Food
for
thought:
how
nutrients
regulate
root
system
architecture
Zaigham
Shahzad
and
Anna
Amtmann
Thespatialarrangementoftheplantrootsystem(rootsystem architecture,RSA)isverysensitivetoedaphicandendogenous signalsthatreportonthenutrientstatusofsoilandplant. SignallingpathwaysunderpinningRSAresponsestoindividual nutrients,particularlynitrateandphosphate,havebeen unravelled.Researchershavenowstartedtoinvestigate interactiveeffectsbetweentwoormorenutrientsonRSA. Severalproteinsenablingcrosstalkbetweensignalling pathwayshaverecentlybeenidentified.RSAispotentiallyan importanttraitforsustainableand/ormarginalagriculture.Itis generallyassumedthatRSAresponsesareadaptiveand optimisenutrientuptakeinagivenenvironment,buthard evidenceforthisparadigmisstillsparse.Herewesummarize recentadvancesmadeintheseareasofresearch.
Address
InstituteofMolecular,CellularandSystemsBiology,CollegeofMedical,
Veterinary,andLifeSciences,UniversityofGlasgow,GlasgowG12
8QQ, United Kingdom
Correspondingauthor:Amtmann,Anna([email protected])
CurrentOpinioninPlantBiology2017,39:80–87
ThisreviewcomesfromathemedissueonCellsignallingandgene
regulation
EditedbyTzyy-JenChiouandToruFujiwara
http://dx.doi.org/10.1016/j.pbi.2017.06.008
1369-5266/#2017 The Authors. Published by Elsevier Ltd. This is an
openaccessarticleundertheCCBYlicense(
http://creativecom-mons.org/licenses/by/4.0/).
Introduction
Rootsarepivotalforanchorageoflandplantsinthesoiland for efficient uptakeof water and mineralnutrients,thus playingacrucialrolein plantfitness.Therootsystemis composedofembryonicroots(primaryrootinArabidopsis, primaryandseminalrootsinafewcerealcrops)and post-embryonicroots(lateralrootsinArabidopsis,lateral,brace andcrownrootsincereals).Rootsystemarchitecture(RSA), theoverallspatial arrangementof individualpartsof the rootsystem,isanimportantfactordetermininghow effi-cientlyplantscan accessresources.RSAishighlyplastic bothgeneticallyandenvironmentally.Thus,different spe-ciesorecotypeshaveevolveddifferentRSAsdependingon theprevailingsoilconditions.ConsiderablydifferentRSAs
canalsobeadoptedwithinthe samegenotypeandeven withinthe lifespanof asingleplantbecausetherateat whichindividualpartsoftherootsystemdevelopandgrow canbealteredbyshort-termenvironmentalsignals, includ-ingchangesinwater, nutrientandoxygen availabilityor pathogensandpests.Unlikeanimals,plantscannotmove awayfromunfavourablesites.Hence,plantresponses to fluctuatingsoilconditionsarebasedonalteredgrowthand development,andtheyrequiresophisticatedsensingand signallingmechanisms.Whilethemechanismscontrolling RSAresponsestoindividualnutrients,especiallynitrogen (N)andphosphorus(P),havebeenextensively investigat-ed,thecrosstalkbetweendifferentnutrientsignalsandthe benefitsofRSAresponsesinaparticularconditionareyetto be characterized systematically. In this review, we will highlightsomeoftherecentlyidentifiedmolecular mech-anismsthatunderpinRSA responsesto singleand com-bined nutrient stress,and we will explore the potential benefitsofRSAresponsesforplantperformance.
The
genetic
and
environmental
context
of
RSA
responses
MoststudiesintonutrientsignallingandRSAhavebeen carriedoutinArabidopsisthaliana(Col-0)grownonagar plates,andbeforewereviewtheknowledgegeneratedby thesestudies,itisimportanttodiscusstheir transferabil-itytootherspeciesandrootenvironments.The endoge-nous developmental programmes underpinning RSA differbetweenplantspecies(especiallybetween mono-cots and dicots) which is apparent in different RSAs underidenticalnutrientsufficientconditions.Therefore, aRSA phenotype producedinresponsetoa particular nutritionalstimuluscan differbetweenspecieseven if thesignallingpathwayfeedingintothedifferent devel-opmental programmes is conserved. Furthermore, the phenotypicconsequenceofachangeinnutrientsupply inagivengenotypewilldependonexactnutrient con-centration,nutrientdistributionand gradients, concen-trations ofother nutrients, developmentalstage ofthe plant,andanyfactorsthat determineplantgrowthrate andnutrientdemand,forexample,light,humidity,etc. [1,2,3].Differencesinthesefactorsarelikelytoexplain many ofthediscrepanciesbetweenstudiescarried out withthesamegenotype,forexample,A.thalianaCol-0. Clearlyonehastobecarefulwhendrawingwider con-clusionsfromobservationsmadeinaspecificgenotypeor environmentalcondition.However,thisdoesnotimply that meaningful data can only be derived from field-based studies on crops. Ultimately, whether findings can be translated between species and environments
ornotdependsonthequestionposed.IftheRSAitselfis thefocusofinterest(e.g.asatraittoenhanceagricultural yieldorasaphenotypetosolvespecificdevelopmental questions)thepreciseexperimentalcontextisofessence andfindingsmaynotbetransferabletoothergenotypes orenvironments.If,however,RSAresponsesareusedas readout toenabletheidentification ofnutrientsensors and signalling components, the context is less crucial, andartificialconditionsareoftenmoreinformativethan naturalconditions.Figure1exemplifiestheusefulnessof Arabidopsis RSA developing on an agar surface for reporting the action of nutrient-specific sensing and signalling pathways. Each specific nutrient treatment leadstoadistinctmultifactorialphenotypicoutputthat can be interrogated through genetic, pharmacological and physiological manipulations. In the following, we willdescribethelatestinsightsthathavebeenobtained usingthislatterapproach.
