TABLE OF CONTENTS:
Vol. 44, No. 5, September 1991
ARTICLES Plant Ecology 418
422
421
434
438
443
Genetic variances for dry matter yield, nitrogen content, and nitrogen yield in crested wheatgrass-alfalfa mixtures by K.H. Asay and H.F. Mayland
Range condition assessment and the concept of thresholds: A viewpoint by M.H. Friedel
Stable states and thresholds of range condition on North American range- lands: A viewpoint by W.A. Laycock
Woody and herbaceous aboveground production of a Patagonian steppe by Roberto J. Fernindez-A., Osvaldo E. Sala, and Rodolfo A. Golluscio Importance of hypocotyl hairs in germination ofArtemisia seeds by James A. Young and Ellen Martens
Interference between yellow starthistle and pubescent wheatgrass during grass establishment by T.S. Prather and R.H. Callihan
Animal Ecology
447 Risk of predation and food consumption by black-tailed jackrabbits by William S. Longland
Plant-Animal Interaction
451 Forage yield and white-tailed deer diets following live oak control by Timothy E. Fulbright and Andres Garza, Jr.
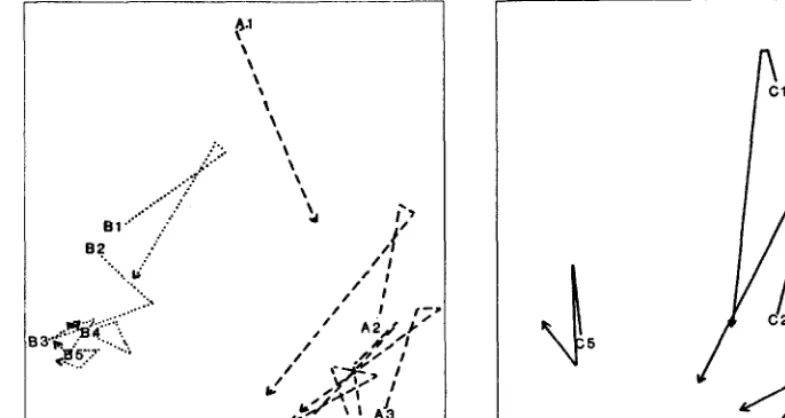
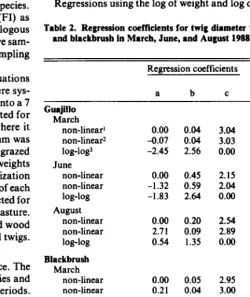
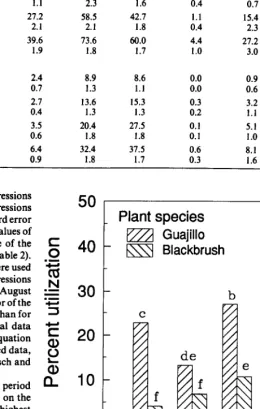
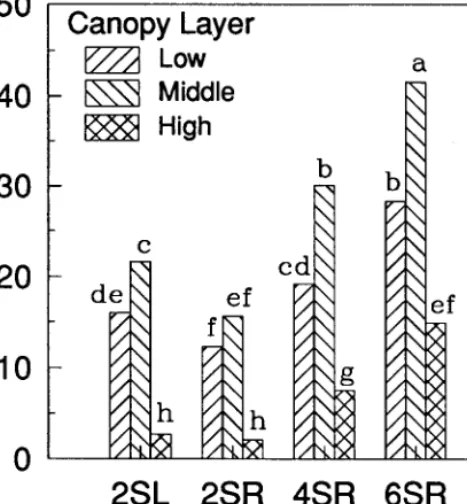
456 Utilization patterns by Angora goats within the plant canopies of two Acuciu shrubs by M. Keith Owens
462 Preference of wintering sage grouse for big sagebrush by Bruce L. Welch, Fred J. Wagstaff, and Jay A. Roberson
466 Grazing effects and range trend assessment on California bighorn sheep range by Brian M. Wikeem and Michael D. Pitt
Grazing Management 471
475 483
487
Soils 491
Effects of seasonal rest in aboveground biomass for a native grassland of the flood Pampa, Argentina by L.G. Hidalgo and M.A. Cauhepe
Overgrazing: Present or absent? by Allan D. Wilson and Neil D. MacLeod Forest Service and livestock permittee behavior in relation to wilderness designation by Mitchel P. McClaran
Desert mule deer use of grazed and ungrazed habitats by Kim E. Ragotz- kie and James A. Bailey
Published bimonthly-January, March, May, July, September, November Copyright 1991 by the Society for Range Management
INDIVIDUALSUBSCRIPTION is by membership in the Society for Range Management.
LIBRARY or other INSTITUTIONAL SUBSCRIP- TIONS on a calendar year basis are $56.00 for the United States postpaid and $66.00 for other coun- tries, postpaid. Payment from outside the United States should be remitted in US dollars by interna- tional money order or draft on a New York bank. BUSlNESSCORRESPONDENCE,concerningsub- scriptions, advertising, reprints, back issues, and related matters, should be addressed to the Manag- ing Editor, 1839 York Street, Denver, Colorado 80206.
EDITORIALCORRESPONDENCE, concerning manu- scripts orothereditorial matters, should beaddressed to the Editor, Gary Frasier. 1300 Wheatridge Ct., Loveland, Colorado 80537. Page proofs should be returned to the Production Editor, 1839 York Street, Denver, Colorado 80206.
INSTRUCTIONS FOR AUTHORS appear on the inside back cover of most issues. A Style Manual is also available from the Society for Range Manage- mentattheaboveaddress@$2,00forsinglecopies; $1.25 each for 2 or more.
THE JOURNAL OF RANGE MANAGEMENT (ISSN 0022-409X) is published six times yearly for $56.00 per year by the Society for Range Management, 1839 York Street, Denver, Colorado 80206. SECOND CLASS POSTAGE paid at Denver, Colorado. POSTMASTER: Return entire journal with address change-RETURN POSTAGE GUARANTEED-to Society for Range Management, 1839 York Street, Denver, Colorado 80206.
Managing Editor PETER V. JACKSON Ill
1639 York Street Denver, Colorado 80206 Editor
GARY FRASIER 1300 Wheatridge Ct. Loveland, Colorado 80537
Nutrition
497
Mountain mahogany and cottonseed meal as supplements for grass hay
by
G. Nunez-Hernandez,
Joe D. Wallace, Jerry L. Holechek, Michael L.
Galyean, Doug W. King, and Robert M. Kattnig
Improvements
501
Mefluidide effect on weeping lovegrass heading, forage, yield, and quality
by Larry M. White
508
Control of woody plants in grazing lands on the Pacific Coast of Mexico
by M. Garcia-Holquin,
R.W. Bovey, and J.L. Schuster
Sampling
512
Chemical composition of forage and feces as affected by microwave oven
drying
by J.F. Karn
515 Diet sample
collection by esophageal fistula and rumen evacuation tech-
niques
by Kenneth C. Olson
TECHNICAL
NOTES
520 Mineral content
of guajillo regrowth following roller chopping
by
Timothy E. Fulbright, J. Paul Reynolds, Samuel L. Beasom, and Stephen
Demarais
BOOK REVIEWS
524
Fabalesby Rupert C. Barneby;
Grazing Research: Design, Methodology, and Analysisedited by G.C. Martin;
Common Legumes of the Great Plainsby James Stubbendieck and Elverne C. Conard;
Secondary Succes- sion and the Evaluation of Rangeland Conditionedited by W.K. Lauen-
roth and W.A. Laycock;
Maintaining the Long-term Productivity fo Pacific Northwest Forest Ecosystemsedited by D.A. Perry et al.
