O R I G I N A L R E S E A R C H
Coronin 3 Promotes the Development of
Oncogenic Properties in Glioma Through the
Wnt/
β
-Catenin Signaling Pathway
This article was published in the following Dove Press journal: OncoTargets and Therapy
Min Wang1 Qi Li1
Shengyuan Yu1 Zexiang Zhang2 Peng Qiu1 Yubao Zhang1 Wei Yang1 Guangming Xu1 Tongjiang Xu1
1Department of Neurosurgery, Shandong
Provincial Hospital Affiliated to Shandong First Medical University, Jinan, Shandong 250014, People’s Republic of China;
2Department of Gastrointestinal Surgery,
Shandong Provincial Hospital Affiliated to Shandong First Medical University, Jinan, Shandong 250014, People’s Republic of China
Purpose: Evidence indicates that the actin-binding protein Coronin 3, which is aberrantly expressed in various cancers, is associated with cancer development and progression. However, little is known about the role of Coronin 3 in glioma tumorigenesis. Here, we aimed to explore the biological function and regulatory mechanism of Coronin 3 in glio-blastoma (GBM).
Materials and Methods:Coronin 3 level in human GBM clinical samples and cell lines was investigated. The shRNA knockdown strategy was used to assess the tumor character-istics of GBM cell lines. The role ofβ-catenin in Coronin 3-mediated oncogenic phenotypes was evaluated.
Results: Coronin 3 was found to be highly upregulated in glioma cell lines. Furthermore, knockdown of Coronin 3 significantly inhibited the growth of glioma cells both in vivo and in vitro and suppressed the expression of Wnt/β-catenin pathway genes, includingβ-catenin, Cyclin D1, and c-Myc. Moreover, we demonstrated that Coronin 3 regulates the expression ofβ-catenin in glioma. Our results revealed that Coronin 3-stimulated tumor growth was β -catenin-dependent.
Conclusion: Our study reveals a new molecular mechanism of Coronin 3 in promoting glioma growth and development through regulating the Wnt/β-catenin signaling pathway.
Keywords:glioblastoma, Coronin 3, proliferation, oncogenic property, Wnt/β-catenin
Introduction
Glioblastoma (GBM), the most aggressive form of malignant glioma, shows poor prognosis and a median survival time of fewer than 15 months.1In contrast to other solid tumors, GBM is characterized by diffuse infiltrative growth around the normal brain parenchyma.2The biological behavior of gliomas is determined by the rate of GBM cell proliferation, invasion, and tumor migration.3However, the mechanisms involved in these processes remain elusive. In an effort to develop potential effective targeted therapies, recent studies have focused on understanding the molecular pathogenesis underlying glioma formation and progression.
Coronin is an actin-binding protein, and the proteins belonging to the coronin family differ both in terms of structure and function.4Functional studies have shown that it can regulate multiple cellular responses.5Coronin 3 (Coronin 1C, CRN2) is a member of the coronin protein family that is expressed ubiquitously—localized to lamellipodia—which reportedly interacts with the actin-related protein 2/3 complex.6,7 In recent years, aberrant expression of Coronin 3 has been identified to be responsible
Correspondence: Tongjiang Xu Department of Neurosurgery, Shandong Provincial Hospital Affiliated to Shandong First Medical University, Jinan, Shandong 250014, People’s Republic of China Tel +86-531-87938911
Email xutongjiangtj@163.com
OncoTargets and Therapy
Dove
press
open access to scientific and medical research
Open Access Full Text Article
OncoTargets and Therapy downloaded from https://www.dovepress.com/ by 118.70.13.36 on 27-Aug-2020
for the metastatic behavior of many malignancies, such as head and neck squamous cell carcinoma,8breast cancer,9and hepatocellular carcinoma.10 Breast cancer patients and cell lines exhibited increased Coronin 3 expression, which was associated with cancer cell invasion and increased metastatic risk.11Upregulation of Coronin-1C enhanced the migration of human HCC cells, whereas suppressed cell migration and proliferation were observed in Coronin-1C-knockdown cells.12Although only a few studies on Coronin 3 alteration in gliomas have been reported, Coronin 3 overexpression detected in glioma samples correlated with the degree of malignancy.13 However, the oncogenic properties of Coronin 3 and its molecular mechanisms in glioma cells have not been fully understood.
Wnt/β-catenin signaling pathway has been reported to regulate many aspects of cell behavior, such as cell-fate decision, cell migration, and cell differentiation.14,15Upon stabilization, active β-catenin accumulates in the cytosol and is translocated to the nucleus, where it forms a transcription complex with T cell factor (TCF)/lymphoid enhancer factor (LEF) to activate the transcription of down-stream target genes.16 Wnt/β-catenin has been reported to promote GBM cell migration and metastasis,17 yet the detailed mechanism of how Coronin 3 activates the onco-genic signaling pathway remains unknown. Therefore, we aimed to ascertain Coronin-3 expression in gliomas and to provide substantial evidence for the role of Coronin-3 in the growth and invasion of glioma cells and the underlying molecular mechanisms. The results of this study are expected to contribute to the understanding of the functions of Coronin-3 in GBM tumorigenesis.