Root
system
architecture
responses
to
individual
nutrients
Nitrate
In Arabidopsisplants grownwith uniformnutrient sup-ply,thedose–effectcurvefornitrateandlateralroot(LR) lengthisbell-shaped,whereasprimaryroot(PR)lengthis
continuouslyinhibitedbyloweringnitrateoverthesame concentration range [4]. The effect of nitrate on LR initiationiscontroversial;severalstudiesreportapositive effect of nitrate onLR density [5,6] while others have foundnoeffectsofnitrateonLRnumberordensity[7]. Non-uniform, localized nitrate supply stimulates LR elongation innitraterichpatches[3].
NITRATETRANSPORTER1.1,NRT1.1(NPF6.3),is a plasma membrane nitrate transporter that alters its affinitydependingonnitrateavailability,andfunctions asanitratesensorupstreamoftranscriptionallow-nitrate responses[8].Italsoactsasabasipetalauxintransporter inthedevelopingLRtipstherebycontrollingbothlocal and systemic LR responses tolow nitrate [3,9]. Phos-phorylation ofNRT1.1 at T101is important forauxin transportactivityandforrepressionofLRemergencein lownitrate(Figure2)[10].Arecentstudy[11]resolved thelongstanding questionhowsuch afunctionis com-patible with theobserved transcriptional down-regula-tionofNRT1.1underlownitrate,bothatthewholeroot levelandinLRprimordia[3,12].Bouguyonand collea-gues showed that in contrast to NRT1.1-mRNA the NRT1.1-protein levels are increased in LR primordia ofnitratestarvedroots withaconcomitant decreaseof
Figure1
-NO3
-+NO3
-Control Nitrate
Phosphate
Potassium Calcium Sulphur
Iron Manganese
Boron Zinc
Nitrate, local
Magnesium
Current Opinion in Plant Biology
Schematic phenotypes of ArabidopsisthalianaCol-0 plants in single-nutrient deficiency. Representative phenotypes are shown for plants grown
on vertical agar plates with media lacking the specified nutrient (except for nitrate which was supplied at low level), based on published data
[1,2].Phenotypesinnutrient-sufficientmediaandphenotypesreportedforlocalizednitratetreatment[3]areshownforcomparison.Lengthof
primaryandlateralrootsisrepresentedbythelengthofthegreyspheres,andlateralrootdensityisrepresentedbytheirwidth.Greenovals
auxin accumulation in LR primordia. These findings affirmaroleofNRT1.1inthelow-nitrateRSAresponse, and the challenge now is to identify the mechanism regulating NRT1.1 proteinstability. Incontrast tothe welldocumentedstimulatoryeffectsoflocalizednitrate onLRgrowth,arecentstudy foundanincreaseinLR lengthunderuniformlow-nitrateconditions,whichwas dependentontheauxinbiosynthesisenzyme TRYPTO-PHANAMINOTRANSFERASERELATED2,TAR2 [13]. Nitrate was suppliedin the form ofNH4NO3 in contrast to previous studies where it was supplied as KNO3,thus making it difficultto distinguishbetween theeffectsofNH4andNO3.
One group of downstreamtargets of NRT1.1mediated N-signallingareANR1MADS-boxtranscriptionfactors, which positivelyregulate LR elongationin response to localized nitrate supply [14]. In a uniform low-nitrate
environment Arabidopsis plants overexpressing ANR1 exhibit longer LRs while no alteration in LR density andPRlengthwasobservedcomparedtowildtype[15]. Rice harboursfive ANR1-like MADS transcription fac-tors,fourofwhich(OsMADS 23,25,27,57and 61)are targetsofamicroRNA,miR144-a,whichisconservedin monocots and absent in Arabidopsis [16]. The overex-pression of miR144-ain rice reduces the expression of theseMADS-boxtranscriptionfactorsandabolishesLR stimulationbylocalizednitratetreatment,suggestingthat oneormoreofthesericeANR1-likegenesplayasimilar role to thatof ArabidopsisANR1. A rice homologue of ArabidopsisNRT1.1,NRT1.1B, wasrecently identified by QTL mapping of natural genetic variation of N signallingand N use efficiency [17].However, a com-prehensivecharacterizationoftheroleofriceNRT1.1B incontrollingrootdevelopmentalresponsestonitrateis still missing. Furthermore, it is also not known if
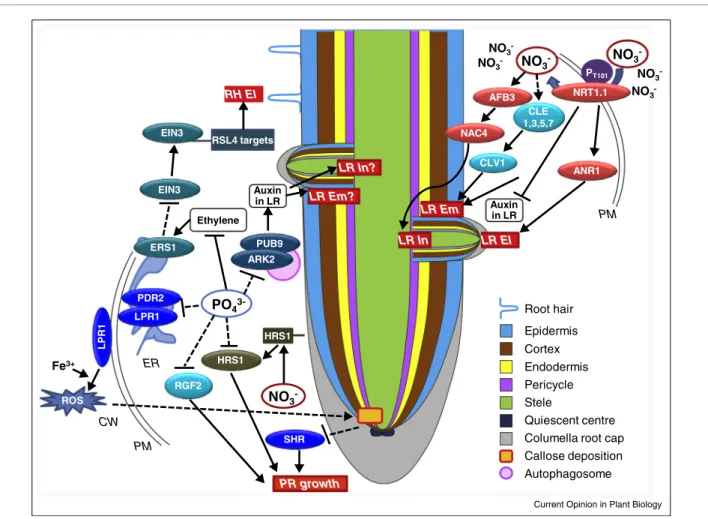
Figure2 PT101 CLE 1,3,5,7 CLV1 PDR2 LPR1 LP R 1 PM ER CW
Endodermis Epidermis Cortex Pericycle Stele
Quiescent centre Columellaroot cap Callose deposition PR growth RGF2 Autophagosome PUB9 ARK2 Auxin in LR ERS1 EIN3 RSL4 targets Root hair ROS Fe3+ HRS1 HRS1 NO3 -NO3 -NO3 -NO3 -NO3 -PM NO3 -NO3 -NRT1.1 ANR1 Auxin in LR AFB3 LR In LR Em NAC4 LR El SHR LR Em? LR In? RH El Ethylene PO4 3-EIN3
Current Opinion in Plant Biology
Summaryofsignallingpathwaysunderpinningrootsystemarchitectureresponsestonitrateorphosphatedeficiency.Componentsofsignalling
pathways discussed in this review are shown as coloured boxes (genes) or circles (proteins). Arrows and bars indicate positive and negative
regulation, respectively. Broken lines represent indirect pathways or circumstantial evidence. RSA targets (red boxes) are primary root (PR)