MISCELLANEOUS
442
SRM Nominations by Petition
523 Application for SRM Executive Vice-President position
ASSOCIATE EDITORS DONALD BEDUNAH
School of Forestry University of Montana Missoula, Montana 59812 DAVID ENGLE
Agronomy Department Oklahoma State University Production Edltor Stillwater, Oklahoma 740;8 PATRICIA G. SMITH TIMOTHY E. FULBRIGHT
Society for Range Management College of Agriculture 1839 York Street Texas A&l
Denver, Colorado 80206 POB 156, Sta. 1 (303) 355-7070 Kingsville, Texas 78363 Book Review Editor G. FRED GIFFORD DAVID L. SCARNECCHIA Range, Wildlife 8 Forestry
Department of Natural Resource Sciences 1000 Valley Road Washington State University University of Nevada Pullman, Washington 99164-6410 Reno, Nevada 89512
KRIS HAVSTAD USDA-ARS, Dept. 3JER Box 3003, NMSU
Las Cruces, New Mexico 88003 RODNEY HEITSCHMIDT
USDA ARS
Livestock 8 Range Research Rt 1 Box3
Miles City, Montana 59301
KIRK MCDANIEL
Dept. of Animal & Range Science Box 31
New Mexico State University Las Cruces, New Mexico 88003-0003 ROY STRANG
British Columbia Inst. of Tech. 3700 Willingdon Ave. Burnaby, British Columbia CANADA V5G 3H2 JERRY HOLECHEK
Animal 8. Range Science Dept. 3-l Box 30003 New Mexico State University Las Cruces, New Mexico 88003-0003
DAVID M. SWIFT
Natural Resources Ecology Colorado State University Ft. Collins, Colorado 80523 HERMAN S. MAYEUX, JR. PAUL TUELLER
Genetic variances for dry matter yield, nitrogen content,
and nitrogen yield in crested wheatgrass-alfalfa mixtures
K.H. ASAY AND H.F. MAYLANDAbstract
Since its introduction from Asia in the early 19OOs, crested wheatgrass [Agropyron cristatum (L.)Gaertner, A. desertorum (Fisch. ex Link) Schultes et al.] has had a major impact on the improvement of western rangelands of North America. Most of the early seedings with this cool-season grass were made as monocul- tures. Present and projected use of rangelands, however, prescribe that future crested wheatgrass cultivars have the genetic potential to be an effective component in a species complex including other grasses, shrubs, and forbs. The present study was conducted to evaluate the effect of associated alfalfa (Medicago sativu L.) on the performance and genetic variability in a 50-clone sample of a tetraploid crested wheatgrass breeding population. Significant (P<O.O5) differences were found among the clonal lines for dry matter (DM) yield, nitrogen (N), and N yield. Opportunities for genetic improvement, as indicated by the magnitude of the genetic variation for these characters, was significantly increased when the grasses were grown in association with alfalfa. Signitlcant (P<O.Ol) and positive correlations of clonal means between stand types indicated that differences among the clonal lines in DM yield, N content, and N yield were relatively consistent when grown with or without alfalfa. These results indicate that initial screening could be effectively done in tetraploid crested wheatgrass in the presence or absence of alfalfa. Final evaluation of breeding lines and exper- imental strains, however, should be done with alfalfa if the object is to develop cultivars to be grown in combination with that species. Key Words: Agropyron cristatum, Agropyron desertorum, Me& cugo sutivu, heritability, genotype by environment, grass breeding, nitrogen uptake, nitrogen transfer, nitrogen mineralization
The first successful introduction of crested wheatgrass [Agro- pyron cristatum (L.) Gaertner, A. desertorum (Fish. ex Link) Schultes, and A. fragile (Roth) Candargy] into North America was made in 1906 (Dillman 1946). It has since had more impact on revegetation of western rangelands in North America than any other grass. Most of the early crested wheatgrass seedings were made as monocultures; however, environmental concerns and multiple demands now made on rangelands dictate that other grass, shrub, and forb species be included in the seeding mix.
The beneficial effects of legumes on succeeding crops in a rota- tion is well documented (Lohnis 1926, Trenbath 1974, Heichel 1987). The positive contribution of nitrogen (N) to this interaction was established in the early 1900s (Fred et al. 1932, Heichel 1987) and will likely have increasing economic impact as the availability of fossil fuels becomes more critical. The major benefits stemming from grass-legume mixtures are related to N transfer from the legume to associated grasses. Nitrogen is released from roots of the legume plant through active or passive release of soluble materials from intact cells, sloughing and decomposition of epidermal cells, lysis of cells by pathogens, and senescence of nodules and roots
Authors are research geneticist and research soil scientist, USDA-ARS, Logan, Utah 84322-6300 and Kimberly, Idaho 83341-9710, respectively.
Cooperative investigations of USDA-ARS and the Utah Agricultural Experiment Station, Logan, Utah 84322. Approved as Journal Paper No. 4045. The authors acknowledge the technical assistance provided by W.T. Hansen, GE. Shewmaker, and S.B. Hansen.
Manuscript accepted 16 December 1990.
418
(Brophy and Heichell989). Nitrogenous compounds secreted into the surrounding soil medium consist of ammonia, glutamate, serine, alanine, and aspartate (Ta et al. 1986).
Brophy and Heichel(1989) studied net N release from actively growing root systems of alfalfa (Medicago sativa L.) and soybeans (Glycine max L. Merr.) in an enclosed root chamber containing sterile sand medium. Alfalfa plants inoculated with an adapted rhizobial strain and irrigated with N-free nutrient solution released 4.5% of symbiotically fixed plant N into the root zone from 85 to 173 days after germination. In these studies, release of N from alfalfa roots was increased by shoot harvest and periodic water stress. In greenhouse trials, Ta and Faris (1987) found that yields of herbage and protein were significantly increased in 5 perennial forage grasses when grown in association with alfalfa, red clover ( Trifolium pratense L.), and birdsfoot trefoil (Lotus corniculatus L.) compared to grasses in monoculture.
In Switzerland, Boller and Nosberger (1987) studied symbiotic production of N in white clover (Trifolium repens L.) and red clover when grown in association with perennial ryegrass (L.olium perenne L.) and Italian ryegrass (Lolium multzjlorum Lam.), respectively. Annual yield of clover N derived from symbiosis averaged I31 kg ha-’ and 308 kg ha-i during the seeding year and first production year, respectively. The apparent clover-to-grass transfer to fixed N contributed up to 52 kg N ha-’ to the N yield of the mixtures. Symbiotic N yield levels were decreased an estimated 25 to 35% by fertilization with supplemental N.
Brophy et al. (1987) reported that significant amounts of N were transferred from alfalfa and birdsfoot trefoil to reed canarygrass (Phalaris arundinacea L.) under field conditions. Although the amount of N transferred was dependent on interspecies distance and legume/grass ratio, 17% of the N fixed by alfalfa plants and 13% of that fixed by birdsfoot trefoil was transferred to the grass when grown within I m.
Leyshom et al. (1981) studied mixtures of grass and alfalfa in semiarid environments of Saskatchewan, Canada. Alfalfa plus crested wheatgrass and alfalfa plus Russian wildrye [ Psathyrosta- chys juncea (Fisch.) Nevski] produced significantly more forage than either crested wheatgrass or Russian wildrye alone. The advantages of alfalfa in crested wheatgrass stands also were dem- onstrated by Rogler and Lorenz (1969). Alfalfa-crested wheatgrass mixtures were more productive than pure stands of crested wheat- grass, due not only to the productivity of the additional alfalfa but the increased vigor of the grass component as well.