Materials and Methods
Human Tissue Specimens and Cell Lines
The GBM tissues and specimens used in this study were obtained by surgical resection from 23 patients at the Neurosurgery Department of Shandong Provincial Hospital from 2017 to 2019, and the study was approved by the Shandong Provincial Hospital Ethics Committee. Histopathological diagnoses and grading of the glioma sam-ples were independently verified by two neuropathologists, based on WHO classification of tumors (2007). None of the patients had received chemotherapy, immunotherapy, or radiotherapy prior to specimen collection. There were five cases of Grade I, seven cases of Grade II, six cases of Grade III, and five cases of Grade IV glioma specimens. Five samples of non-tumor brain tissue were removed during
intracranial decompression surgery. The samples were immediately frozen and stored at–80 °C until analysis. The normal human astrocyte (NHA) cell and human glioblastoma cell lines (LN-18, A-172, U87, SNB19, LN-229, and U251MG) were purchased from the cell bank of the Chinese Academy of Sciences (Shanghai, China). Cells were cultured in Dulbecco’s modified Eagle’s medium (DMEM; Thermo Scientific, Waltham, MA, USA) supple-mented with 10% fetal bovine serum (FBS; Thermo Scientific, Waltham, MA, USA) and 100 U/mL penicillin (Invitrogen Life Technologies, Carlsbad, CA, USA) at 37 °C in an atmosphere containing 5% CO2.
Immunohistochemistry (IHC)
IHC was performed on polyformalin-fixed and paraffi n-embedded glioma tissues and healthy brain tissues. Tissue sections were incubated with primary antibodies against Coronin 3 (ab1571; 1:200; Abcam), β-catenin (9562S; 1:300; CST), and Ki67 (ab15580; 1:200; Abcam) at 4 °C overnight, followed by incubation with an HRP-conjugated secondary antibody. Diaminobenzidine was used for color reactions and images were acquired using a brightfield microscope (Leica, Germany).
Western Blotting
Total protein from tissues and cells was extracted using RIPA buffer containing 1% phenylmethylsulphonyl fl uor-ide. Protein concentration was determined before loading using a BCA protein assay kit (Beyotime Bio Corp, China). Equal amounts of protein (20μg) were loaded onto a 12% SDS polyacrylamide gel. The membrane was blocked using 5% bovine serum albumin (BSA) and probed with primary antibodies against Coronin 3 (ab1571; 1:200; Abcam, MA, USA) overnight at 4 °C and then incubated with an HRP-conjugated secondary antibody at 25 °C for 1 h. Finally, protein bands were visualized using the ECL detecting system (Applygen, Beijing). The integrated density values were normalized against those of GAPDH.
Plasmid Construction and Transfection
Scrambled negative control (SC) or Coronin 3 shRNA1/2 were purchased from Addgene (Beijing, China) and cloned into the pLKO.1 vector. The plasmid construct encoding human Coronin 3, was prepared by PCR amplification and subsequently subcloned into the pCDH-CMV-MCS-EF1-copGFP construct designed by Addgene (Beijing, China). 293FT cells were transfected with shRNA plasmids or over-expression plasmids. Lipofectamine 2000 (Life Technologies,
OncoTargets and Therapy downloaded from https://www.dovepress.com/ by 118.70.13.36 on 27-Aug-2020
USA) was used for all transfections according to the manu-facturer’s instructions. Forty-eight hours after transfection, the culture supernatant was collected and used to infect GBM cells. Transfections were confirmed by Western blotting before various experiments.
Cell Growth Curve Analysis
Cell proliferation assay was performed using Cell Counting Kit-8 (CCK-8) (Beyotime Institute of Biotechnology, Shanghai, China) according to the manufacturer’s instructions. Briefly, 2 × 106cells were seeded in each well of a 96-well plate and were cultured overnight. The CCK-8 reagent was added at 24, 48, 72, 96, or 120 h and incubated at 37 °C for 1 h. Each assay was independently repeated three times in triplicate.
Immuno
fl
uorescence
The cells werefixed with 4% paraformaldehyde at 37 °C for 1 h, permeabilized with 0.5% Triton X-100 for 10 min, and blocked in 5% BSA for 1 h at 25 °C. Then, the cells were probed with 5% BSA containing primary antibody anti-β -catenin (MAB8814, CST) overnight at 4 °C, followed by incubation with 488-conjugated secondary antibody (Zymed Laboratories, San Francisco, CA, USA) for 1 h at room temperature in the dark. After washing with 0.1M PBS for three times, the cells were stained with DAPI (H-1200,Vector laboratories). Then, cells were mounted on glass slides, and the fluorescence was visualized by laser scanning confocal microscopy (Olympus, Tokyo, Japan).
EdU Staining
Cells were seeded in 96-well plate and incubated with 50 μM of 5-ethynyl-2′-deoxyuridine (EdU, RiboBio; R11053), according to the manufacturer’s instructions. Subsequently, the DNA content was stained with DAPI and visualized under a laser scanning confocal microscope (Olympus, Tokyo, Japan).
Colony Formation Assay
Cells were seeded in 6-well plates (1 × 103cells/well) and cultured in DMEM with 10% FBS. After 6 days, the cells were washed with phosphate-buffered saline, fixed with methanol, stained with 1% crystal violet, and observed under a microscopy (Olympus, Tokyo, Japan).
Wound Scratch Assay
Cells were seeded in a 6-well plate, and after reaching 90% confluence, the cell monolayer was scraped with a pipette tip,
and re-incubated in with serum-free medium. Images were acquired at 0 h and 24 h after wounding using an inverted microscopy (Olympus, Tokyo, Japan) to assess wound closure.
