.n⫽3 control,n⫽3 R155C andn⫽1 R191Q.B,representative image of a neuron transfected with mitAT1.03 probe.C, representative traces of mitochondrial ATP after ADP application. Oligomycin was applied as a negative control.Scale barrepresent 10m.D,quantification of the rate of ATP increase in control and VCP mutation bearing iPS cells.E,quantification of the rate of ATP increase in control and VCP iPS-derived neurons. *,p⬍0.05; *,p⬍0.05; ***,p⬍0.001.
at ROYAL VET COLL./LIBRARIAN on July 24, 2018
http://www.jbc.org/
where a lower NADH redox index and faster basal V2 respira-tion inVCPKD cells was reported. These features, together with a lower RCR, have been implicated in a number of neuro-degenerative models. For example, Plun-Favreau et al. (6) showed similar phenotypes in an HTRA2 knock-out mouse model where mutations in theHTRA2gene have been linked to Parkinson’s disease. Additionally, studies have shown how the most common genetic cause of Parkinson’s disease, G2019S mutations in the leucine-rich repeat kinase 2 gene (LRRK2), displayed uncoupling and low RCR implicating UCP2 and -4 proteins (19). Other transgenic mice for Huntington’s disease reflected metabolic and thermoregulatory defects associated to uncoupling and reduced RCRs (20).
We extended our study by employing VCP patient-derived iPSCs and iPSC-derived neurons, which allowed us to
investi-gate the dynamics of ADP and ATP translocation at the mito-chondria in a human neuronal model. Investigations utilizing this highly energy-demanding cell type enabled us to validate previously observed mitochondrial abnormalities in the cell type affected by FTD.
Electron microscopy of mitochondria in iPSC-derived neu-rons displayed a significantly higher subset of swollen mito-chondria that exhibit disordered inner mitomito-chondrial mem-branes in VCP-deficient cells compared with those from control derived neurons. Damaged mitochondria were de-scribed as a common feature in VCP-associated disease models and in patient cell samples but there have been no reports showing this feature in neurons (5, 11, 21). For example,
John-sonet al.(21) in 2015 showed damaged mitochondria in muscle
cells fromDrosophilaand Nalbandianet al.(11) in 2012 showed
0.0 0.5 1.0
0.8 1.0 1.2 1.4 1.6 1.8 2.0 M itochondr ial A T P (527/475nm ) Time (min) Control R155C R191Q ATP
B
hES C ontrol Cont rol 2 Cont rol 3 R155 C R191 Q 0.0 0.5 1.0 1.5 2.0iPS - ATP addition
** R a te of A T P upt ak e (ra tio U/min ) *
A
D
Cont rol Cont rol +CATR
Cont rol +
BKA R191 Q 0 1 2 3 4 R a te o f AT P i n c re a s e (r a tio U/m in ) *** iPSCs - ATP addition
ns
hES C ontrol
Cont rol 2
Cont rol 3
R155C R191Q
0 1 2 3 4 ***
Rate of ATP uptake
(ratio U/min)
Neurons - ATP addition
***
E
C
Cont rol Cont rol + CA TR Cont ro + BKA R155C R191Q 0.0 0.2 0.4 0.6 0.8 1.0 1.2 1.4 1.6 1.8 2.0 Ra te o f AT P u p ta k e (ra ti o U /mi n ) ** *Neurons - ATP addition
ns
Figure 5.A,representative traces of mitochondrial ATP uptake.B,quantification of the rate of ATP uptake in control and VCP mutation bearing iPS cells.C,
quantification of the rate of ATP uptake in control and VCP iPS-derived neurons.D,quantification of the rate of ATP uptake in control iPSCs or pre-exposed to ANT inhibitors (BKA/ATRAC) as well as mutation bearing iPSCs.E,quantification of the rate of ATP uptake in control neurons, neurons pre-exposed to ANT inhibitors (BKA/ATRAC), and mutations bearing neurons. *,p⬍0.05; **,p⬍0.01; ***,p⬍0.001.
at ROYAL VET COLL./LIBRARIAN on July 24, 2018
http://www.jbc.org/
mitochondrial megaconia and disrupted cristae in myoblasts from a VCP mouse model. The same group more recently showed damaged mitochondrial dynamics and function in fibroblasts and myoblasts fromVCPpatients (5). Here for the first time, we are showing EM images of damaged mitochondria in human neurons withVCPmutations. Additionally, the swol-len mitochondria observed in theVCPmutant neurons could be an indicator of uncoupling, a feature originally described by Weinbachet al.(22) in 1963 and subsequently confirmed in a number of reports (23, 24).