Rumbaugh et al. (1982) found that fourwing saltbush [Atriplex canescens (Pursh) Nutt.], cicer milkvetch (Astragalus cicer L.), sicklepod milkvetch (A. falcatus Lam.), and alfalfa significantly increased the forage yield, protein concentration, and protein yield of crested wheatgrass on a semiarid range site in central Utah. In grazing trials conducted during the winter by Gade and Provenza (1986), sheep maintained higher intake and protein levels on crested wheatgrass-shrub mixtures than on pure grass stands.
Rumbaugh and Pendery (1986) evaluated estimated gains from selection in a lOO-clone alfalfa breeding population when grown alone and in association with 5 perennial grasses. They suggested that grasses and the legumes to be grown in association with them be bred concurrently and that germplasm should be screened in
mixed stands. the forage yield and N data and expressed as g N plant“. Casler and Drolsom (1984) evaluated differences in forage yield
among cultivars of 4 cool-season perennial grass species in pure stands and in mixtures with alfalfa. No consistent differences were found in the magnitude of heritability values between the 2 stand types and rank correlations between stand types were generally nonsignificant. They suggested that cultivars of species evaluated in their trials be evaluated in mixed stands if results are to be applied to mixed stands. Reich and Casler (1985) found that alfalfa competition tended to accentuate differences among smooth bromegrass (Bromus inermis Leyss.) genotypes for neutral deter- gent fiber (NDF), acid detergent fiber (ADF), and cellulose. Fam- ily by stand type interactions were nonsignificant for ADF and cellulose at most growth stages, and for NDF at the early growth stage.
Data were subjected to standard analyses of variance and regres- sion procedures. Broad-sense heritability values were computed on a mean basis as the ratio c$ / c&,, where &! is the variance compo- nent arising from differences among clonal lines, and &I, is the phenotypic variance among the clonal lines or the variance of a clonal mean. In the computation of the variance components and F tests, clonal lines, replications, and years were considered as ran- dom variables and stand types as fixed. Approximate F tests were computed when determined necessary by evaluation of expected mean squares.
Results and Discussion
Complex symbiotic and competitive relationships among diverse species in the range ecosystem must be understood if grass breeders are to make realistic genetic progress. Unfortunately, little is known regarding these relationships in semiarid environments and their potential impact on genetic responses of individual species in the plant community. In the present study, forage yield, N concen- tration, and N yield were determined for 50 clonal lines of crested wheatgrass when grown in monoculture and in association with alfalfa during 3 production years. Objectives were to: (1) assess the effect of stand type on genetic variances and heritability values, and (2) evaluate the genotype by stand type interactions.
Significant (PCO.01) variation was found among the 50 crested wheatgrass clonal lines in forage yield, N content, and N yield when grown with and without alfalfa during each of the 3 harvest years, and in the analyses combined over years (Tables 1,2, and 3). The significance levels of the mean squares from the analyses of var- iance, along with the magnitude of the range in means, genetic variance, and broad-sense heritability values indicate that excel- lent opportunities exist to improve the genetic potential of this Hycrest breeding population to produce more and higher quality (based on crude protein) forage.
Materials and Methods
Table 1. Mean squares from analyses of variance for forage yield, nitrogen concentration, and nitrogen yield data from 50 clonal lines of crested wheatgrass grown alone and with alfalfa.
Plant materials consisted of 50 clonal lines randomly selected from a breeding population of the crested wheatgrass cultivar ‘Hycrest’. This cultivar was originally derived from a hybrid between an induced tetraploid form of A. cristatum and natural tetraploid A. desertorum (Asay et al. 1986). The experiment was conducted at the Utah State University Evans Experimental Farm, 2 km south of Logan, Utah (41 45’ N, 11148’ W, at an altitude of 1,350 m above sea level). Vegetative propagules, initially started in pots in the greenhouse, were transplanted in the field in mid-April 1986 as single-plant plots on 0.5 m centers. Plots were arranged in a split-plot design with 4 replicates. Whle plots were randomly designated as grass-alone or grass-alfalfa and the 50 clonal lines were randomly assigned to subplots in each whole plot. Grass- alone plots were surrounded (0.5 m) on 4 sides by propagules of another crested wheatgrass plant. Grass plants established with alfalfa were surrounded (0.5 m) on each side by an alfalfa plant derived from the NC-83-l germplasm pool.
Source of
variation 1987 1988 1989 1987-89
Stand type (S) Clone (C) sxc Year (Y) SXY CXY SXCXY
,_ ________ DMYield(gplant-‘)---
19899 69567** 126558** 192822.
5736”. 1866** 5269** 10964+*
1321 305** 807** 1408
558905” 11601* 954** 513**
_.
Stand type (S) Clone (C) sxc Year(Y) SXY CXY SXCXY .-_____________N(mg~~1)_-_---244;--
5380.2** 525.8’. 849.7**
52.9** 5.2** 6.6** 32.6** .
7.6 1.5** 2.6 4.5
6309.1** 755.8** 16.1** 3.6
Soil at the study site is a Nibley silty clay loam and is classified as a fine, mixed mesic aquic Argiustoll. The area was fertilized with 45 kg N per ha in October 1985 and received no additional fertilizer or supplemental irrigation during the period of the study. Following a year of establishment, all plots were harvested at a 7-cm stubble height twice in 1987 (2 June and 15 September) and once in each of the 2 subsequent years (9 June 1988 and 12 June 1989). With the exception of the second harvest in 1987, grasses were harvested at or near the bloom stage of development. An additional harvest was made each year from the alfalfa plots on 27 August 1987, 18 July 1988, and 31 July 1989.
Stand type (S) Clone (C) sxc Year(Y) SXY
--_--___-__NYield(gNP~nt~‘)---
136.697. 20.207** 41.620’. 170.832.
CXY SXCXY
1.832;’ 0.288** 0.769** 2.090**
0.516 0.056* 0.190** 0.388
338.939** 13.846*
0.399** 0.187*
l ~**Significant at 0.05 and 0.01 level of probability level, respectively.
Herbage was dried at 60” C for 48 hours in a forced draft and ground by Wiley mill to pass through I-mm screen. Dry weights were determined and expressed as g plant“. Total nitrogen (N) was determined by the Kjeldahl procedure 31-3 (Bremner and Mulva- ney 1982) modified by excluding the selenium catalyst in the diges- tion salt. Analytical precision and accuracy were typified by 98% (C.V. = 2%) recovery of total N in the National Bureau of Stand- ards citrus leaf sample, NBS-1572, which was analzyed with the grass samples. Nitrogen yield was determined for each plot from
Crested wheatgrass clones grown with alfalfa produced signifi- cantly more dry matter than when grown alone during each of the 3 years and when the data were combined over years. The effect of alfalfa on DM yield of associated grass plants was particularly evident in 1988 and 1989. The clones produced 133,66,94 g plant-’ in 1987, 1988, and 1989, respectively, with alfalfa compared to 119, 39, and 59 g plant-’ when grown when without alfalfa. The smaller effect of associated alfalfa in 1987 was possibly due to the residual N remaining in the soil from the application made during the fall of 1985. Also, substantially more precipitation was received in 1987 than in 1988 and 1989.