growth,lateralrootinitiation(LRIn),emergence(LREm)andelongation(LREl),aswellasroothairelongation(RHEl).Cellularlocalizationis
indicatedascellwall(CW),plasmamembrane(PM)orendoplasmicreticulum(ER).Roottissuesandotherfeaturesarecolouredasdetailedinthe
NRT1.1B acts upstream of rice ANR1-like genes to regulate theirexpressionin additiontothemiR144-a. AnessentialroleofsmallpeptidesinperceptionofNand alterationofrootdevelopmentwasrecentlydiscoveredin Arabidopsis.Inaheterogeneousnitrateenvironment, C-terminallyencodedpeptides(CEPs)aresecretedbythe N-starvedrootsandareconsideredto betranslocatedto theshoot wheretheyare perceivedbytwo leucine-rich repeat receptor kinases (CEPR1, CEPR2 LRK-RKs). This process triggers systemic N-demand signalling in rootsfacingnitraterichpatchesasevidencedbyanalysing themRNAregulationof NRTgenes [18].Inadditionto CEPpeptides,CLE(CLAVATA3/Endosperm surround-ing region-related) peptides were also detected in the xylemsap[19],andthissignalalsoappearstobeinvolved in the adjustment of root development to N-need (Figure 2) [4]. mRNAs of CLE peptides are induced by low nitrate andrepressed byhigh nitrate, and over-expressionofArabidopsisCLE1to7wasshowntoinhibit LR development under uniform low nitrate conditions [4]. In summary, there is increasing evidence for the involvementofpeptidesinNsignalling,butthe integra-tion of these signalling pathways with the NRT1.1-de-pendentsignallingpathwayremainsto beresolved. AdetaileddiscussionofhownitratemodulatesLR initi-ationandPRgrowthcanbefoundelsewhere[20].Briefly, thecurrentstateofknowledgesuggeststhatLRinitiation isregulatedbyaninteractionbetweennitrateandauxin signalling pathways and involves NRT1.1-dependent nitrateaccumulationthatinducestheAUXIN SIGNAL-LINGF-BOX 3, AFB3,and aNACdomain containing transcriptionfactor, NAC4, actingdownstream ofAFB3 (Figure 2)[6,21].
Phosphate
PhosphatestarvationstronglyinhibitsPR,stimulatesLR growth, and enhances root hair production. Numerous genetic components regulating root developmental responses to phosphate starvation have been identified [22].InArabidopsisCol-0LOWPHOSPHATEROOT1 (LPR1)andPHOSPHATEDEFICIENCYRESPONSE 2 (PDR2) appear to be central for sensing phosphate deprivation and triggering PR growth inhibition (Figure2)[23].LPR1,anditshomologueLPR2,encode ferroxidases whilePDR2encodes a P5-typeATPaseof unknown substrate specificity. The LPR1 and PDR2 proteins are expressed in thesame domains of theroot apical meristem (RAM) [24]. Phosphate starvation-in-duced PR growth inhibition is markedly reduced in
lpr1andlpr2singlemutantsandtheresponseis complete-lyabolishedinlpr1lpr2doublemutantplants.Bycontrast,
pdr2 mutantsexhibitahypersensitiveshort-root pheno-type under phosphate limitation [25,26]. Phosphate deprivedwildtypePRsshowincreasedradialcelldivision intherootmeristemanddisorganizationofthequiescent
centre[27].Phosphatedeficiencyalsoleadstoapoplastic callose deposition possibly due to enhanced ROS pro-ductioninstemcellnicheandRAM(Figure2),whichis more pronounced in pdr2 mutant and not evident in
lpr1lpr2mutantplants[26].Depositionofcallose inter-fereswiththecelltocellmovementofSHORTROOT HAIR(SHR),akeytranscriptionfactorregulatingradial rootpatterning,whichcouldexplaintheimpactonRAM maintenance. The lpr1lpr2pdr2 triple mutant plants do notshow callosedeposition or hypersensitiveshort-root phenotypeunderphosphatedeprivationwhichindicates that LPR1 works downstream of PDR2 in the same pathway [24,26]. Recent studies highlight that the impairmentofrootmeristematicactivityunderphosphate deficientconditionsisprecededbyanimpairmentofcell expansion in the root elongation zone, which is again LPR1dependent.ASENSITIVETOPROTON TOX-ICITY1-ALUMINUM-ACTIVATED MALATE TRANSPORTER1 (STOP1-ALMT1) module acts up-stream ofLPR1to controlthisearlyresponse[28,29]. Aroleofpeptidesignallingmechanismsisalsoemerging for low-P sensing and modulation of RSA (Figure 2). Arabidopsis mutant plants for RGF2,a member of the ROOT GROWTH FACTOR/GOLVEN/CLE-like (RGF/GLV/CLEL)familyofpeptides,showa hypersen-sitiveshort-rootphenotypeaftertransfertolow-Pmedia [27], indicating that RGF2 maintains longitudinal PR growth under phosphate deprivation. Mutant analysis furtherrevealedthatRGF2alsoenhancesradialdivisions inepidermis,cortex,andendodermis.Bycontrast, anoth-er RGF gene,RGF1,represses radialdivisionsin these tissues[27].ThissuggeststhatthedifferentRGF pep-tidesplay specificroles to modulateRSAunder P defi-ciency.Whetherandhowapeptidesignallingpathwayis integratedwith thePDR2-LPR1module remainsto be investigated.