Cell Migration and Invasion Assay
Cell migration and invasion were detected using the Transwell assay (Beyotime, Shanghai, China). Cells were seeded on uncoated (for migration assays) or Matrigel-coated (for inva-sion assays) upper chambers (BD Bioscience, New Jersey, USA). For the migration assay, the lower chamber wasfilled with a medium containing 10% FBS. For the invasion assay, cells were seeded in a serum-free medium in the upper cham-ber, which was pre-coated with Matrigel. Next, the lower chamber wasfilled with the complete medium. After 24 h of incubation at 37 °C, the cells werefixed with 4% paraformal-dehyde and stained with 0.1% crystal violet. Eight random visual fields were counted per chamber using an inverted microscope (Olympus, Tokyo, Japan).
T-Cell Factor (TCF)-Responsive
Luciferase Reporter Assay
The effect of Coronin 3 on Wnt/β-catenin signaling was detected using a pair of luciferase reporter constructs, as pre-viously described.18 Briefly, TOPflash and FOPflash (Millipore) were used; TOPflash contains 3 TCF/LEF sites
—upstream of a thymidine kinase promoter—and a firefly luciferase gene, while the FOPflash construct contains 3 mutated TCF/LEF sites, and is used as a control for measur-ing nonspecific reporter activation (Promega, Madison, WI). Luciferase activity was measured using a dual-luciferase assay system kit (Promega), according to the manufacturer’s proto-col, with theRenilla reniformis(sea pansy) luciferase activity as an internal control. Relative luciferase activity was expressed as normalized-fold change to controls.
Glioma Xenograft Mouse Models
Approximately 5 × 105 active U87 cells infected with lentiviruses containing shRNA or empty vector were resuspended in 200 μL PBS and subcutaneously injected into groups of seven nude mice. After 35 days of injection, the mice were euthanized, and xenografts were excised, weighed, and photographed.
Hematoxylin and Eosin Staining
The xenografts were cut in small pieces to ensure thorough
fixation. Next, the tumors werefixed in 4% paraformaldehyde
OncoTargets and Therapy downloaded from https://www.dovepress.com/ by 118.70.13.36 on 27-Aug-2020
for 24 h, stored in 70% ethanol overnight, and embedded in paraffin. Next, 5-μm-thick sections were prepared with a microtome (Leica, Germany) and deparaffinized with 100% xylene, and rehydrated using descending ethanol series. Finally, the sections were stained with hematoxylin and eosin (Sangon Biotech).
Statistical Analysis
Statistical analyses were performed using GraphPad Prism version 7.0 for Windows (San Diego, CA, USA). The data are presented as the mean ± SD or mean ± SEM. The Student’s t-test was used to compare the test and control groups, and comparisons between the two groups were determined by the least significant difference using the one-way analysis of variance method. P< 0.05 indicated a significant difference.
Results
Coronin 3 is Highly Expressed in GBM
and Human GBM Cell Lines
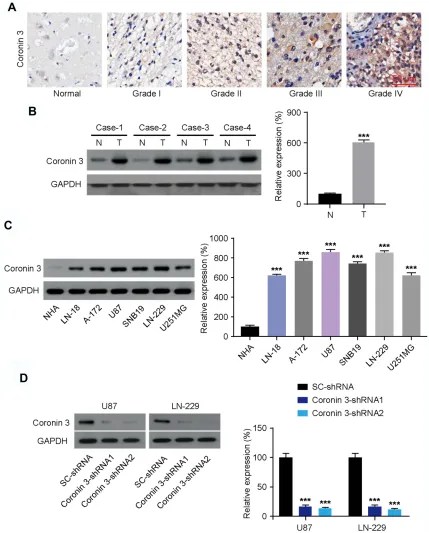
Human GBM clinical samples—including glioma tissues (Grade I–IV) and non-tumor healthy brain tissues—were collected to examine Coronin 3 expression by IHC. The results confirmed that Coronin 3 expression was higher in the GBM tissues than in healthy tissues, and that its expression increased with the GBM histological grade (Figure 1A). Concordantly, Western blotting revealed that the expression of Coronin 3 was higher in human GBM tissues (n=4) than that in the paired adjacent non-tumor tissues (Figure 1B). We further investigated the level of Coronin 3 in different subtypes of GBM cell lines. Western blotting revealed that Coronin 3 expression was markedly upregulated in all the tested GBM cell lines compared with that in primary NHA (Figure 1C). Collectively, these results indicate that Coronin 3 is upre-gulated in GBM.
Knockdown of Coronin 3 in GBM Cells
Results in Decreased Cell Proliferation
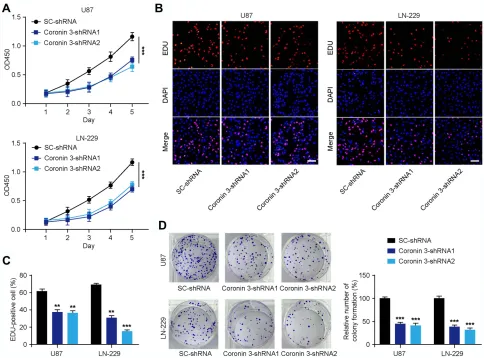
To further assess the role of Coronin 3 in GBM cells, U87 and LN-229 cell lines with stable shRNA knockdown of Coronin 3 were generated. The transfection efficacy of Coronin 3-shRNA constructs was verified by Western blotting (Figure 1D). CCK-8 assay was used to examine the effect of Coronin 3 knockdown on the proliferative ability of GBM cells. The results revealed that the growth rate of U87 and LN-229 cells transfected with Coronin
3-shRNA was significantly decreased compared with that of the scrambled control (Figure 2A). The EdU incorpora-tion assay was performed to examine the effect of Coronin 3 knockdown on DNA replication, as a more specific evaluation of proliferation. U87 and LN-229 cells trans-fected with Coronin 3-shRNA revealed a significantly reduced number of EdU-positive cells compared with that in cells transfected with the scrambled control (Figure 2BandC). Concordantly, colony formation assay showed that knockdown of Coronin 3 could suppress the proliferation of U87 and LN-229 cells (Figure 2D).