The results suggest a decreased transport of ADP/ATP by ANT, which explains the mechanism behind the reduced ATP levels in the cells as previously shown in multiple other disease models (5, 25, 26). Inhibition of ADP/ATP transport could be observed in all cell types employed by this study. However, neu-rons require more energy than any other cell type (such as iPSC) and rely predominantly on oxidative phosphorylation for ATP production (with a very small impact of glycolysis). This makes neurons more vulnerable to mitochondrial uncoupling and inhibition of ATP transport (27).
Our data provide evidence that mutations in VCP lead to mitochondrial uncoupling due to a reduced ADP/ATP translo-cation by ANT. Here we provide unique analysis of the bioen-ergetics ofVCPmutant human neurons with patient genotypes, allowing rational and relevant bioenergetic analysis in the cell type affected by neurodegeneration. The underlying deficiency in mitochondrial bioenergetics may help to explain disease in the context ofVCPmutation and provide a focus for further studies into the brain metabolism in healthy aging and disease.
Experimental procedures
Cell culture
Human neuroblastoma (SH-SY5Y) cells were purchased from the European Collection of Cell Cultures (Health Protec-tion Agency, Salisbury, UK) and maintained as previously described (28). Unless otherwise stated, SH-SY5Y cells were seeded at a density of 4⫻ 104cells/cm2and maintained in Dulbecco’s modified Eagle’s medium (DMEM) supplemented with 10% (v/v) fetal bovine serum (FBS), 2 mM L-glutamine, and
1% (v/v) penicillin/streptomycin at 37 °C and 5% CO2. The scramble and VCP KD SH-SY5Y cells were generated by lenti-viral infection with either a pool of 5 shRNA vectors or individ-uals shRNA (Thermo Fisher) as described previously by Bar-tolomeet al.(4). Subsequent clonal selection was carried out using puromycin (1g/ml). Two different independent clones and a pool with 90% approximate reduction in gene expression were used in all experiments. 1 clone expressing nontargeting RNA vector was used as control (stable SCR).
Cell differentiation
Fibroblast and iPSC cultures were performed as described previously (29, 30) and all reagents were obtained from Life Technologies, unless otherwise stated. Clinical data about the patients was described previously (4). Fibroblasts were cultured in DMEM with 10% fetal bovine serum and passaged with 0.05% trypsin. Reprogramming was performed as per the method detailed by Okitaet al.(7) using episomal reprogramming plas-mids (given as a gift from Shinya Yamanaka; Addgene plasplas-mids
#27077, #27078, and #27080) and the P2 nucleofection kit (Amaxa). One week after nucleofection, cells were passaged onto mouse embryonic feeders (AMS Bio) and media was changed to KO-DMEM, 20% knock-out serum replacement, 2 mM L-glutamine, non-essential amino acids, 50M
-mercap-toethanol, and 10 ng of FGF2 (Peprotech).
Newly formed iPSC colonies were manually picked and, together with control iPSCs and hESCs, were maintained in Essential 8 medium on Geltrex-coated plates. Karyotype anal-ysis was performed by Cell Guidance Systems, Cambridge, UK. Differentiation to cortical neurons was performed as detailed (8, 34). 10 days of dual SMAD inhibition using 1M dorsomor-phin and 10MSB431542 was followed by 70 –90 additional
days of differentiation in N2B27 media containing retinoids on laminin-coated dishes and coverslips. The hESC line SHEF6 was obtained from the United Kingdom Stem Cell Bank, con-trol iPS line 1 was obtained from the lab of Dr. Tilo Kunath, and control iPS line 2 from Coriell repository.