A functional module comprising S-DOMAIN RECEP-TOR KINASE 1-6 (SDK6 (ARK2)) and its interacting partner,theU-box/ARMREPEATcontainingE3ligase AtPUB9, plays arole in LR developmentin phosphate deprived roots [30]. An Arabidopsis ark2pub9 double mutant exhibits severe reduction of phosphate starva-tion-inducedLRgrowthstimulation,whichisassociated withdecreasedauxinaccumulationinPRaswellasLR tipscomparedtowildtype(Figure2).ThePUB9protein is localized to the autophagosomes in the presence of ARK2indicatingapotentialroleofautophagyin stimu-latingLRformationunderphosphatedeficientconditions [30]. This notion was further supported by the lack of stimulationof LR formation whenP-starvedrootswere co-treated with 3-methyladenine (3-MA), an autophagy inhibitor.
As mentionedabove, phosphate starvation also induces roothair production.The molecularcomponentsofthis
response were recently identified through a large-scale genetic screen. A dominant negative mutation in the ethylene receptor ERS1, ‘hypersensitive to phosphate starvation 5’ (hsp5), resulted in a constitutive increase inthenumberandlengthofroothairs,andthephenotype was markedly pronounced under phosphate deficient conditions[31].Thismutationcausesaconstitutive eth-yleneresponseduetoenhancedproteinaccumulationof ETHYLENEINSENSITIVE3(EIN3),akey transcrip-tion factorinvolved in the ethylenesignalling pathway (Figure 2). Interestingly, EIN3 protein, which was also foundtobeinducedbyphosphatedeprivation,bindsto thedirecttargetsofROOTHAIRDEFECTIVE6-like4 (RSL4),atranscriptionfactorcontrollingroothair devel-opment,thustriggeringthesignallingpathwayregulating roothairdevelopment.
Othernutrients
Althoughrootsystemarchitecturalresponsesto deficien-cyofothernutrientsthanNand Phavebeenreported [1,2,30], very little is known about the mechanisms controllingthem.Ingeneral,atthemorphologicallevel, inhibitionofPR growthisacommonresponsetomost nutrientdeficiencies,exceptforsulphurandzinc. How-ever,theseverityofinhibitionvariesbetweennutrients. Amongstthemacronutrientsphosphateisbyfarthemost importantnutrientdeterminingPRlengthin Arabidop-sisCol-0[1],butinsomeotheraccessionspotassium(K) has astronger impact thanP [32]. Deficiency-induced LRresponsesvaryconsiderablybetweennutrients pro-ducingnutrient-specificpatternsofLRlength, density and branching [2]. In Col-0, K starvation leads to a stronginhibitionoffirst-orderLRlengthwhile branch-ingisstimulated,leadingtoincreasedsecond-orderLR density. The branching response is compromised in knockout mutants of AKT1, an inward rectifying K-channel, CIPK23, a CBL-interacting protein kinase, and NRT1.1 [1]. CIPK23 when bound to CBL1/9 phosphorylates AKT1 and enhances channel activity [33].NRT1.1isalsoatargetofCIPK23[8],whichmight explain its involvement in the K-starvation response. However,AKT1isnotthemainpathway forK-uptake under K-starvation, suggesting that its primary role in RSAregulationisthatofaK-sensor (‘transporter-recep-tor’),similartotheroleofNRT1.1inN-signalling.This viewis furthersupportedbythe findingthat the high-affinityK-uptaketransporterHAK5,whichiscrucialfor K-nutrition in low K, is not required for the RSA re-sponse[1].
Severalotherionsincludingmicronutrientsalsomodulate LR features (Figure 1) [2]. Starvation for calcium or boronincreasesfirst-orderLRdensitywithout significant-ly affecting LR length. Magnesium or iron starvation resultinareductionoffirst-orderLRdensityandlength. Manganese starvation inhibits LR length and slightly reducesbranching whereas zinc deficiencyalso inhibits
LR growth but stimulates LR branching. In summary, plants respond to deficiencies of most nutrients with nutrient-specific RSA phenotypes. The next challenge isnowto extendourknowledgeonsignallingpathways controllingRSAtonutrients otherthanPandN.