Knockdown of Coronin 3 Inhibits GBM
Cell Invasion and Migration in vitro
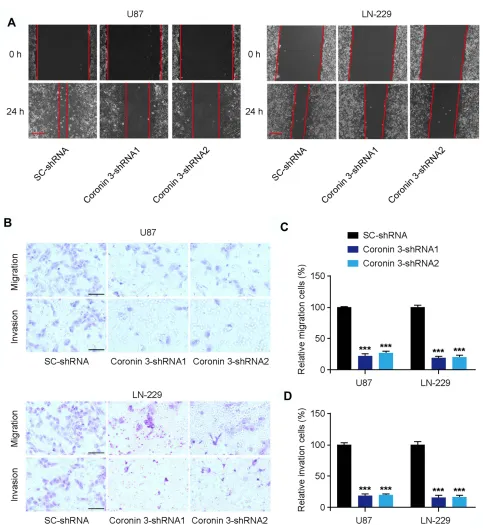
To further investigate the effect of Coronin 3 on GBM cell invasion and migration, we employed a wound healing assay and a Transwell invasion assay. The wound area was much smaller in SC-shRNA-transfected U87 and LN-229 cells compared with that in Coronin 3-shRNA-transfected cells 24 h after wounding, indicating that Coronin 3 promoted GBM cell migration (Figure 3A). Moreover, the downregulation of Coronin 3 inhibited the migration and invasion of U87 and LN-229 cells, as deter-mined by the Transwell assay (Figure 3B–D). These results suggest that the downregulation of Coronin 3 inhi-bits the invasion and migration of GBM cells.
The Signaling Basis for Coronin 3
Function in Tumorigenesis
Next, we investigated the molecular mechanism underlying the tumor-promotive role of Coronin 3. Upon downregula-tion of Coronin 3, the protein levels of MMP-9, integrinβ1, cyclin D1, c-Myc, andβ-catenin, which are the key signal-ing intermediaries of the canonical Wnt pathway, were significantly decreased (Figure 4A–F). These changes were corroborated by a marked decrease in the
immuno-fluorescence ofβ-catenin in Coronin 3-shRNA-transfected U87 and LN-229 cells compared to that in SC-shRNA-transfected cells (Figure 4G).
Coronin 3 Regulates the Expression of
β
-Catenin
To thoroughly test the relationship between Coronin 3 and
β-catenin, we obtained U87 and LN-229 cells stably expres-sing pcDNA-Coronin 3. We observed that the overexpression of Coronin 3 remarkably increased the expression of
β-catenin protein, compared with that in the control cells
OncoTargets and Therapy downloaded from https://www.dovepress.com/ by 118.70.13.36 on 27-Aug-2020
(Figure 5AandB). Furthermore, the effect of Coronin 3 on Wnt/β-catenin signaling was detected using a pair of lucifer-ase reporters containing TCF sites. Dual-luciferlucifer-ase reporter assay also revealed that Coronin 3 overexpression
significantly promoted the activity of firefly luciferase that carried wild-type but not mutant TCF binding sites in U87 as well as LN-229 cells (Figure 5C and D). These findings indicated that Coronin 3 drives the expression ofβ-catenin.
Figure 1Coronin 3 is overexpressed in glioblastoma (GBM) cells. (A) Immunohistochemistry for Coronin 3 in GBM (WHO Grade I–IV) and non-tumor (normal) tissues.
Scale bar represents 50μm. (B) Western blotting for Coronin 3 in four matched GBM tissues (T) and adjacent normal brain tissues (N) (n = 8). Coronin 3 expression is
normalized to that of GAPDH. ***P< 0.001. (C) Western blotting for Coronin 3 in GBM cell lines compared with that in normal human astrocytes (NHA) (n = 6). Coronin
3 expression is normalized to that of GAPDH. ***P< 0.001 vs NHA. (D) Western blotting for evaluating the efficiency of Coronin 3 knockdown in U87 and LN-229 cells
(n = 6). Coronin 3 expression is normalized to that of GAPDH. ***P< 0.001 vs SC-shRNA. One-way ANOVA was used to analyze the data, which are shown as the mean ±
SEM.
OncoTargets and Therapy downloaded from https://www.dovepress.com/ by 118.70.13.36 on 27-Aug-2020
Coronin 3 Exerts Its Oncogenic Effect by
Promoting the Wnt/
β
-Catenin Signaling
Pathway
The role of β-catenin in the development of Coronin 3-induced oncogenic phenotypes was evaluated. First, res-cue experiments with an activator of the Wnt/β-catenin pathway—lithium chloride (LiCl)19—were performed to verify the involvement of the Wnt/β-catenin signal in Coronin 3-induced oncogenic phenotypes. The results showed that the reduced growth rate of Coronin 3-shRNA1-transfected U87 cells was reversed after LiCl treatment (Figure 6A). Besides, the metastatic ability of U87 cells was enhanced in response to LiCl treatment, as evidenced by the results of the Transwell assay (Figure 6B). Next, we examined whether blocking the activity ofβ-catenin with the β-catenin inhibitor IWP-2,20 could result in the
attenuation of the effect of Coronin 3 upregulation. As expected, Coronin 3 overexpression promoted the prolifera-tion, migraprolifera-tion, and invasion of U87 cells. However, treat-ment with the β-catenin inhibitor IWP-2 significantly attenuated the promotive effect of Coronin 3 overexpres-sion on the proliferation, migration, and invaoverexpres-sion of U87 cells (Figure 6Cand D). Taken together, ourfindings sug-gest that Coronin 3 promotes the development of oncogenic phenotypes in GBM cells, partly via promoting Wnt/β -catenin signaling.