Table 2. Summery of DM yield data from 50 cionai lines of crested wbeetgrass grown alone and in essociation with eifaifa.
Parameter 1987 1988
Grass alone
1989 1987-89 1987 1988
Grass-alfalfa
1989 1987-89 Min.1
Max. Mean2 E Hb Mean
60.6 182.2 118.7~ 692.9**
0.80
12.3 67.8 39.3c 143.1**
0.83
10.0 30.1 95.0 111.7 58.7~ 72.2 385.9” 304.7**
0.84 0.76
_______ _ _____- -G~ssComponent________________
52.9 23.7 26.3 34.7
196.6 99.8 157.5 142.3
132.8b 65.5b 94.2b 97.6 495.0** 305.1+* 932.3”. 489.2**
0.55 0.82 0.88 0.78
________ ---AlfalfaComponent ______ _________
274.4a 90.8a 204.3a 189.8
‘Data are expressed in g/plant; d = Variance Component among clonal lines! Hb q Heritability computed in the broad sense on a mean basis.
2Means for grass alone, grass component with alfalfa, and alfalfa component with grass are not significantly different within years at the 0.05 probability level when followed by the same letter. Approxunate F test was significant (P<O.OS) in analysis combined over years (1987-89).
***Means squares from which variance components were computed are significant at 0.05 and 0.01 probability levels, respectively.
The benefits of associated alfalfa were also reflected in concen- tration and yield of N. The clonal lines had a higher N concentra- tion and N content when grown with alfalfa than without alfalfa all 3 years (Table 3). In contrast to the DM yield data, the effect was at least as great in 1987 as in the 2 subsequent years. Forage of the Hycrest clones had 15.5, 11.4, and 10.5 mg N g-’ without alfalfa in 1987, 1988, and 1989, respectively, compared to 22.8, 13.7, and 13.4 mg N g“ plots with alfalfa. Similar trends were observed in nitrogen yield of the clonal lines. In the analysis combined over the 3 years (Table l), the approximate F test lacked the precision to detect statistical significant differences in N concentration due to stand type.
Opportunities for selection, as indicated by the genetic variances among clones and the range in clonal means, were also enhanced by the associated alfalfa plants. As was the case with mean DM yield, the positive effects of alfalfa on the genetic variance and range for DM yield was much more pronounced in 1988 and 1989 than during the first year of data collection, 1987 (Table 2). In 1988 and 1989, the genetic variance (& among clones grown with alfalfa was more than twice as large as the variance among the same clones grown without alfalfa (305 vs 143 and 932 vs 386). The range in clonal means was also substantially greater in the grass-alfalfa plots. Forage yield of the crested clonal lines ranged from 12 to 68 and from 10 to 95 g/plant in 1988 and 1989 for grass alone, compared with a range of 24 to 100 and 26 to 158 g/plant for the grasses with alfalfa. Broad-sense heritability values for DM yield were similar for the 2 stand types and ranged from 0.55 to 0.88.
A positive effect of associated alfalfa on the range and genetic
variance for N concentration and N yield was evident during the 3 years of the study and in the combined analyses. The effect was particularly noteworthy in the analyses of the N yield data (Table 3). The genetic variances for N yield were 0.11,0.02, and 0.04, and 0.04 in each of the 3 years and in the combined analysis, respec- tively. Comparable values for grass with alfalfa were 0.25, 0.05, 0.16, and 0.10. As was the case with DM yield, broad-sense herita- bility values were not substantially affected by the associated legume, ranging from 0.67 to 0.8 1 without alfalfa and from 0.56 to 0.83 in the grass with alfalfa plots. Similar trends were detected in the N concentration data.
The clone by stand type interaction was significant (KO.01) in the 1988 and 1989 analyses of the DM yield data, but was nonsig- nificant in 1987 and in the combined analysis (Table 1). This suggests that the relative productivity of individual clonal lines was somewhat inconsistent when grown alone and with alfalfa. How- ever, annual forage yields of the clones in the 2 stand types were positively and significantly (P<O.O1,48 df) correlated in all 3 years and in the combined analysis of the data. The correlation coeffi- cients (r) between stand types was 0.63,0.77,0.80, and 0.79 in 1987,
1988, 1989, and in the combined analysis, respectively.
A similar degree of consistency was found for N concentration and N yield. Although the clone by stand type interaction was significant in 3 of the 8 instances, correlation analyses indicated that differences among the clonal lines were relatively consistent for the 2 stand types. Correlation coefficients between stand types for the data combined over years were 0.78 for N concentration and 0.79 for N yield.
Table 3. Summary of nitrogen concentration and nitrogen yield data from 50 cionai lines of crested wbeatgrrss grown alone and in eesocietion with aifaifi
Grass alone Grass-alfalfa
Parameter 1987 1988 1989 1987-89 1987 1988 1989 1987-89
~~~~~~_~~~~_______~~____________________N(~gg~’)--~~~---~~~--~~~~~~~~~~~~~~~
Min. il.8 9.6 9.1 10.4 18.6 11.9 10.8 14.4
Max. 23.6 13.2 12.6 15.8 31.9 15.8 17.1 20.4
Mean’ 15.5 11.4 10.5 12.5 22.8 13.7 13.4 16.6
az2 4.6** 0.5’. 0.3** 0.6+* 7.9** 0.6** i .o+* 0.9**
Hb 0.86 0.74 0.50 0.53 0.80 0.66 0.58 0.44
---~~---~________.________________NYieid(gNP~ant~‘)---~---~~~~~~~~
Min. 0.862 0.135 0.115 0.420 1.268 0.283 0.316 0.623
Max. 2.562 0.695 1.066 1.361 4.390 1.359 2.104 2.389
Mean 1.795 0.444 0.613 0.951 2.964 0.893 1.258 1.705
XL! 0.109** 0.016** 0.040** 0.03551 0.247** 0.053** o.i59** 0.100*+
Hb 0.74 0.80 0.81 0.67 0.56 0.81 0.83 0.65
‘Means for grass grown alone or with alfalfa within years and when combined over years are significantly different at the 0.05 probability level, except for N concentration fo$bin+ over y&F
- Vanance Corn onent among clonal lines; Hb = Heritability computed in the broad sense on a mean basis.
***Means squares rom which variance components were computed are significant at 0.05 and 0.01 probability levels, respectively.
Although the objective of this study was not to compare the yields of the grass and alfalfa plants in the mixture, it is noteworthy that alfalfa plants produced significantly more total forage than their grass counterparts in all 3 years (Table 2). This superiority resulted entirely from the aftermath yields of the alfalfa component in 1987 and 1988. Crested wheatgrass produced significantly (P<O.OS) more forage than the bordering alfalfa plants at the first harvest in 1987 (114 vs 97 g plant-‘) and the first-harvest yields of the 2 species were equivalent in 1988. In 1989, the alfalfa compo- nent (148 g plant-‘) was significantly (P<O.Ol) more than that produced by the associated grass clones (94 g plant-‘).
It is evident that sufficient genetic variability remains in this breeding population of crested wheatgrass to effectively select for improved forage yield and nitrogen content. Genetic variability for these attributes was significantly increased by the association of alfalfa plants in the mixture. Higher N concentration in grasses grown with alfalfa indicates that the increased genetic variability resulted primarily from a more optimum environment provided by the apparent transfer of biologically fixed N from the associated alfalfa plants to the grass root systems. These results support earlier findings for perennial range grasses (Rumbaugh et al. 1984) and perennial humid grasses (Reich and Casler 1985). The rela- tively consistent performance of the grass clones across stand types is encouraging and may have been associated with autotetraploid nature of the Hycrest breeding population. Additional research would be in order to evaluate the stability of diploid populations of crested wheatgrass. Our findings indicate that initial screening for DM yield, nitrogen concentration, and nitrogen yield could be effectively done in tetraploid crested wheatgrass in plots without alfalfa. Final evaluation of breeding lines and experimental strains, however, should be done with alfalfa if the object is to develop cultivars to be grown in combination with that species.