For mesoderm and endoderm fate specification a defined monolayer protocol was followed, which allowed up-regulation of mesoderm- and endoderm-specific fate determinant gene networks, as published by others (31, 32). In brief, endoderm differentiation was achieved via a 3-day treatment consisting of 0.1 g/ml of activin (days 1–3), 80 ng/ml of FGF2 (days 1–3), 10 ng/ml of BMP4 (days 1–2), and 3MChir (day 1). Mesoderm
differentiation was directed via a 2-day treatment with 0.1 g/ml of Activin, 20 ng/ml of FGF2, 10 ng/ml of BMP4, and 5 MChir.
Immunocytochemistry
Cells were fixed in 4% paraformaldehyde for 15 min and washed three times in PBS containing 0.3% Triton X-100 (PBST), before blocking in 3% BSA in PBST for 20 min. Primary antibodies (Table 1) were incubated overnight at 4 °C in block-ing solution, followed by three washes in PBST. Secondary anti-bodies were then added for 1 h at room temperature in the dark (Alexa Fluor, Invitrogen), DAPI was added for one wash at 1M
and a final two washes was performed in PBST before mounting the coverslips for imaging on a Zeiss LSM microscope or a Zen confocal microscope. Cell counts were performed on 5 images per condition from three independent experiments, using the Table 1
Antibodies used for immunocytochemistry and Western blotting
Antibody name Source Dilution Species OCT4 Santa Cruz 1:500 gt
SSEA4 BioLegend 1:200 m
MAP2 Abcam 1:10,000 ch
PSD95 Abcam 1:1000 m
vGLUT1 Synaptic systems 1:1000 rb
TUJ1 Covance 1:1000 m/rb
TBR1 Abcam 1:500 rb
SATB2 Abcam 1:100 m
CTIP Abcam 1:500 rat
VCP Cell Signalling 1:1000 m
ANT1/2/3 Santa Cruz 1:200 gt
UCP2 (C-20) Santa Cruz 1:200 gt
GAPDH Ambion 1:5000 m
-actin Sigma 1:5000 rb
hGATA-4 Santa Cruz 1:100 m
hSox17 R&D 1:200 gt
hCDX1 Abcam 1:200 r
OXPHOS complexes Abcam 1:250 m
at ROYAL VET COLL./LIBRARIAN on July 24, 2018
http://www.jbc.org/
ITCN nuclear counting software plug-in for ImageJ with con-sistent settings between all conditions.
Western blotting analysis
Cells were lysed for 1 h at 4 °C in lysis buffer containing 10 mMTris, pH 8, 140 mMNaCl, 1 mMEDTA, 0.5 mMEGTA, 1%
Triton X-100, 0.1% sodium deoxycholate, 0.1% SDS plus prote-ase and phosphatprote-ase inhibitors (Roche Applied Science), before centrifugation at 10,000 rpm for 15 min at 4 °C. Proteins were separated on NuPage 10% SDS-polyacrylamide gels (Novex) and transferred onto nitrocellulose membranes. Membranes were blocked in PBS containing 0.1% Tween and 3% milk solu-tion. Primary antibodies were incubated in blocking solution overnight at 4 °C (Table 1). Subsequent washes were performed in PBS-Tween and secondary antibodies (800 nm Rockland and 680 nm Molecular Probes) were incubated for 1 h at room tem-perature. Final washes were in PBS and membranes were scanned on an Odyssey imaging system (Li-Cor). All gels have been run under the same experimental conditions.
qPCR analysis
RNA was isolated from cells using TRIzol reagent and the manufacturer’s instructions (Thermo Fisher). Reverse tran-scription was performed using SuperScript III Reverse Tran-scriptase and random hexamer primers (Thermo Fisher). qPCR was performed using Power SYBR Green master mix (Thermo Fisher) and the Agilent Mx300P qPCR system and software with annealing of primers at 60 °C (Table 2). All results are relative to the expression of three housekeeping genes,GAPDH,
CYCLOPHILIN, and-ACTIN.