Knockdown of Coronin 3 Inhibits GBM
Growth in vivo
To explore the effects of Coronin 3 on tumor growth in vivo, U87 cells stably transfected with Coronin 3-shRNA or SC-shRNA were subcutaneously injected into nude mice
Figure 2Knockdown of Coronin 3 inhibits the proliferation of glioblastoma (GBM) cells in vitro. (A) Effect of Coronin 3 knockdown on cell proliferation at different time
points in U87 and LN-229 cells (n = 7). ***P< 0.001 vs shRNA. (B) Cell proliferation analyzed by EdU incorporation in U87 and LN-229 cells transfected with
SC-shRNA and Coronin 3-SC-shRNA. Scale bar represents 50μm. (C) Percentage of EdU-positive cells (n = 16 visionfields). **P< 0.01, ***P< 0.001 vs SC-shRNA. (D) Effect of
Coronin 3 knockdown on colony formation by cells (n = 18 visionfields). ***P< 0.001 vs SC-shRNA. One-way ANOVA was used to analyze the data, which are shown as
the mean ± SD.
OncoTargets and Therapy downloaded from https://www.dovepress.com/ by 118.70.13.36 on 27-Aug-2020
to establish xenograft models. The tumors formed in mice transfected with Coronin 3-shRNA grew significantly slower than those in SC-shRNA-transfected mice, indicat-ing that Coronin 3 knockdown inhibited tumor growth (Figure 7AandB). We then performed HE and Ki67 stain-ing on tumors from each group of mice with xenograft
tumors. H&E staining revealed decreased mitosis in tumors from mice injected with cells stably expressing Coronin 3-shRNA (Figure 7C). Ki67 expression was significantly weaker in tumors derived from mice injected with cells stably expressing Coronin 3-shRNA (Figure 7C). These results are consistent with those in vitro, which firmly
Figure 3Coronin 3 knockdown suppresses glioblastoma (GBM) cell migration and invasion in vitro. (A) Wound healing assay was used to evaluate the motility of GBM cell lines.
The representative images of migration were acquired 24 h after wounding. Scale bar represents 100μm (n = 15 visionfields). (B) Transwell cell migration and invasion assays were
used to investigate the effect of Coronin 3 knockdown on GBM cell migration and invasion. Scale bar represents 50μm. (CandD) Quantification of U87 and LN-229 cells showing
migration and invasion (n = 12 visionfields). ***P< 0.001 vs SC-shRNA. One-way ANOVA was used to analyze the data, which shown as the mean ± SD.
OncoTargets and Therapy downloaded from https://www.dovepress.com/ by 118.70.13.36 on 27-Aug-2020
validated the oncogenic role of Coronin 3 in the tumorigen-esis of GBM. To further investigate whether the Wnt/β -catenin pathway plays a vital role in the Coronin 3-induced tumor growth in vivo, we performed IHC, and found that the downregulation of Coronin 3 reduced the expression of
β-catenin (Figure 7CandD). A summaryfigure illustrates the keyfindings of the study (Figure 7E).
Discussion
Coronin 3 displays highly co-operative binding to actin
filaments; it is a transcriptionally dynamic gene whose expression is known to be aberrantly upregulated in multi-ple types of clinically aggressive cancer.21 Its expression
was reportedly increased in diffuse hepatocellular carci-noma and was known to be correlated with the degree of tumor malignancy.12,22 Coronin 3 is also known to be highly expressed in gastric cancer tissue and to be posi-tively correlated with tumor metastasis.23Concordant with these reports, Coronin 3 knockdown is known to result in significantly attenuated migration of HeLa and HEK 293 cells.24However, there are very few studies on the dereg-ulation of Coronin 3 in gliomas, and its functional mechanism in GBM is elusive. Thal et al first reported that the expression of Coronin 3 in glioma tissues was significantly higher than that in healthy brain tissues, and that it was related to malignancy.13 Ziemann et al further
Figure 4Wnt/β-catenin pathway was downregulated in Coronin 3-knocked down glioblastoma (GBM) cells. (A) Representative Western blots of MMP-9, integrinβ1, cyclin
D1, c-Myc, andβ–catenin. (B–F) Quantification of MMP-9, integrinβ1, cyclin D1, c-Myc, andβ-catenin expression, where expression is normalized to that of GAPDH
(n = 6). ***P< 0.001 vs SC-shRNA. (G) Immunofluorescence andfluorescent signal intensity analysis forβ-catenin in U87 and LN-229 cells. Nuclei were labeled by DAPI.
***P< 0.001 vs SC-shRNA. Scale bar represents 100μm. One-way ANOVA was used to analyze the data, which are shown as the mean ± SD.
OncoTargets and Therapy downloaded from https://www.dovepress.com/ by 118.70.13.36 on 27-Aug-2020
proved that Coronin 3 promotes GBM progression via modulation of the actin cytoskeleton.25 However, there are other plausible mechanisms to explain the GBM growth observed in this study, and these should therefore be targeted in the future to develop novel therapeutic approaches.