References
Asay, K.H., D.R. Dewey, F.B. Gomm, W.H. Horton, and K.B. Jensen. 1986. Genetic progress through hybridization of induced and natural tetraploids in crested wheatgrass. J. Range Manage. 39:261-263. Boiler, B.C., and J. Nosberger. 1987. Symbiotically fixed nitrogen from
field-grown white and red clover mixed with ryegrasses at low levels of r>N-fertilization. Plant and Soil 104219-226.
Bremner, J.M., and C.S. Mulvaney. 1982. Nitrogen-total. p. 595-624. In: A.L. Page (ed.). Aaron. No. 9 Part 2. Amer. Sot. Agron. Publ. Madison, Wisconsin.
Brophy, L.S., and G.H. Heichel. 1989. Nitrogen release from roots of alfalfa and soybean grown in sand culture. Plant and Soil 116:77-84. Brophy, L.S., G.H. Heichel, and M.P. Russelle. 1987. Nitrogen transfer
from forage legumes to grass in a systematic planting design. Crop Sci. 27:753-758.
Casler, M.D., and P.N. Drolsom. 1984. Yield testing cool-season forage grasses in pure stands vs. binary mixtures with alfalfa. Crop Sci. 24:453-456.
Dillmnn, AC. 1946. The beginnings of crested wheatgrass in North Amer- ica. J. Amer. Sot. Agron. 38:237-250.
Fred, E.B., I.L. Baldwin, and E. McCoy< 1932. Root nodule bacteria and leguminous plants. Studies in Science 5: Univ. of Wisconsin Press, Madison.
Gade, A.E., and F.D. Provensa. 1986. Nutrition of sheep grazing crested wheatgrass versus crested wheatgrass-shrub pastures during winter. J. Range Manage. 39:527-530.
Heichel, C.H. 1987. Legume nitrogen: symbiotic fixation and recovery by subsequent crops. p. 63-80. In: Helsel, Z.R. (ed.) Energy in plant nutri- tion and pest control. Elsevier Science Publ. B.V., Amsterdam, The Netherlands.
Leyshor, A.J., M.R. Kilcher, and J.D. McElgunn. 1981. Seeding rates and row spacings for three forage crops grown alone or in alternate grass- alfalfa rows in southwestern Saskatchewan. Can. J. Plant Sci. 61:7 1 l-7 17. Lohnis, F. 1926. Effect of growing legumes upon succeeding crops. Soil Sci.
22:355-389.
Reich, J.M., and M.D. Casler. 1985. Effect of maturity and alfalfa competi- tion on expected selection response for smooth bromegrass forage qual- ity traits. Crop Sci. 25:635-640.
Rogler, G.A., and R.L. Lorenz. 1969. Pasture productivity of crested wheatgrass as influenced by nitrogen fertilization and alfalfa. USDA- ARS Tech. Bull. 1402 p. l-33.
Rumbaugh, M.D., D.A. Johnson, and G.A. Van Epps. 1982. Forage yield and quality in a Great Basin shrub, grass, and legume pasture experi- ment. J. Range Manage. 35604609.
Rumbaugh, M.D., and B.M. Pendery. 1986. Interspecific relations and the breeding of pasture plants for semiarid regions. Agron. Sot. New Zea- land. p. 285-290. In: T.A. Williams and G.S. Wratt (ed.) Plant breeding symposium DSIR 1986. Spec. Pub. 5. Aaron. Sot. New Zealand. Ta, T.C., and M.A. Faris. 1987. Species variation in the fixation and
transfer of nitrogen from legumes to associated grasses. Plant and Soil 98~265-274.
Ta, T.C., F.D.H. MacDowell, and M.A. Faris. 1986. Excretion of nitrogen assimilated from Ns fixed by nodulated roots of alfalfa (Medicago sut- ha). Can. J. Bot. 64:2063-2067.
Trenbath, B.R. 1974. Biomass productivity of mixtures. Adv. Agron. 26177-210.
Range condition assessment and the concept of thresholds:
A viewpoint
M.H. FRIEDEL
Abstract
Dissatisfaction persists with current approaches to range condi- tion and trend assessment. Sometimes assessed condition does not truly represent the past or the potential of range. One of the likely causes is a failure to re-examine and change if necessary the theo- retical basis of assessment, in line with developing understanding of ecological processes. The concept of thresholds of environmen- tal change appears to provide a reasonable alternative in some circumstances to the concepts of gradual retrogression and secon- dary succession which are currently accepted. I suggest that envi- ronmental change can be discontinuous, with thresholds between alternative states. Once a threshold is crossed to a more degraded state, the former state cannot be attained without significant man- agement effort, such as prescribed burning, ploughing, or herbi- cide application, rather than simple grazing control. Examination of data from extensive monitoring programs and from a study of grazing impact, as well as more general sources of information, indicates that thresholds of change may be identifiable in arid rangelands. A practical means of monitoring proximity to thresh- olds is available and, with the aid of multivariate analysis, the effects of spatial variability and season can be separated from those of management. The potential of this approach deserves investiga- tion in a wider variety of environments.
Key Words: succession, site potential, ordination, classification, trend
The assessment of range condition and trend remains a source of controversy, despite years of practical experience and discussion (Lauenroth and Laycock 1989). What attributes are measured and how they are measured, and how the measurements are inter- preted, are the subject of continuing debate. More importantly, perhaps, the advances in our understanding of ecological processes have not been matched by developments in the theory and practice of range assessment. The concept of thresholds of environmental change between domains of relative stability is well recognized in ecological literature (Holling 1973, Wissell984). It is not at present a part of range assessment philosophy and may offer a new approach to range monitoring.
Anomalies in the condition assessments of central Australia’s arid rangelands stimulated an examination of existing data sets (e.g., Foran et al. 1986) and alternative methods of data collection (e.g., Friedel et al. 1988b). At the same time, the “quantitative climax” approach of Dyksterhuis (1949) as commonly applied demanded challenge. In this paper, I present evidence from a variety of sources that the concept of thresholds offers a useful framework for identifying important environmental changes. I also consider whether practical ways of recognizing thresholds can be devised for range assessment.
Concepts of Environmental Change
The quantitative climax approach to range management and assessment has focused attention on successional processes (Lauen- roth and Laycock 1989). Excessive grazing is perceived to lead to
The author is principal research scientist, CSIRO Division of Wildlife and Ecology, Alice Springs, Northern Territory 0871, Australia.
Manuscript accepted 19 October 1990.
422
retrogression or decline in range condition (Stoddart et al. 1975) and it is assumed that reduction or removal of grazing pressure allows successional processes to restore the range to what it was, essentially by reversing the path taken by retrogression.
In environments driven by unpredictable events, e.g., highly variable rainfall throughout any year or a variable starting time for rainy seasons, succession and retrogression do not coincide, and there is commonly a hysteresis effect where decline and recovery follow different paths (Noy-Meir and Walker 1986), even if the system returns to its original starting point.