Responses
to
combined
nutrient
stress
Whetherplants growin thewild or in agriculture, they will often face multiple nutritional deficiencies. The importance of interactive effects of nutrients on RSA hasonly recentlybeen highlighted in astudy applying multiplecombinationsofsufficientandlowN,P,KandS, aswellashigh andlowlight,to Arabidopsisplants[1]. Varioustypesofantagonistic,synergistic,ornulleffectsof combined nutrient deficiencies as compared to single nutrient deficiencies were observed. For instance, S wasfoundtohaveinteractiveeffectsonRSAwithseveral othernutrientsdespitenotcausinganyRSAphenotypes insingedeficiency,whileKandNincreasedLR branch-inginsinglebutnotindoubledeficiency[1,2].Clearly, conventionalsingle-nutrientdeficiencyexperimentscan highlightonlyaverysmall partof thecomplexnutrient signallingnetwork that underpinsRSA in differentsoil conditions. Large-scale experiments would need to be carried out, usingamulti-dimensional design to enable quantificationofRSAinvariousconcentrationratiosofall essentialnutrients.At least for a subset of combined nutritional stresses perception and signalling crosstalk has recently been studied.Amolecularmechanismintegratingphosphate andnitratesignals wasdescribedforArabidopsisCol-0 [34].Amajorpartofthephosphatestarvation-induced PR inhibitionwas found tobe controlled bythe tran-scriptionfactorHYPERSENSITIVETOLOW Pi-ELI-CITEDPRSHORTENING(HRS1)anditshomologue HHO1,butonlywhennitratewaspresentinthegrowth media [34]. HRS1 and HHO1 are transcriptionally induced early during nitrate starvation inan NRT1.1-dependent manner,but HRS1 protein is stabilized by phosphate starvation (Figure 2). This indicates that transcriptional and post-transcriptional regulation of HRS1bynitrateandphosphaterespectivelyconstitutes amechanismtoexhibitanintegratedrootresponsetotwo distinctnutritionalsignals.
Cross-talkbetweenphosphateandheavymetalnutrients such as iron and zinc has long been appreciated. In particular, the phosphate starvation-induced inhibition of PR growthhas been shownto bedependent onthe availabilityof iron[35],which islikelyto beduetothe importanceofFe3+inthePDR2-LPR1signallingmodule [26].LPR1 proteinexhibitsdual sub-cellular localiza-tionintheendoplasmicreticulumandinthecellwall,but isconsideredto function primarilyasacellwall ferrox-idase.LPR1-dependentapoplasticFe3+accumulationin theroottipispotentiallyasourceofROSproduction.In
thelatestmodel,ROSaccumulationthenleadsto meri-stem-specific callose formationand restrictionof cellto cellmovement of SHR1,whichultimately impedesPR development(Figure 2,andabove).
Benefits
of
adjusting
RSA
to
nutrient
availability
RSAplasticityundernutrientstressisgenerallydescribed as an adaptive measure to enhance nutrient uptake through improved soil foraging [36].However, there is surprisingly little experimental evidence to prove this concept. Intra-species variation of RSAprovides an ex-cellentopportunitytoassesstheconsequencesof differ-entrootarchitecturesonnutrientuptakeefficiency,shoot growth and yield in different conditions in a similar geneticbackground.
In the caseofN, asteep, cheap anddeeproot system has been proposed to be ideal for N acquisition, be-cause nitrateis very mobile inthe soil [37]. This root modelwasindeed validatedbystudyingvarious geno-typesofmaize.Maize‘FL’genotypeswithfewbutlong LRs performed better under low-N than ‘MS’ geno-types with many but short LRs [37,38]. LR density was found to be negatively correlated with the root depth which was positively correlated with plant N content, shoot growth and grain yield under low N conditions. One possibleexplanation is that allocation of resources required forroot growthsuch assugars is more efficiently relocated in FL genotypes, thus de-creasingthecostofrootgrowthwhileimproving explo-ration of the soil for nitrate. Similarly, rice genotypes that showed less inhibition of root length exhibited higher nitrateuptake,nitrogenuseefficiencyandyield under low N conditions [39].
InthecaseofPdeficiency,atopsoil-foragingrootsystem is considered to be better thana deep root system as phosphate often accumulates in topsoil layers [40,41]. Indeed,inmaizegenotypeshigherLRbranchingdensity wascorrelatedwithhigheryieldonPdeficientsoils[42]. Evidencefortheimportanceoftotalrootlengthandroot surface area for P acquisition can also be found in phenotypes related to natural and transgenic alleles
of PHOSPHORUS STARVATION TOLERANCE 1
(PSTOL1), a gene underlying a major QTL of P-defi-ciencytolerance inrice[43].Furthermore, overexpres-sionofexpansins,forexample,GmEXPB2insoybeanor
TaEXPB23 in tobacco, increased LRnumber and im-proved phosphorus use efficiency [44–46]. Root hair elongation also contributes to P acquisition under P deficientconditionsasshownfortheroothair-less Ara-bidopsisline,NR23[47].Usinginbredlinesofcommon beandifferingforshallowbasalrootangleandroothair lengthanddensity,itwasfoundthatthetraits synergis-ticallyimprovedPacquisitionunderphosphate depriva-tion[48].
WhilethesestudiesstronglysupporttheimportanceofRSA fornutrientuptake andyield,theyaremostlycorrelative andoftenlimitedbypoorcontrolofthenutrientprofilein thesoilandlackofknowledgeondepletionkineticsinthe rhizosphere. Furthermore, some of the observed differ-ences may be based on differences in overall root size ratherthandifferencesinthespatialarrangementof indi-vidualrootparts.Toobtainabetterunderstandingofthe exactconsequences,costsandconstraintsofdifferentRSAs in different root environments it will be necessary to developanexperimentalsystemthatallowsprecise spatio-temporalcontrolandmonitoringofnutrientconcentrations intherhizospherewhileenablingcontinuousmeasurement ofRSAfeaturesandphysiologicalparameters.