In our study, we focused on the regulatory mechanisms of Coronin 3 in GBM. Our results showed that Coronin 3 was markedly overexpressed in malignant gliomas, and that its expression level were positively correlated to tumor grade, which confirmed that Coronin 3 might be associated with the development and progression of glio-mas; this suggests that Coronin 3 functions as an onco-gene. Western blotting demonstrated that the expression of Coronin 3—in the tumor tissue as well as GBM cell lines—was increased, which indicates that the expression of Coronin 3 is correlated with the growth and invasive-ness of tumor cells. Therefore, we hypothesized that the
downregulation of Coronin 3 might serve as a potential strategy for glioma treatment.
Reorganization of the actin cytoskeleton is the primary mechanism underlying the motility of cells and is essential for most types of cell migration.26 Coronin 3 has been reported to be actively involved in mediating cell migration and progression.27 In the present study, Coronin 3 knock-down using shRNA transfection resulted in significantly reduced Coronin 3 expression in glioma cells, as indicated by Western blotting. In vitro, Coronin 3 downregulation resulted in significantly inhibited proliferation, migration, and invasion, which suggested that Coronin 3 expression was associated with the metastatic potential of glioma. Next, we demonstrated that the knockdown of Coronin 3 resulted in the downregulation of the Wnt/β-catenin signal-ing pathway by reducsignal-ing the expression of the pathway intermediaries at the protein level, suggesting that Wnt/β -catenin pathway is regulated by Coronin 3 in gliomas.
Figure 5Coronin 3 interacts directly withβ-catenin. (AandB) Coronin 3 overexpression markedly enhanced the and protein level ofβ-catenin in U87 and LN-229 cells.
GAPDH served as an internal control (n = 7). (CandD) U87, and LN-229 cells were transfected withfirefly luciferase reporter plasmids containing either wild-type or
mutant TCF binding sites. Thefirefly luciferase activity of each sample was normalized to the Renilla luciferase activity (n = 8). **P< 0.01 vs pc-DNA-Con, ***P< 0.001 vs
pc-DNA-Con. Thet-test was used to analyze the data, which are shown as the mean ± SD.
OncoTargets and Therapy downloaded from https://www.dovepress.com/ by 118.70.13.36 on 27-Aug-2020
Concordantly, the dual-luciferase reporter assay revealedβ -catenin to be a direct target of Coronin 3, which indicated that Coronin 3 knockdown inhibits the Wnt/β-catenin signal path-way viaβ-catenin.
Wnt/β-catenin signaling pathway is a major pathway that is activated during the carcinogenesis of glioma.28This path-way contributes to GBM pathology at multiple levels, includ-ing tumor initiation, maintenance of stem-cell status, invasion, and angiogenesis.29,30 Although GBMs do not harbor genetic alterations in the components of the Wnt/β -catenin pathway, aberrant activation of Wnt signaling appears to be achieved mainly via epigenetic silencing of the negative regulators of the Wnt/β-catenin pathway and overexpression of the positive regulators.31 Coronin 3 has been shown to promote the development of neuroblastomas, but the mechanism in gliomas has not been elucidated fully. Therefore, we speculated that Coronin 3 played a role in the development of glioma by regulating the Wnt/β-catenin sig-naling pathway. This study identified Coronin 3 as a positive regulator of the Wnt/β-catenin signaling pathway in GBM.
We confirmed our hypothesis by observing the tumor pheno-types in response to LiCl-induced activation of the Wnt signaling pathway in Coronin 3-shRNA-transfected GBM cells, and IWP-2-induced inhibition of the Wnt signaling in pc-DNA-Coronin 3-transfected GBM cells. Several actin-binding proteins have been reported to regulate the actin dynamics around the E-cadherin–β-catenin complex.32,33 As a conserved actin-binding protein that governs the cellular actin dynamics, Coronin 3 may also be modulated by β -catenin to regulate actinfilament assembly and disassembly. At the cell-cell contacts,β-catenin also links the cytoplasmic domain of cadherin-type adhesion receptors to Coronin 3,34 which allows the cells to interact via robust intercellular adhesion junctions. These potential interaction mechanisms between Coronin 3 and β-catenin should be further investigated.
Conclusion
In summary, our study suggests that Coronin 3 expression might serve as a significant independent prognostic factor
Figure 6Wnt/β-catenin signaling is involved in mediating Coronin 3-induced promotion of oncogenic properties. (A) Treatment with LiCl (30 mM) reversed the
anti-proliferation effect of Coronin 3-shRNA. ***P< 0.001 vs SC-shRNA.##
P< 0.01 vs Coronin 3-shRNA (n = 8). (B) Treatment with LiCl reversed the migration and
anti-invasion effect of Coronin 3-shRNA. ***P< 0.001 vs SC-shRNA.###P< 0.001 vs Coronin 3-shRNA (n = 16 visionfields). (C) Treatment withβ-catenin inhibitor IWP-2
(1μM) attenuated the promotive effect of pcDNA-Coronin 3 on proliferation. ***P< 0.001 vs pcDNA-Con.##
P< 0.01 vs pcDNA-Coronin 3 (n = 8). (D) Treatment with
β-catenin inhibitor IWP-2 attenuated the pro-migration and pro-invasion effect of pcDNA-Coronin 3. ***P< 0.001 vs pcDNA-Con.###P< 0.001 vs pcDNA-Coronin
3 (n = 16 visionfields). One-way ANOVA was used to analyze the data, which are shown as the mean ± SD.