In practice, a starting point cannot be determined because it depends on current seasonal conditions. An alternative model is of range in a series of short-lived states under different combinations of seasons, grazing, fire, and local variability. Most of these states do not preclude a shift to other short-lived states. However, certain combinations may push the system into a new state that is not readily reversed; e.g., an eroded soil surface can develop following heavy grazing coupled with torrential rain, in which case the sys- tem has crossed a threshold.
A threshold has these characteristics: first, it is the boundary in space and time between 2 states, e.g., grassland and shrub-invaded grassland; and second, the initial shift across the boundary is not reversible on a practical time scale without substantial intervention by the range manager, e.g., with herbicides, heavy machinery, or fire. The concept is compatible with the state-and-transition model of Westoby et al. (1989), focusing on those transitions which, for management purposes, are l-way. Stocking-rate reductions are not enough to cause a reversion to the former state.
Current Approaches to Range Assessment
Two different philosophies have influenced assessment up to the present: the ecological approach based on comparisons with the climax state (e.g., Dyksterhuis 1949); and the site potential approach based on productive potential for a particular use (e.g., Humphrey 1949). Both focus on the forage layer. Wilson (1986) has outlined an alternative based on multivariate site potential, which combines important elements of the 2 philosophies and includes soil assessment. This approach has yet to significantly affect entrenched range monitoring systems, although it represents a philosophical advance.
The ecological or climax approach to range condition assess- ment has,been criticized for some time (Smith 1978, 1989; Wilson 1989), but it is still in active use in a wide variety of rangelands (Tainton 1986, Pendleton 1989). The weaknesses of the approach have been summarized by Smith (1978) as: (1) climax is not always the most desirable condition; (2) pristine conditions may not be the actual climax for a site; (3) it does not allow for exotic species; and (4) it is not well suited to woodland and forested rangeland.
If one takes the approach of assessing site potential for particu- lar uses, a single vegetation state can have a variety of potentials depending on use, Site stability or degradation cannot be inferred from the condition rating because the rating will vary with poten- tial use. Whatever approach is taken to assessment, irreversible changes in the soil will be important indicators of site deteriora- tion. As a consequence, range monitoring must incorporate 3 tiers of assessment-the herbaceous layer and soil, and the tree-shrub layer, where it is present.
2.40
1.60 Fusion Level -
0.80
0.00
-1
A
-3:-
,_ _---
________
l-3
_
r-l
Ltal information. This approach does not assume a climax, but produces classes and orders of locations which can be interpreted according to known site factors, seasonal conditions, and man- agement history.
1
J_Foran et al. (1986) classified forage composition from 65 moni- toring locations within an arid shrubby grassland range site. They concluded that, apart from differences attributable to soil “type”(= variable depth or minor textural differences, G.N. Bastin pers. comm.) or season of assessment, there were just 2 classes associated with management effects (Fig. 1). These classes were identified as good and poor condition states, also distinguished by ordination on the basis of forage quality and quantity and soil stability. The same divisions into 2 classes of condition were apparent in moni- toring data from 4 other range sites, each analysed independently. The gradual deterioration implied by percentage deviation from a climax state, which the ecological approach creates, was not in evidence. Instead, a shift from 1 state or “domain”to another (Fig. 2, from Bradbury et al. 1983/4) appeared to have occurred.
v
--T-I
Poor condition”
Winter and Mixed growth response Minor soil variability Poor condition
Summer growth response Minor soil variability Good condition
Summer growth response
Fig. 1. Classification of 65 monitoring locations in an arid shrubby grass- land range site, on the basis of forage composition (from Foran et al. 1986).
In the application of the ecological or climax approach, as outlined by Dyksterhuis (1949) and Tainton et al. (1980), the seasonal fluctuations in forage composition are allowed for by comparing the monitored vegetation to an ecological benchmark which represents the vegetation climax and which is presumed to have experienced the same seasonal variations. There are many weaknesses in such a system. Benchmarks may be in atypically favoured locations, they may experience different rainfall or have a different fire history, or they may simply not exist and must be theoretically constructed. Differences due to variability within a site may be inappropriately attributed to management. Multiple benchmarks have been proposed (Wilson 1984, Bosch et al. 1987) to represent the variability within a range site or land unit.
Fig. 2. An ecosystem experiencing mild impacts, e.g., light graxing and changing seasons, shifts about within its original domain. A severe impact, e.g., extended heavy grazing, shifts the ecosystem across a thresh- old and into an entirely new domain (from Bradbury et al. 1983/4). A similar shift is evident in an ordination of the forage composi-
Changing States: Evidence of Thresholds
The development of multivariate analysis and increasingly easy access to it suggest the next step in the assessment of condition: the comparison of each monitored location with all others, using classification and ordination of forage composition. Classification assigns locations to classes on the basis of composition, not neces- sarily in any hierarchical order. Ordination arranges them in a lowdimensional space so that similar locations are near each other and dissimilar locations are far apart (Gauch 1982). Ordination axes identify major elements in the distribution patterns of the locations, which can be compared with independent environmen-
tion on a series of sample plots in an arid shrubby grassland range site (Friedel unpubl.). The plots were distributed along a gradient of grazing, centered on 3 independent watering points. Waterpoint A (Fig. 3a) had a history of heavy grazing by cattle while B (also Fig. 3a) was only lightly grazed because the water supply was ephemeral. The history of C (Fig. 3b) is detailed below. Sample plots Al and Bl were within 100 m of their waterpoints, A5 and BS were 2.0 km and 3.7 km, respectively, out from their waterpoints and intermediate plots were distributed along the radius from the waterpoint to sample plot 5. Each plot was 10 X 10 m, divided into 100, l-m2 quadrats. Ten quadrats from within each plot were chosen at random on the first sampling occasion and were resampled on 3 subsequent 6-monthly visits, making 4 samples in all. Frequency data for all species were collected from the quadrats and analysed with the multivariate analysis package TAXON (Ross et al. 1983), using the Bray-Curtis measure of dissimilarity. In the ordination graph (Fig. 3a,b), similar samples are close together, so that species composition of B5 for instance did not I change much over the 4 sampling occasions, whereas Bl changed a lot, but remained more like B5 than like A5.
Close to the waterpoints, where grazing is heaviest, shifts in
B
1
i
\ \
!41 +
Fig. 3. (a) left, (b) right. Ordination of the forage composition on sample plots in an arid shrubby grassland range site, showing axes 1 (x-axis) and 2 (y-axis). Plots were distributed on a radius out from 3 independent watering points and were measured 4 times at Cmonthly intervals. Watering points A and B (3a) are shown separately from water point C (3b) for clarity but are from the same ordination. Lines connect the sequential samples for each plot; arrows indicate the final occasion. Note that not all samples are distinguishable when composition differences are small. Craxing around waterpoints A and C was generally heavy; around waterpoint B it was generally light. The plots furthest from water were AS 2.Okm, B5 3.7km and CS 5.7km. See text for further details.
species composition over time can be large (Fig. 3a), and tend to become less at increasing distance from the waterpoints. A and B plots occupy different areas of the ordination space although they approach one another in their most degraded state (Al, Bl). In other words, A and B appear to be in 2 different domains of system behaviour.
If we now consider Fig. 3b, extracted from the same ordination as Fig. 3a, the change from 1 domain to another is indicated more clearly. Waterpoint C had a history of heavy grazing but CS was 5.7 km out from it, beyond the distance commonly walked by cattle. The closer 4 plots were in the same domain as those centered on A, while C5 was in the same domain as B. Although the range site is regarded as the same throughout, there is a change in state, in this case possibly a change in the soil, as a consequence of greater grazing pressure. Almost certainly a threshold has been crossed, but I have yet to determine finally that the change cannot be reversed.