Conclusions
Nutrient sensing and signalling continues to bean im-portantandactivefieldofresearch,andquantificationof root system architecture as a phenotypic output has provided excellent opportunitiesto identify crucial sig-nalling components. Overrecentyears novelty has par-ticular arisen from firstly, the integration of transcriptional, translational, redox and cell-wall based regulatory processes,secondly, thediscoveryof peptide signals inlocal andsystemicnutrient signalling,thirdly, theidentificationofmolecularhubsunderpinning inter-active effectsof differentnutrients,andfinally,the cor-relationofRSAwithnutrientuseefficiencyandyieldin crop genotypes. A concerted effort is now required to bettercontrolandmonitorthenutrientprofilearoundthe rootandtosystematicallymanipulateseveralnutrientsas wellasotherenvironmentalfactorsthatdeterminegrowth and development.Combinedwith genetics,particularly the exploration of natural variation, such a systemic approach could generate the necessary knowledge to generatemodelsthatcanpredictRSAforany genotype for anygivencombinationof nutrients.LinkingRSAto nutrientuptake,growthandyieldwouldbethenextstep. Itispossiblethatnutrient-useefficiencyundera moder-atefertilizerregimecouldbeincreasedbyde-sensitizing the plant for nutrient-deficiency signals. Explorationof this and other crop improvement strategies requires a precise understanding of how nutrient-signalling net-works arehard-wired.
Acknowledgement
ThisworkwasfundedbytheBBSRC(grantBB/N018508/1).
References
and
recommended
reading
Papers of particular interest, published within the period of review,
have been highlighted as:
of special interest
of outstanding interest
1.
KellermeierF,ArmengaudP,SeditasTJ,DankuJ,SaltDE, AmtmannA:Analysisoftherootsystemarchitectureof Arabidopsisprovidesaquantitativereadoutof crosstalkbetweennutritionalsignals.PlantCell2014, 26:1480-1496.
Thisstudysystematicallyrecordedtheeffectsofmultiplenutrient
com-binations on root system architecture of Arabidopsis, and usedthe
information to test the function of transporters and kinases in K–N
signalingcrosstalk.
2.
GruberBD,GiehlRFH,FriedelS,vonWire´nN:Plasticityofthe Arabidopsisrootsystemundernutrientdeficiencies.Plant Physiol2013,163:161-179.
The paper shows dose–effect curves for Arabidopsis root system
architectural features over nutrient concentration. Macronutrients
andmicronutrientswerevariedindividuallyinacommonbackground
solution.
3. MounierE,PerventM,LjungK,GonjonA,NacryP: Auxin-mediatednitratesignallingbyNRT1.1participatesinthe adaptiveresponseofArabidopsisrootarchitecturetothe spatialheterogeneityofnitrateavailability.PlantCellEnviron
2014,37:162-174.
4. ArayaT,MiyamotoM,WibowoJ,SuzukiA,KojimaS,TsuchiyaYN, SawaS,FukudaH,vonWire´nN,TakahashiH:CLE-CLAVATA1 peptide-receptorsignalingmoduleregulatestheexpansionof plantrootsystemsinanitrogen-dependentmanner.ProcNatl AcadSciUSA2014,111:2029-2034.
5. VidalEA,ArausV,LuC,ParryG,GreenPJ,CoruzziGM, Gutie´rrezRA:Nitrate-responsivemiR393/AFB3regulatory modulecontrolsrootsystemarchitectureinArabidopsis thaliana.ProcNatlAcadSciUSA2010,107:4477-4482.
6. VidalEA,MoyanoTC,RiverasE,Contreras-Lo´pezO,Gutie´rrezRA: Systemsapproachesmapregulatorynetworksdownstream oftheauxinreceptorAFB3inthenitrateresponseof
Arabidopsisthalianaroots.ProcNatlAcadSciUSA2013, 110:12840-12845.
7. ZhangH,JenningsA,BarlowPW,FordeBG:Dualpathwaysfor regulationofrootbranchingbynitrate.PlantBiol1999, 96:6529-6534.
8. HoCH,LinSH,HuHC,TsayYF:CHL1functionsasanitrate sensorinplants.Cell2009,138:1184-1194.
9. KroukG,LacombeB,BielachA,Perrine-WalkerF,MalinskaK, MounierE,HoyerovaK,TillardP,LeonS,LjungKetal.: Nitrate-regulatedauxintransportbyNRT1.1definesamechanismfor nutrientsensinginplants.DevCell2010,18:927-937.
10.
Bouguyon E, Brun F, Meynard D, Kubesˇ M, Pervent M, Leran S,
Lacombe B, Krouk G, Guiderdoni E, Zazˇı´malova´ E etal.: Multiple
mechanismsofnitratesensingbyArabidopsisnitrate transceptorNRT1.1.NatPlants2015http://dx.doi.org/10.1038/ nplants.2015.15.
TheauthorsdemonstratethatphosphorylationofNRT1.1controlslow
nitrate-inducedinhibitionofLRdevelopmentandthatphosphorylated
andnon-phosphorylatedformsofNRT1.1havedistinctfunctionsin
N-signalling.
11. BouguyonE,Perrine-WalkerF,PerventM,RochetteJ,CuestaC, BenkovaE,Martinie`reA,BachL,KroukG,GojonAetal.:Nitrate controlsrootdevelopmentthroughpost-transcriptional regulationoftheNRT1.1/NPF6.3transporter/sensor.Plant Physiol2016,172:1237-1248.