Sanger sequencing
Genomic DNA was extracted from VCP cells using the QuickExtract DNA extraction solution (Epicenter) as per the manufacturer’s instructions. The regions of the VCP mutations were PCR amplified with VCP exon 5 (forward, TTAAGACA-GGTGGGGTGGAG; reverse, CCCATCTCAGTCTCCCA-AAG) primers using POLYMERASE HERE (it may be: FastStart PCR Master (Roche Applied Science)) in PCR containing 10l of FastStart of PCR Master, 1l of each primer, 1l of DMSO, 10l of H2O, and 1l of extracted gDNA. PCR was performed using a 65td55 standard cycling program. PCR products were subsequently purified by incubation with Exonuclease I/ FastAPTM Thermosensitive Alkaline Phosphatase (Thermo Scientific) mixture as per the manufacturer’s instructions. 5l of purified products were used in sequencing reactions with 0.5 l of Big Dye (BigDye Terminator V.3.1 Cycle Sequencing Kit,
Applied Biosystems), 2l of sequencing buffer (BigDye Termi-nator V.3.1 Cycle Sequencing Kit, Applied Biosystems), 1l of primer, and 2.5l of H2O. For the removal of BigDye Termi-nator, sequencing reactions were filtered through Sephadex (Sigma) containing filter plates (Corning). Sequencing was per-formed using an Applied Biosystems 3730XI Sequencer and analysis of results was carried out using the Sequencher soft-ware (Gene Codes Corporation).
Transmission electron microscopy
Neurons were fixed in 3% gluteraldehyde (EM grade, Agar Scientific), 0.1Msodium cacodylate, and 5 mMcalcium chlo-ride. Neurons were then post-fixed in osmium tetroxide, stained with uranyl acetate and Reynold’s lead citrate, and dehydrated with stepwise washes with increasing concentra-tions of ethanol. Cells were then mounted in epoxy resin using propylene oxide to aid infiltration, which was hardened at 60 °C for 2 days. Ultrathin sections were taken using a diamond knife (Leica Reichert Ultracut), sections were mounted on copper mesh grids, stained with uranyl acetate, and images were taken using the Philips CM12 transmission electron microscope using the Olympus Soft Imaging System iTEM and Mega-ViewIII camera. Mitochondria were analyzed in triplicate from two control lines and one line from each patient. At least 15 fields of view were analyzed per line for every replicate.
Oxygen consumption measurements
SH-SY5Y cells were permeabilized with 40Mdigitonin and
subsequent re-suspension in a hypotonic medium containing 135 mMKCl, 10 mMNaCl, 20 mMHEPES, 0.5 mMKH2PO4, 1 mMMgCl2, 5 mMEGTA, and 1.86 mMCaCl2, pH 7.1. Oxygen consumption was measured in the Clark-type oxygen electrode (Hansatech) thermostatically maintained at 25 °C. Glutamate (5 mM), malate (5 mM), and inhibitors were indicated were added to the chamber before the measurement took place. To assess V3, 50 nmol of ADP was added and all data were obtained using the Oxygraph Plus system with chart recording software.
Fura-2 imaging
Calcium measurements using fura-2 were generated using an epifluorescence inverted microscope equipped with a⫻20 flu-orite objective (33). Changes in cytosolic calcium levels were monitored in single cells using excitation light provided by a Xenon arc lamp, the beam passing monochromator at 340 and 380 nm (Cairn Research, UK). The emitted fluorescence light was reflected through a 515-nm long pass filter to a cooled CCD camera (Retiga, QImaging, Canada) and digitized to 12-bit res-Table 2
Primers used for qPCR
Gene Forward Primer Reverse Primer Product
GAPDH atgacatcaagaaggtggtg cataccaggaaatgagcttg 177 bp CYCLOPHILIN ggcaaatgctggaccaaacac ttcctggacccaaaacgctc 147 bp
-ACTIN tcaccaccacggccgagcg tctccttctgcatcctgtcg 351 bp OCT4 ttctggcgccggttacagaacca gacaacaatgaaaatcttcaggaga 218 bp SOX2 catggcaatcaaaatgtcca tttcacgtttgcaactgtcc 119 bp NANOG gcttgccttgctttgaagca ttcttgactgggaccttgtc 256 bp ANT1 aagatcttcaagtctgatgg ataatatcggcccctttc 263 bp
ANT2 gcttgtcttgtatgatgaa aatagttgccactgagaa 145 bp
DNMT3b tttagggagaacgggaat agcaccagtaagaagagt 88 bp
at ROYAL VET COLL./LIBRARIAN on July 24, 2018
http://www.jbc.org/
olution. All imaging data were collected and analyzed using software from Andor (Belfast, UK).