Examples of Thresholds
Two thresholds are readily recognized in arid and semiarid rangelands: one separates grassland from woodland, and the other, stable from degraded soil. The change from grass to woody vegeta- tion can arise when the grass layer is grazed beyond its capacity to recover quickly, and fire suppression enhances the survival of woody plant seedlings. Grasses which have been grazed too heavily cannot grow fast enough to compete effectively for moisture in the topsoil. Germinating woody plants, which are not grazed, can out-compete the grasses and subsequently gain access to subsoil water as their root systems develop.
This kind of change has occurred in western New South Wales, where open eucalypt woodlands have become infested with inedi- ble shrubs, to the detriment of grazing sheep (Harrington et al. 1984). In southern New Mexico, drought rather than grazing has enabled honey mesquite (Prosopis glandulosa Torr.) to occupy black grama (Bouleloua eriopodu (Torr.) Torr.) grassland, and to modify the site to the extent that black grama is unable to establish (Hennessy et al. 1983).
Alternatively, periods of high rainfall can produce more topsoil
424
moisture than grasses can use, even when there is no grazing, and so woody seedlings once again can reach and tap the subsoil water supply. After a series of high rainfall years in central Australia, I found no evidence that grazing by cattle had affected the massive growth of shrubs in Acacia woodlands (Friedel 1985) and attrib- uted the cause to the absence of fire. Without fire, the conversion to shrubland was essentially permanent.
The second threshold is reached when soil erosion outstrips replenishment and soil physical and chemical properties are effec- tively altered irreversibly, for example when the sandy, nutrient-
rich A horizon is stripped from an impoverished medium or flne- textured B horizon. Rainfall is lost in runoff and the environment becomes too xeric for the ready establishment of either grasses or woody plants. In all cases, crossing a threshold means the vegeta- tion occupies a new domain and will not revert to its former state without considerable intervention.
Other thresholds include the change from palatable perennial species to palatable but shorter-lived species, e.g., from Atriplex spp. to grasses and Sclerolaena spp., which may not be significant in terms of animal production (Wilson 1989). Less commonly, there is a change from palatable short-lived grasses to unpalatable perennial grasses, e.g., from Enneupogon spp. to Aristidu strigosu (Henr.) S.T. Blake ex J.M. Black (B.D. Foran pers. comm.).
Noy-Meir and Walker (1986) have outlined models of grass/ woody plant and palatable/unpalatable grass interactions in Afri- can savannas. The Themeda triandra Forsk. grasslands of Natal change to Aristida junciformis Trin. & Rupr., B with overgrazing and this may represent a change of state which is not reversible within a realistic time frame for management. A wide variety of North American examples has been provided by Laycock (1991). Different range sites have differential susceptibilities to change. Some grasslands on heavy clay plains are unlikely to approach either threshold; species composition and productivity will simply shift about in response to grazing, fire, and season. Some savannas may develop into woodlands in the absence of deliberate interven- tion with fire, but may not be liable to soil erosion. Friedel(1987) found evidence for the existence of both types of threshold within a
I
2-I I I i
400 600 ,200 1600 2000
Tree equivalents (TEllha
Fig. 4. Proposed relationship between condition index (bezed on forzge
composition) and tree density in a zemierid zevznne rznge site, where tree equivalents are the number of treez standrrdieed to a height of 1.5 m. A-G are zempie points. Arrows indicate suggested areas for tbrezholds of chenge from (1) a graskdomineted domain to a tree-domineted domzin and (2) a tree-dominated dom& to en eroded domzin with reduced regeneration of vegetation (from Friedei 1987). The regression applies to points from D znd E to A.
South African savanna range site (Fig. 4). There, increased grazing was initially associated with greater tree density but, at an extremely overgrazed area, soil loss was considerable and tree establishment was suppressed. Both thresholds are found in some arid and semiarid Australian range sites and, almost certainly, the same is true elsewhere.
All the examples given so far refer to environmental changes away from a preferred state, that is, to range deterioration. Lay- cock (1989,199l) has observed that there are “suspended”stages of succession during the recovery process in arid range, too. Despite careful grazing management, range condition does not improve. Sometimes, for example, a rare combination of environmental conditions must occur before recovery proceeds; the necessary conditions are the means by which a threshold can be crossed.
The discontinuities in environmental change that are implied by the threshold concept can often be linked to the pulsed nature of rainfall, particularly in arid environments. Rainfall at different times of the year favours plants with different growth strategies; e.g., broad-leafed species may respond to winter rain, grasses to summer rain, while amount of rain determines whether perennial species or only ephemerals will establish (Westoby 1979/ 80). These shifts give rise to short-lived vegetation states. Changes from one long-lived state to another can also be driven by larger climatic events: several wet years may produce massive shrub establishment or enough fuel for major fires, or single large storms may initiate extensive erosion.
Detecting Thresholds
In view of the seasonal and spatial variability of vegetation in arid and semiarid rangelands, especially those rangelands that are currently dominated by short-lived species, range monitoring may be best served by a focus on the identification of thresholds. I investigated the possibility of recognizing thresholds at a single point in time, by developing relationships between the species composition of forage and the yield, the quantity of woody vegeta- tion and the soil erosion status of sample locations, using several large data sets similar to those described by Foran et al. (1986). If the relationship between composition and, for example, soil ero- sion status was linear, a gradual transition from good to poor condition was indicated; that is, there was no threshold. If on the other hand there was a discontinuity in the relationship, then a threshold was possible. The final test would be to follow monitor- ing locations through time to determine whether the poor condi- tion state was reversible, but assessment at a single point in time might provide a useful first indication.
For 21 relatively uniform range monitoring locations in a spar-
- good condition f poor condition
n atypical
-0 100 200 300 400 500 600 700 Yield(kg/ha)
Fig. 5. Reiztionship between specie-s composition end yield of forage in 8n arid open woodland range site. The ordinztion zcore which provided the composition index was adjusted to zero end multiplied X 1,000 to simplify presentation. Clusters of monitoring iocationz eszeszed in good condition (+) end poor condition (*) ue outlined to emphzsize their dizcontinuity. Remaining locations (a) are ztypicai, e.g., ecotonai, unusually wet.
sely treed arid woodland, assessed between July and September 1986, a potential threshold was indicated in the relationship between the forage yield and composition (Fig. 5). The ordination score of species composition was obtained from ordination of the range monitoring locations, using the Bray-Curtis metric in the TAXON package, as described earlier. Other attempts to detect thresholds at a single point in time were not successful due to site variability and the influence of the tree layer on forage composition.
A more promising approach is to follow the trend in forage composition over time. Foran et al. (1986) followed the time trajec- tories of a series of monitored areas on the same range site, traced through ordination space by a procedure similar to that described above for grazing gradients (Friedel unpubl.). They found that composition on poorly managed areas fluctuated much more than that on well-managed areas, and that the 2 areas occupied different parts of the ordination space (Fig. 6). The same behaviour was observed in the grazing gradient study (Fig. 3). More importantly, they showed that location No. 245 (Fig. 6) was shifting across the ordination space from the well managed area, dominated by palat- able Enneapogon spp. in summer and winter, to the poorly man- aged area, occupied by a fluctuating mix of short-lived species, in response to heavy grazing pressure. It was nearing or crossing a threshold and, at this point, management action was required to reverse the trend.