12. OkamotoM,VidmarJJ,GlassADM:RegulationofNRT1and NRT2genefamiliesofArabidopsisthaliana:responsesto nitrateprovision.PlantCellPhysiol2003,44:304-317.
13. MaW,LiJ,QuB,HeX,ZhaoX,LiB,FuX,TongY:Auxin biosyntheticgeneTAR2isinvolvedinlownitrogen-mediated reprogrammingofrootarchitectureinArabidopsis.PlantJ
2014,78:70-79.
14. RemansT,NacryP,PerventM,FilleurS,DiatloffE,MounierE, TillardP,FordeBG,GojonA:TheArabidopsisNRT1.1 transporterparticipatesinthesignalingpathwaytriggering rootcolonizationofnitrate-richpatches.ProcNatlAcadSciUS A2006,103:19206-19211.
15. GanY,BernreiterA,FilleurS,AbramB,FordeBG: OverexpressingtheANR1MADS-boxgeneintransgenic plantsprovidesnewinsightsintoitsroleinthenitrate regulationofrootdevelopment.PlantCellPhysiol2012, 53:1003-1016.
16. YanY,WangH,HameraS,ChenX,FangR:MiR444ahasmultiple functionsinthericenitrate-signalingpathway.PlantJ2014, 78:44-55.
17.
HuB,WangW,OuS,TangJ,LiH,CheR,ZhangZ,ChaiX, WangH,WangYetal.:VariationinNRT1.1Bcontributesto nitrate-usedivergencebetweenricesubspecies.NatGenet
2015,47:834-838.
The paper contributes to the translation of gene function from model to
crop. In this case allelic variation in a rice NRT1.1 homologue was shown
to correlate with nitrogen use efficiency and yield.
18. TabataR,SumidaK,YoshiiT,OhyamaK,ShinoharaH, MatsubayashiY:Perceptionofroot-derivedpeptidesbyshoot LRR-RKsmediatessystemicN-demandsignaling.Science
2014,346:343-346.
19. OkamotoS,SuzukiT,KawaguchiM,HigashiyamaT, MatsubayashiY:Acomprehensivestrategyforidentifying long-distancemobilepeptidesinxylemsap.PlantJ2015, 84:611-620.
20. FordeBG:Nitrogensignallingpathwaysshapingrootsystem architecture:anupdate.CurrOpinPlantBiol2014,21:30-36.
21. VidalEA,AlvarezJM,Gutie´rrezRA:NitrateregulationofAFB3 andNAC4geneexpressioninArabidopsisrootsdependson NRT1.1nitratetransportfunction.PlantSignalBehav2014, 9:e28501.
22. BouainN,DoumasP,RouachedH:Therootsystemresponseto phosphatedeficiencyinArabidopsis.CurrGenomics2016, 17:308-314.
23. HoehenwarterW,MonchgesangS,NeumannS,MajovskyP,
AbelS,MullerJ:Comparativeexpressionprofilingrevealsa
roleoftherootapoplastinlocalphosphateresponse.BMC PlantBiol2016http://dx.doi.org/10.1186/s12870-016-0790-8. 24. TicconiCA,LuceroRD,SakhonwaseeS,AdamsonAW,CreffA,
NussaumeL,DesnosT,AbelS:ER-residentproteinsPDR2and LPR1mediatethedevelopmentalresponseofrootmeristems tophosphateavailability.ProcNatlAcadSciUSA2009, 106:14174-14179.
25. SvistoonoffS,CreffA,ReymondM,Sigoillot-ClaudeC,RicaudL, BlanchetA,NussaumeL,DesnosT:Roottipcontactwith low-phosphatemediareprogramsplantrootarchitecture.Nat Genet2007,39:792-796.
26.
Mu¨llerJ,ToevT,HeistersM,TellerJ,MooreKL,HauseG, DineshDC,Bu¨rstenbinderK,AbelS:Iron-dependentcallose depositionadjustsrootmeristemmaintenancetophosphate availability.DevCell2015,33:216-230.
This paper reports a suite of elegant experiments dissecting the
mole-culareventsunderpinningprimaryrootinhibitioninresponsetoalow-P
signal.TheFe-dependentpathwayinvolvesanapoplasticmulticopper
oxidase, Fe-accumulation, ROS production and callose deposition,
whichrestrictstheintercellularmovementofakeytranscriptionfactor.
27.
CederholmHM,BenfeyPN:Distinctsensitivitiestophosphate deprivationsuggestthatRGFpeptidesplaydisparaterolesin
Arabidopsisthalianarootdevelopment.NewPhytol2015, 207:683-691.
ThestudyfoundthatRGFpeptidesregulate rootsystemarchitecture
underP-deficiency,andthatdifferentRGFisoformsdifferentiallyaffect
differentpartsoftherootsystem.
28. Mora-Macı´as J,Ojeda-RiveraJO,Gutie´rrez-Alanı´s D, Yong-VillalobosL,Oropeza-AburtoA,Raya-Gonza´lezJ, Jime´nez-Domı´nguez G,Cha´vez-CalvilloG,Rella´n-A´lvarezR, Herrera-EstrellaL:Malate-dependentFeaccumulationisacritical checkpointintherootdevelopmentalresponsetolow phosphate.ProcNatlAcadSciUSA2017,114:E3563-E3572.