Kinetic ATP measurements
To determine the ATP levels, patient-derived iPSC and neurons were transfected with a mitochondrial ATP probe (mitAT1.03), which was generated by Imamuraet al.(12) using Effectine (Qiagen) according to the manufacturer’s instruc-tions. Confocal images were obtained using a Zeiss 710 VIS CLSM microscope equipped with a META detection system and a⫻40 oil immersion objective. The FRET was quantified by the 527:475 nm ratio with an excitation of 405 nm and a filter from 515 to 580 nm. ATP kinetics were assessed in permeabi-lized iPSC and iPSC-derived neurons. To permeabilize the transfected cells, a buffer (0.137MNaCl, 5 mMKCl, 0.7 mM,
NaH2PO4, 25 mM, Tris-HCl, pH 7.1) containing a low concen-tration of digitonin (20M) was added to the cells. Upon per-meabilization, the buffer containing digitonin was replaced by fresh buffer containing mitochondrial substrates (5 mM
gluta-mate and 5 mMmalate). Cells were allowed to rest for at least 5
min before measurements took place. Basal mitochondrial ATP levels were recorded for 1 min before 500 nmol of ADP/ATP was added to the imaging chamber (500l final volume). Oli-gomycin (2g/ml) was added as a negative control at the end of the ADP experiments.
Statistical analysis
Statistical analysis and exponential curve fitting were per-formed using Origin 9 (Microcal Software Inc., Northampton, MA) software. Experimental data are shown as column scat-ter ⫾ S.D. n represents experiments undertaken. Statistical analysis between samples was performed using a one-way anal-ysis of variance with Bonferroni correction. Differences were considered to be significantly different ifp⬍0.05 (*).
Ethics approval and consent to participate
Informed consent was obtained from all patients in this study. All experimental protocols were carried out according to approved guidelines and regulations by the National Hospital for Neurology and Neurosurgery and the Institute of Neurology joint research ethics committee (09/0272).
Author contributions—M. H. R. L., C. A., F. B., M. D. V., and E. P. performed the experiments; M. H. R. L., C. A., and F. B. analyzed the data. A. Y. A., S. W., E. C., H. H. contributed reagents/materials/ analysis tools. M. H. R. L., C. A., F. B., A. Y. A., S. W., S. G., and H. H wrote the paper. All authors read and approved the final manuscript.
Acknowledgments—We thank Dr. Imamura (Kyoto University, Japan) for the mitAT1.03 construct. We also thank Kerrie Venner from the EM imaging unit at the National Hospital for Neurology and Neurosurgery for technical expertise.