When quantitative methods are available for the assessment of soil condition, the status of soils may be treated similarly. Multiv- ariate analysis has already been used to reduce complex species composition data to a few functional groups (Friedel et al. 1988a). The potential exists to characterize thresholds in terms of the relative proportions of a few groups, under specified seasonal conditions.
-
Site No. 245
t2*
Fig. 6. Ordination of 10 monitoring locationsin an arid shrubby grassland range site, assessed on 4 different occasions (from Fonn et al. 1986). First year of data collection, 0, good condition state -, poor condition state - -, site (location) no. 245 -. Ordination axis 2 (x-axis) and
axis 3 (y-axis) are indicated.
The use of thresholds is compatible with the multivariate site potential approach of Wilson (1986). Climax is not identified. Instead, on the basis of potential for livestock production, areas are assessed to be on one side of a threshold or the other-they are in good or bad condition. If there are 2 thresholds, then 3 condition classes should be evident. The trend over time is determined by shifts in ordination space; departure from the pattern of seasonal responses shown by most monitoring locations signals the crossing of a threshold. Such a departure may be bad or good, and the management causes are worth investigating in either case.
Conclusiops
There are a number of problems associated with existing methods of range condition assessment, not the least of which is an inadequate theoretical base. As a consequence, assessments are produced that do not fit what we know to be happening in our rangelands. I have put forward the possibility of focusing on thresholds of change from one domain or state, to another. I propose that range does not necessarily deteriorate linearly as grazing pressure increases. Instead, it may retain the capacity to recover up to a critical point, beyond which it cannot readily return to its former state. Often some other factor, such as drought, fire, or torrential rain, coincides with excessive grazing to tip the balance.
Thus far, the proposition has had a preliminary testing in some arid rangelands. It requires much further testing in a variety of environments before we can be certain that it offers a real alterna- tive to existing approaches. Its greatest relevance may be in envi- ronments like the arid rangelands which are highly variable and unpredictable.
References
Bosch, O.J.H., F.P.J. van Rensburg,andS. duT. Truter. 1987. Identifica- tion and selection of benchmark sites on litholitic soils of the western grasslands biome of South Africa. J. Grassl. Sot. Southern Afr. 45962.
426
Bradbury, R.H., L.S. Hammond, R.E. Reichelt, and P.C. Young. 1983/4. Prediction versus explanation in environmental impact assessment. Search 14:323-325.
Dyksterhuis, E.J. 1949. Condition and management of range land based on quantitative ecology. J. Range Manage. 2: 104-l 15.
Form, B.D., C. Bastin, and K.A. Shaw. 1986. Range assessment and monitoring in arid lands: the use of classification and ordination in range survey. J. Environ. Manage. 22~67-84.
Friedel, M.H. 1985. The population structure and density of central Aus- tralian trees and shrubs, and relationships to range condition, rabbit abundance and soil. Aust. Rangel. J. 7: 130-l 39.
Friedel, M.H. 1987. A preliminary investigation of woody plant increase in the Western Transvaal and implications for veld assessment. J. Grassl. Sot. Southern Afr. 425-30.
Friedel, M.H., G.N. Bastin, and G.F. Griffin. 1988~. Range assessment and monitoring in arid lands: the derivation of functional groups to simplify vegetation data. J. Environ. Manage. 27:85-97.
Friedel, M.H., V.H. Chewings, and G.N. Bastin. 198813. The use of com- parative yield and dry-weight-rank techniques for monitoring arid range- land. J. Range Manage. 41:430-435.
Gauch, H.G. 1982. Multivariate analysis in community ecology. Cam- bridge Univ. Press, Cambridge.
Harrington, G.N., D.M.D Mills, A.J. Pressland, and K.C. Hodgkinson. 1984. Semi-arid woodlands, p. 189-206. In: G.N. Harrington, A.D. Wilson, and M.D. Young (eds), Management of Australia’s Rangelands. CSIRO Australia, Melbourne.
Hennessy, J.T., R.P. Gibbens, J.M. Tremble, and M. Cudenas. 1983. Vegetation changes from I935 to 1980 in mesquite dunelands and former grasslands of southern New Mexico. J. Range Manage. 36:370-374. Helling, C.S. 1973. Resilience and stability of ecological systems. Annu.
Rev. Ecol. Systematics 4: l-23.
Humphrey, R.R. 1949. Field comments on the range condition method of forage survey. J. Range Manage. 2:1-10.
Laycock, W.A. 1989. Secondary succession and range condition criteria: introduction to the problem. In: W.K. Lauenroth, and W.A. Laycock (eds), Secondary succession and the evaluation of rangeland condition. Westview Press, Boulder, Colo.
Laycock, W.A. 1991. Stable states and thresholds of range condition on North American rangelands: A Viewpoint. J. Range Manage. 44z427-433. Lauenroth, W.K., and W.A. Laycock. (eds.). 1989. Secondary succession
and the evaluation of rangeland condition. Westview Press, Boulder, Cola.
Noy-Meir, I., and B.H. Walker. 1986. Stability and resilience in rangelands, p. 21-25. In: P.J. Joss, P.W. Lynch, and O.B. Williams (eds), Range- lands: a resource under seige. Australian Acad. Sci., Canberra. Pendleton, D.T. 1989. Range condition as used in the Soil Conservation
Service. In: W.K. Lauenroth, and W.A. Laycock (eds), Secondary suc- cession and the evaluation of rangeland condition. Westview Press, Boulder, Colo.
Ross, D., M. Dale, K. Shields, and C. Hulett. 1983. TAXON use&manual, edn P4. CSIRO Div. of Computing Res., St. Lucia, Queensland. Smith, E.L. 1978. A critical evaluation of the range condition concept.
Inter. Congr. Rangel. Proc. 1st (Denver, Colo.) p. 266-267.
Smith, E.L. 1989. Range condition and secondary succession: a critique. In: W.K. Lauenroth, and W.A. Laycock (eds), Secondary succession and the evaluation of rangeland condition. Westview Press, Boulder, Colo. Stoddart, L.A., A.D. Smith, and T.W. Box. 1975. Range management.
McGraw-Hill, New York, 3rd edn.
Tainton, N.M. 1986. A system for assessing range condition in South Africa. Inter. Congr. Rangel. Proc. 2nd (Adelaide, So. Aust.) p. 524. Tainton, N.M., P.J. Edwards, and M.T. Mentis. 1980. A revised method
for assessing veld condition. Proc. Grassl. Sot. Southern Afr. 15:3742. Westoby. M. 1979/80. Elements of a theorv of veeetation dynamics in arid
rangeiands. Is&l J. Bot. 28:169-194. _ -
Westoby, M., B. Walker, and I. Noy-Meir. 1989. Opportunistic manage- ment for rangelands not at equilibrium. J. Range Manage. 42:265-273. Wilson, A.D. 1984. Points of reference in the assessment of change in
vegetation and land condition. Aust. Rangel. J. 6:69-74.
Wilson, A.D. 1986. The monitoring of changes in range condition: a multivariate site potential approach. Inter. Congr. Rangel. Proc. 2nd (Adelaide, So. Aust.) p. 517-521.
Wilson, A.D. 1989.The development of systems of assessing the condition of rangeland in Australia. In: W.K. Lauenroth, and W.A. Laycock (eds), Secondary succession and the evaluation of range condition. Westview Press, Boulder, Colo.
Wissel, C. 1984. A universal law of the characteristic return time near thresholds. Oecologia 65:101-107.