29. Balzergue C, Dartevelle T, Godon C, Laugier E, Meisrimler C,
Teulon JM, Creff A, Bissler M, Brouchoud C, Hage`ge A etal.: Low
phosphateactivatesSTOP1-ALMT1torapidlyinhibitrootcell elongation. NatCommun2017http://dx.doi.org/10.1038/ ncomms15300.
30. DebS,SankaranarayananS,WewalaG,WiddupEE,SamuelMA: TheS-domainreceptorkinaseAtARK2andthe U-box/ARM-repeat-containingE3ubiquitinligaseAtPUB9module mediateslateralrootdevelopmentunderphosphate starvationinArabidopsis.PlantPhysiol2014,165:1647-1656.
31. SongL,YuH,DongJ,CheX,JiaoY,LiuD:Themolecular mechanismofethylene-mediatedroothairdevelopment inducedbyphosphatestarvation.PLoSGenet2016, 12:e1006194.
32. KellermeierF,ChardonF,AmtmannA:Naturalvariationof Arabidopsisrootarchitecturerevealscomplementing adaptivestrategiestopotassiumstarvation.PlantPhysiol2013, 161:1421-1432.
33. XuJ,LiHD,ChenLQ,WangY,LiuLL,HeL,WuWH:Aprotein kinase,interactingwithtwocalcineurinb-likeproteins, regulatesK+transporterAKT1inArabidopsis.Cell2006, 125:1347-1360.
34.
Medici A, Marshall-Colon A, Ronzier E, Szponarski W, Wang R,
Gojon A, Crawford NM, Ruffel S, Coruzzi GM, Krouk G: AtNIGT1/
HRS1integratesnitrateandphosphatesignalsatthe Arabidopsisroottip.NatCommun2015http://dx.doi.org/ 10.1038/ncomms7274.
TheauthorsshowedthatHRS1,aGARPtranscriptionfactor,integratesN
andPsignaling.HRS1inhibitsPRgrowthunderPdeficiency,butonlyin
thepresenceofnitrate.Interestingly,HRS1regulationbyNandPisatthe
leveloftranscriptorproteinrespectively.
35. StriederML,PintoKG,BertoldiC,SchneiderADB,DelatorreCA:
ResponseofArabidopsisthalianarootgrowthtophosphorus anditsrelationwithmediachemicalcomposition.BiolPlant
2017http://dx.doi.org/10.1007/s10535-017-0713-z.
36. TianH,DeSmetI,DingZ:Shapingarootsystem:regulating lateralversusprimaryrootgrowth.TrendsPlantSci2014, 19:426-431.
37. LynchJP:Steep,cheapanddeep:anideotypetooptimize waterandNacquisitionbymaizerootsystems.AnnBot2013, 112:347-357.
38.
ZhanA,LynchJP:Reducedfrequencyoflateralrootbranching improvesNcapturefromlow-Nsoilsinmaize.JExpBot2015, 66:2055-2065.
Thepaperprovidesanexperimentalvalidationofa‘steep,cheapand
deep’rootideotypeoptimizingNacquisitionfromthesoil.Maize
geno-types withfewandlongrootshadbetternitrogenuseefficiencythan
genotypeswithmanybutshortroots.
39. JuC,BureshRJ,WangZ,ZhangH,LiuL,YangJ,ZhangJ:Root andshoottraitsforricevarietieswithhighergrainyieldand
highernitrogenuseefficiencyatlowernitrogenrates application.FieldCropRes2015,175:47-55.
40. Paez-GarciaA,MotesC,ScheibleW-R,ChenR,BlancaflorE, MonterosM:Roottraitsandphenotypingstrategiesforplant improvement.Plants2015,4:334-355.
41. HeppellJ,TalboysP,PayvandiS,ZygalakisKC,FliegeJ, WithersPJA,JonesDL,RooseT:Howchangingrootsystem architecturecanhelptackleareductioninsoilphosphate(P) levelsforbetterplantPacquisition.PlantCellEnviron2015, 38:118-128.
42. PostmaJA,DatheA,LynchJP,ScienceP,PennsylvaniaJAP: Theoptimallateralrootbranchingdensityformaize dependsonnitrogenandphosphorus.PlantPhysiol2014, 166:590-602.
43. GamuyaoR,ChinJH,Pariasca-TanakaJ,PesaresiP,CatausanS, DalidC,Slamet-LoedinI,Tecson-MendozaEM,WissuwaM, HeuerS:TheproteinkinasePstol1fromtraditionalriceconfers toleranceofphosphorusdeficiency.Nature2012,488:535-539.
44. HanYY,ZhouS,ChenYH,KongX,XuY,WangW:The
involvementofexpansinsinresponsestophosphorus availabilityinwheat,anditspotentialsinimproving phosphorusefficiencyofplants.PlantPhysiolBiochem2014, 78:53-62.
45. GuoW,ZhaoJ,LiX,QinL,YanX,LiaoH:Asoybeanb-expansin geneGmEXPB2intrinsicallyinvolvedinrootsystem architectureresponsestoabioticstresses.PlantJ2011, 66:541-552.
46. ZhouJ,XieJ,LiaoH,WangX:Overexpressionofb-expansin geneGmEXPB2improvesphosphorusefficiencyinsoybean.
PhysiolPlant2014,150:194-204.
47. TanakaN,KatoM,TomiokaR,KurataR,FukaoY,AoyamaT, MaeshimaM:Characteristicsofaroothair-lesslineof
Arabidopsisthalianaunderphysiologicalstresses.JExpBot
2014,65:1497-1512.
48. MiguelMA,PostmaJA,LynchJP:Phenesynergismbetween roothairlengthandbasalrootgrowthangleforphosphorus acquisition.PlantPhysiol2015,167:1430-1439.