References
1. Yin, H. Z., Nalbandian, A., Hsu, C. I., Li, S., Llewellyn, K. J., Mozaffar, T., Kimonis, V. E., and Weiss, J. H. (2012) Slow development of ALS-like spinal cord pathology in mutant valosin-containing protein gene knock-in mice.Cell Death Dis.3,e374
2. Weihl, C. C., Miller, S. E., Hanson, P. I., and Pestronk, A. (2007) Trans-genic expression of inclusion body myopathy associated mutant p97/VCP causes weakness and ubiquitinated protein inclusions in mice.Hum. Mol. Genet.16,919 –928
3. Nalbandian, A., Llewellyn, K. J., Badadani, M., Yin, H. Z., Nguyen, C., Katheria, V., Watts G., Mukherjee, J., Vesa, J., Caiozzo, V., Mozaffar, T., Weiss, J. H., and Kimonis, V. E. (2013) A progressive translational mouse model of human valosin-containing protein disease: the VCP(R155H/⫹) mouse.Muscle Nerve47,260 –270
4. Bartolome, F., Wu, H. C., Burchell, V. S., Preza, E., Wray, S., Mahoney, C. J., Fox, N. C., Calvo, A., Canosa, A., Moglia, C., Mandrioli, J., Chiò, A., Orrell, R. W., Houlden, H., Hardy, J., Abramov, A. Y., and Plun-Favreau, H. (2013) Pathogenic VCP mutations induce mitochondrial uncoupling and reduced ATP levels.Neuron78,57– 64
5. Nalbandian, A., Llewellyn, K. J., Gomez, A., Walker, N., Su, H., Dunnigan, A., Chwa, M., Vesa, J., Kenney, M. C., and Kimonis, V. E.(2015) In vitro studies in VCP-associated multisystem proteinopathy suggest altered mi-tochondrial bioenergetics.Mitochondrion22,1– 8
6. Plun-Favreau, H., Burchell, V. S., Holmström, K. M., Yao, Z., Deas, E., Cain, K., Fedele, V., Moisoi, N., Campanella, M., Miguel Martins, L., Wood, N. W., Gourine, A. V., and Abramov, A. Y. (2012) HtrA2 deficiency causes mitochondrial uncoupling through the F1F0-ATP synthase and consequent ATP depletion.Cell Death Dis.3,e335
7. Okita, K., Matsumura, Y., Sato, Y., Okada, A., Morizane, A., Okamoto, S., Hong, H., Nakagawa, M., Tanabe, K., Tezuka, K., Shibata, T., Kunisada, T., Takahashi, M., Takahashi, J., Saji, H., and Yamanaka, S. (2011) A more efficient method to generate integration-free human iPS cells.Nat. Meth-ods8,409 – 412
8. Shi, Y., Kirwan, P., Smith, J., Robinson, H. P., and Livesey, F. J. (2012) Human cerebral cortex development from pluripotent stem cells to func-tional excitatory synapses.Nat. Neurosci.15,477– 486, S1
9. Edri, R., Yaffe, Y., Ziller, M. J., Mutukula, N., Volkman, R., David, E., Jacob-Hirsch, J., Malcov, H., Levy, C., Rechavi, G., Gat-Viks, I., Meissner, A., and Elkabetz, Y. (2015) Analysing human neural stem cell ontogeny by con-secutive isolation of Notch active neural progenitors.Nat. Commun.6,
6500
10. Meyer, H., Bug, M., and Bremer, S. (2012) Emerging functions of the VCP/p97 AAA-ATPase in the ubiquitin system. Nat. Cell. Biol. 14,
117–123
11. Nalbandian, A., Llewellyn, K. J., Kitazawa, M., Yin, H. Z., Badadani, M., Khanlou, N., Edwards, R., Nguyen, C., Mukherjee, J., Mozaffar, T., Watts, G., Weiss, J., and Kimonis, V. E. (2012) The homozygote VCP(R(1) (5)(5)H/R(1)(5)(5)H) mouse model exhibits accelerated human VCP-as-sociated disease pathology.PLoS ONE7,e46308
12. Imamura, H., Nhat, K. P., Togawa, H., Saito, K., Iino, R., Kato-Yamada, Y., Nagai, T., and Noji, H. (2009) Visualization of ATP levels inside single living cells with fluorescence resonance energy transfer-based genetically encoded indicators.Proc. Natl. Acad. Sci. U.S.A.106,15651–15656 13. Ludtmann, M. H., Angelova, P. R., Ninkina, N. N., Gandhi, S., Buchman,