OncoTargets and Therapy downloaded from https://www.dovepress.com/ by 118.70.13.36 on 27-Aug-2020
Figure 7Knockdown of Coronin 3 suppressed glioma growth in vivo. U87 cells transfected with Coronin 3-shRNA or SC-shRNA were injected into nude mice (n = 8 per group). (A) Tumors from the xenograft were assessed for volume after 35 days of injection. (B) Images of the dissected tumors. (C) Representative images of H&E staining
and immunohistochemistry of Ki-67 andβ-catenin are shown. Scale bar represents 100μm. (D) Semi-quantitative analysis ofβ-catenin positive rate (n = 12 sections from
3 mice). (E) A summaryfigure to illustrate keyfindings of the study. ***P< 0.001 vs SC-shRNA. One-way ANOVA was used to analyze the data, which are shown as the
mean ± SD.
OncoTargets and Therapy downloaded from https://www.dovepress.com/ by 118.70.13.36 on 27-Aug-2020
of high-grade glioma. Further, Coronin 3 promotes glioma cell proliferation, invasion and migration, thereby leading to the activation of the Wnt/β-catenin signal pathway and promoting the development of glioma phenotypes in vitro and in vivo. Our results suggest that Coronin 3 plays an important role in gliomagenesis and may serve as a potential target for glioma therapy.
Abbreviations
GBM, glioblastoma; TCF, T cell factor; LEF¸ lymphoid enhancer factor.
Data Sharing Statement
We declare that materials described in the manuscript, including all relevant raw data, will be freely available upon reasonable request.
Funding
This work was supported by the Shandong Provincial Natural Science Foundation, China (Grant No. ZR2015HM015) and the Science and Technology Development Plan Project of Jinan City (Grant No. 201302038).
Disclosure
The authors declare that they have no competing interests.
References
1. Ferguson SD, Lariviere MJ, Mansour N, Lesniak MS. Malignant glioma: chemovirotherapy.Tumors Cent Nerv Syst.2011;1:357–364. 2. D’Alessandro G, Catalano M, Sciaccaluga M, Chece G, Limatola C.
KCa3.1 channels are involved in the infiltrative behavior of glioblastoma in vivo.Cell Death Dis.2013;4:e773. doi:10.1038/cddis.2013.279 3. Cheng YC, Tsai WC, Sung YC, Chang HH, Chen Y. Interference with
PSMB4 expression exerts an anti-tumor effect by decreasing the inva-sion and proliferation of human glioblastoma cells. Cell Physiol Biochem.2018;45:819–831. doi:10.1159/000487174
4. Roadcap DW, Clemen CS, Bear JE. The role of mammalian coronins in development and disease.Subcell Biochem.2008;48:124–135. 5. Clemen CS, Rybakin V, Eichinger L. The coronin family of proteins.
Sub Cell Biochem.2008;48:1–5.
6. Rosentreter A, Hofmann A, Xavier CP, Stumpf M, Noegel AA, Clemen CS. Coronin 3 involvement in F-actin-dependent processes at the cell cortex. Exp Cell Res. 2007;313:878–895. doi:10.1016/j. yexcr.2006.12.015
7. Lizaka M, Han H-J, Akashi H, et al. Isolation and chromosomal assignment of a novel human gene, CORO1C, homologous to coronin-like actin-binding proteins. Cytogen Cell Gene. 2000; 88:221–224. doi:10.1159/000015555
8. Walk EL, Hong W, Garcia MV, Bear JE, Weed SA. Abstract 4752: cortactin and coronin 1B cooperate to promote tumor cell invasion in head and neck squamous cell carcinoma.Cancer Res.2011;71:4752–4762.
9. Wang J, Tsouko E, Jonsson P, et al. Abstract P4-07-12: miR-206 inhibits cell migration through direct targeting of the actin-binding protein coronin 1C in triple-negative breast cancer. Cancer Res. 2013;73:1690–1702.
10. Wu L, Peng CW, Hou JX, et al. Coronin-1C is a novel biomarker for hepatocellular carcinoma invasive progression identified by proteo-mics analysis and clinical validation. J Exp Clin Cancer Res. 2010;29:17. doi:10.1186/1756-9966-29-17
11. Castagnino A, Castro-Castro A, Marie Irondelle M, et al. Coronin 1C promotes triple-negative breast cancer invasiveness through regula-tion of MT1-MMP traffic and invadopodia function. Oncogene. 2018;37:6425–6441. doi:10.1038/s41388-018-0422-x
12. Wang ZG, Jia MK, Cao H, Bian P, Fang XD. Knockdown of Coronin-1C disrupts Rac1 activation and impairs tumorigenic poten-tial in hepatocellular carcinoma cells. Oncol Rep. 2012;29:1066‒ 1072.