V. L., and Abramov, A. Y. (2016) Monomeric␣-synuclein exerts a physi-ological role on brain ATP synthase.J. Neurosci.36,10510 –10521 14. Krauss, S., Zhang, C. Y., and Lowell, B. B. (2005) The mitochondrial
un-coupling-protein homologues.Nat. Rev. Mol. Cell Biol.6,248 –261 15. Brand, M. D., Pakay, J. L., Ocloo, A., Kokoszka, J., Wallace, D. C., Brookes,
P. S., and Cornwall, E. J. (2005) The basal proton conductance of mito-chondria depends on adenine nucleotide translocase content.Biochem. J. 392,353–362
16. Kokoszka, J. E., Waymire, K. G., Levy, S. E., Sligh, J. E., Cai, J., Jones, D. P., MacGregor, G. R., and Wallace, D. C. (2004) The ADP/ATP translocator is not essential for the mitochondrial permeability transition pore.Nature 427,461– 465
17. Vieira, H. L., Haouzi, D., El Hamel, C., Jacotot, E., Belzacq, A. S., Brenner, C., and Kroemer, G. (2000) Permeabilization of the mitochondrial inner membrane during apoptosis: impact of the adenine nucleotide transloca-tor.Cell Death Differ.7,1146 –1154
18. Zhivotovsky, B., Galluzzi, L., Kepp, O., and Kroemer, G. (2009) Adenine nucleotide translocase: a component of the phylogenetically conserved cell death machinery.Cell Death Differ.16,1419 –1425
at ROYAL VET COLL./LIBRARIAN on July 24, 2018
http://www.jbc.org/
19. Papkovskaia, T. D., Chau, K. Y., Inesta-Vaquera, F., Papkovsky, D. B., Healy, D. G., Nishio, K., Staddon, J., Duchen, M. R., Hardy, J., Schapira, A. H., and Cooper, J. M. (2012) G2019S leucine-rich repeat kinase 2 causes uncoupling pro-tein-mediated mitochondrial depolarization.Hum. Mol. Genet.21,4201–4213 20. Weydt, P., Pineda, V. V., Torrence, A. E., Libby, R. T., Satterfield, T. F.,
Lazarowski, E. R., Gilbert, M. L., Morton, G. J., Bammler, T. K., Strand, A. D., Cui, L., Beyer, R. P., Easley, C. N., Smith, A. C., Krainc, D.,et al.
(2006) Thermoregulatory and metabolic defects in Huntington’s disease transgenic mice implicate PGC-1␣in Huntington’s disease neurodegen-eration.Cell Metab.4,349 –362
21. Johnson, A. E., Shu, H., Hauswirth, A. G., Tong, A., and Davis, G. W. (2015) VCP-dependent muscle degeneration is linked to defects in a dy-namic tubular lysosomal networkin vivo.Elife4,10.7554/eLife.07366 22. Weinbach, E. C., Sheffield, H., and Garbus, J. (1963) Restoration of
oxida-tive phosphorylation and morphological integrity to swollen, uncoupled rat liver mitochondria.Proc. Natl. Acad. Sci. U.S.A.50,561–568 23. Ding, W. X., Li, M., Biazik, J. M., Morgan, D. G., Guo, F., Ni, H. M.,
Goheen, M., Eskelinen, E. L., and Yin, X. M. (2012) Electron micro-scopic analysis of a spherical mitochondrial structure.J. Biol. Chem. 287,42373– 42378
24. Ganote, C. E., and Armstrong, S. C. (2003) Effects of CCCP-induced mi-tochondrial uncoupling and cyclosporin A on cell volume, cell injury and preconditioning protection of isolated rabbit cardiomyocytes.J. Mol. Cell. Cardiol.35,749 –759
25. Bobba, A., Amadoro, G., Petragallo, V. A., Calissano, P., and Atlante, A. (2013) Dissecting the molecular mechanism by which NH2htau and A1– 42 peptides impair mitochondrial ANT-1 in Alzheimer disease.