13. Thal D, Xavier CP, Rosentreter A, et al. Expression of coronin-3 (coronin-1C) in diffuse gliomas is related to malignancy.J Pathol. 2008;214:415–424. doi:10.1002/path.2308
14. Luu H, Zhang R, Haydon R, et al. Wnt/β-catenin signaling pathway as novel cancer drug targets.Curr Cancer Drug Targets.2004;4:653–671. doi:10.2174/1568009043332709
15. Wang X, Meng X, Sun X, et al. Wnt/β-catenin signaling pathway may regulate cell cycle and expression of cyclin A and cyclin E protein in hepatocellular carcinoma cells. Cell Cycle. 2009;8:1567–1570. doi:10.4161/cc.8.10.8489
16. Leung JY, Kolligs FT, Wu R, Zhai Y, Fearon ER. Activation of AXIN2 expression by beta-catenin-T cell factor. A feedback repressor pathway regulating Wnt signaling. J Biol Chem. 2002;277:21657–21665. doi:10.1074/jbc.M200139200
17. Zhang L, Liu H, Mu X, Cui J, Peng Z. Dysregulation of Fra1 expression by Wnt/β-catenin signaling promotes glioma aggressive-ness through epithelial-mesenchymal transition.Biosci Rep.2017;37: BSR20160643. doi:10.1042/BSR20160643
18. Chen L, Li M, Li Q, Wang CJ, Xie SQ. DKK1 promotes hepatocel-lular carcinoma cell migration and invasion throughβ-catenin/MMP7 signaling pathway. Mol Cancer. 2013;12:157. doi:10.1186/1476-4598-12-157
19. Hao HP, Wen LB, Li JR, et al. LiCl inhibits PRRSV infection by enhancing Wnt/β-catenin pathway and suppressing inflammatory responses. Antivir Res. 2015;117:99–109. doi:10.1016/j.antiviral. 2015.02.010
20. Guo YZ, Xie XL, Fu J, Xing GL. SOX9 regulated proliferation and apoptosis of human lung carcinoma cells by the Wnt/β-catenin signal-ing pathway.Eur Rev Med Pharmacol Sci.2018;22(15):4898–4907. doi:10.26355/eurrev_201808_15626
21. Chan KT, Roadcap DW, Holoweckyj N, Bear JE. Coronin 1C har-bours a second actin-binding site that confers co-operative binding to F-actin.Biochem J.2012;444:89–96. doi:10.1042/BJ20120209 22. Wu L, Hou JX, Peng CW, et al. Increased coronin-1C expression is
related to hepatocellular carcinoma invasion and metastasis. Chin J Hepatol.2010;18:516.
23. Gui R, Qifei T, Yanxin A, et al. Coronin 3 promotes gastric cancer metastasis via the up-regulation of MMP-9 and cathepsin K. Mol Cancer.2012;11:67. doi:10.1186/1476-4598-11-67
24. Samarin SN, Koch S, Ivanov AI, Parkos CA, Nusrat A. Coronin 1C negatively regulates cell–matrix adhesion and motility of intestinal epithelial cells.BBRC.2010;391:0–400.
25. Ziemann A, Simon H, Ridhirama B, et al. CRN2 enhances the invasiveness of glioblastoma cells.Neuro Oncol.2013;5:548–561. 26. Yamazaki D, Kurisu S, Takenawa T. Regulation of cancer cell
moti-lity through actin reorganization. Cancer Sci. 2005;96:379–386. doi:10.1111/j.1349-7006.2005.00062.x
27. Sun Y, Shang Y, Ren G, et al. Coronin3 regulates gastric cancer invasion and metastasis by interacting with Arp2.Cancer Biol Ther. 2014;15:1163–1173. doi:10.4161/cbt.29501
28. Liu C, Tu Y, Sun X, et al. Wnt/beta-Catenin pathway in human glioma: expression pattern and clinical/prognostic correlations.Clin Exp Med.2011;11:105–112. doi:10.1007/s10238-010-0110-9
OncoTargets and Therapy downloaded from https://www.dovepress.com/ by 118.70.13.36 on 27-Aug-2020
29. Tao Q, Wu C, Xu R, et al. Diallyl trisulfide inhibits proliferation, invasion and angiogenesis of glioma cells by inactivating Wnt/β -catenin signaling.Cell Tissue Res.2017;370(3):379‒390. doi:10.1007/ s00441-017-2678-9
30. Wang G, Shen J, Sun J, et al. Cyclophilin A maintains glioma-initiating cell stemness by regulating Wnt/β-catenin signaling.Clin Cancer Res. 2017;23(21):6640‒6649. doi:10.1158/1078-0432.CCR-17-0774 31. Arnés M, Casas Tintó S. Aberrant Wnt signaling: a special focus in CNS
diseases.J Neurogenet.2017;31:1–7. doi:10.1080/01677063.2017.1338 696
32. Bahloul A, Simmler MC, Michel V, Leibovici M, Petit C. Vezatin, an integral membrane protein of adherens junctions, is required for the
sound resilience of cochlear hair cells. EMBO Mol Med.
2009;1:125–138. doi:10.1002/emmm.200900015
33. Lilien J, Balsamo J. The regulation of cadherin-mediated adhesion by tyrosine phosphorylation/dephosphorylation ofβ-catenin.Curr Opin Cell Biol.2005;17:459–465. doi:10.1016/j.ceb.2005.08.009 34. Gottardi CJ. Adhesion signaling via catenins. Bone. 2010;47:S18.
doi:10.1016/j.bone.2010.04.013
OncoTargets and Therapy
Dove
press
Publish your work in this journal
OncoTargets and Therapy is an international, peer-reviewed, open access journal focusing on the pathological basis of all cancers, potential targets for therapy and treatment protocols employed to improve the management of cancer patients. The journal also focuses on the impact of management programs and new therapeutic
agents and protocols on patient perspectives such as quality of life, adherence and satisfaction. The manuscript management system is completely online and includes a very quick and fair peer-review system, which is all easy to use. Visit http://www.dovepress.com/ testimonials.php to read real quotes from published authors.
Submit your manuscript here:https://www.dovepress.com/oncotargets-and-therapy-journal
OncoTargets and Therapy downloaded from https://www.dovepress.com/ by 118.70.13.36 on 27-Aug-2020