Biochim. Biophys. Acta1827,848 – 860
26. Schirris, T. J., Ritschel, T., Herma Renkema, G., Willems, P. H., Smeitink, J. A., and Russel, F. G. (2015) Mitochondrial ADP/ATP exchange inhibi-tion: a novel off-target mechanism underlying ibipinabant-induced myo-toxicity.Sci. Rep.5,14533
27. Abramov, A. Y., and Duchen, M. R. (2010) Impaired mitochondrial bio-energetics determines glutamate-induced delayed calcium deregulation in neurons.Biochim. Biophys. Acta1800,297–304
28. Muqit, M. M., Abou-Sleiman, P. M., Saurin, A. T., Harvey, K., Gandhi, S., Deas, E., Eaton, S., Payne Smith, M. D., Venner, K., Matilla, A., Healy, D. G., Gilks, W. P., Lees, A. J., Holton, J., and Revesz, T.,et al.(2006) Altered cleavage and localization of PINK1 to aggresomes in the presence of proteasomal stress.J. Neurochem98,156 –169
29. Sposito, T., Preza, E., Mahoney, C. J., Setó-Salvia, N., Ryan, N. S., Mor-ris, H. R., Arber, C., Devine, M. J., Houlden, H., Warner, T. T., Bushell, T. J., Zagnoni, M., Kunath, T., Livesey, F. J., Fox, N. C.,et al.(2015) Developmental regulation of tau splicing is disrupted in stem cell-derived neurons from frontotemporal dementia patients with the 10⫹ 16 splice-site mutation in MAPT.Hum. Mol. Genet.24,5260 –5269 30. Wray, S., Self, M., NINDS Parkinson’s Disease iPSC Consortium, NINDS
Huntington’s Disease iPSC Consortium, NINDS ALS iPSC Consortium, Lewis, P. A., Taanman, J. W., Ryan, N. S., Mahoney, C. J., Liang, Y., Devine, M. J., Sheerin, U. M., Houlden, H., Morris, H. R., Healy, D.,et al.(2012) Creation of an open-access, mutation-defined fibroblast resource for neu-rological disease research.PLoS ONE7,e43099
31. Brown, S., Teo, A., Pauklin, S., Hannan, N., Cho, C. H., Lim, B., Vardy, L., Dunn, N. R., Trotter, M., Pedersen, R., and Vallier, L. (2011) Ac-tivin/Nodal signaling controls divergent transcriptional networks in human embryonic stem cells and in endoderm progenitors.Stem Cells 29,1176 –1185
32. Vallier, L., Touboul, T., Brown, S., Cho, C., Bilican, B., Alexander, M., Cedervall, J., Chandran, S., Ahrlund-Richter, L., Weber, A., and Pedersen, R. A. (2009) Signaling pathways controlling pluripotency and early cell fate decisions of human induced pluripotent stem cells.Stem Cells27,
2655–2666
33. Angelova, P. R., Ludtmann, M. H., Horrocks, M. H., Negoda, A., Cre-mades, N., Klenerman, D., Dibson, C. M., Wood, N. W., Pavlov, E. V., Ghandhi, S., and Abramov, A. Y. (2016) Calcium is a key factor in␣ -sy-nuclein induced neurotoxicity.J. Cell Sci.129,1792–1801
34. Shi, Y., Kirwan, P., and Livesey, F. J. (2012) Directed differentiation of human pluripotent stem cells to cerebral cortex neurons and neural net-works.Nat. Protoc.7,1836 –1846
at ROYAL VET COLL./LIBRARIAN on July 24, 2018
http://www.jbc.org/
Abramov
Elisavet Preza, Eva Carro, Henry Houlden, Sonia Gandhi, Selina Wray and Andrey Y.
Marthe H. R. Ludtmann, Charles Arber, Fernando Bartolome, Macarena de Vicente,
neurons
across the mitochondrial membrane and impair energy metabolism in human
doi: 10.1074/jbc.M116.762898 originally published online March 30, 2017 2017, 292:8907-8917.
J. Biol. Chem.
10.1074/jbc.M116.762898
Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted
•
When this article is cited
•
to choose from all of JBC's e-mail alerts
Click here
http://www.jbc.org/content/292/21/8907.full.html#ref-list-1
This article cites 34 references, 7 of which can be accessed free at
at ROYAL VET COLL./LIBRARIAN on July 24, 2018
http://www.jbc.org/