GENETIC CONTROL OF THE UPTAKE
OF
AMINO ACIDS IN ASPERGILLUS NlDULANSUMAKANT SINHAT
Department of Biochemistry, University of Leicester, University Road, Leicester, England
Received August 19, 1968
NALOGUES of some essential metabolites are inhibitory to a variety of
A
micro-organisms and these analogues are taken up into the cell by the same route and mechanism as their natural competitors (RICHMOND 1962). Selection for resistance to the structural analogues of metabolites as a means of obtaining mutants deficient in specific as well as general permeases have, therefore, been widely used, especially in studying the transport of amino acids. Such mutants provide the information regarding the genetic control and specificity of transport mechanisms.111 fungi evidence has been presented for lack of specificity as well as for multi- plicity of amino acid permeases (far review see GRENSON 1967). p-fluoro- phenylalanine
(FPA)
resistant mutants of A s p e r g i l h nidulans described in this paper appear to be deficient in the uptake of phenylalanine (PHE),
some otheramino acids and their analogues. Wild-type A . nidulans has an effective system
for taking up and concentrating these compounds from the medium. In the FPA- resistant mutants at the fpaD locus this uptake system is greatly reduced or is absent. This locus has been mapped in the ni7-arg3 interval of linkage group VI11 and has been found to be dominant or semidominant over its wild-type al- lele in heterozygous diploids. A preliminary report of some of these observations was presented earlier ( SINHA 1967a, b)
.
MATERIALS A N D METHODS
Media: Minimal medium (MM) was Czapek-Dox medium with 1% w/v glucose. Complete medium (CM) was a complex medium containing yeast extract, hydrolysed casein, hydrolysed nucleic acid, vitamins etc. Solid media contained 2% agar.
Strains: All the strains used were either from the stocks maintained in the Department of Genetics of the University of Glasgow or their derivatives. Mutants f p c r D l l and fpaC43 were isolated by MCCULLY (DORN 1967) and fpaD56 was isolated by SINHA (1967~). br42 was a mor- phological mutant (CLUTTERBUCK 1967).
Methods of genetic analysis: The general techniques described by PONTECORVO et al. (1953), PONTECORVO and KXFER (1958) and Gm (1958) were used. Assignment of genes to linkage groups and tests f a r translocations were carried out with the help of 'Master strain F' (MSF) as described by MCCULLY and FORBES (1965). Diploid strains were prepared from heterocaryons
by the method of ROPER (1 952). Gene symbols and linkage relationships of various loci have been listed by DORN (1967).
Preparntiom of hyphal suspensions for uptake experiments: 108 conidia of required strains
496 U M A K A N T S I N H A
were grown for 16-18 hrs at 37°C in shaken cultures (Gyrotory shaker-New Brunswick) in 400 ml liquid MM supplemented with required growth factors, contained in 2 1 baffled flasks. The hyphae were filtered on Buchner filters and washed with distilled water. Excess liquid was removed by suction filtration and the pads were suspended in 100 ml of distilled water. Such suspensions (3-6 mg dry weight/ml) were suitably diluted for uptake experiments.
Uptake measurements: Uptake was assayed by incubating suitable aliquots of the hyphal suspension with 3H-amino acids ( 1 0 - 4 ~ labelled with 0.1 pc tritium) in B shaking water bath. At the end of the incubation period 5 ml samples were filtered quickly on millipore filters (pore size 0.8 p, diameter 2.5 cm) and washed with 20 ml of distilled water. Each filter was then placed in 5 ml of BRAT’S mixture /BRAY 1960) contained in 15 ml scintillator vials and stirred thor- oughly on a Vortex mixer. Counting was carried out for 10 min in a Packard liquid scintillation counter.
Chemicals: All the chemicals used were of analytical grade and were obtained from com- mercial sources. 3H-amino acids were supplied by the Radiochemical Centre, Amersham, Bucks., U.K.
R E S U L T S
Haploid as well as heterozygous diploid strains for each of the four FPA- resistant mutants (fpaDI1, f p d 4 3 , fpuD56 and f p a A l ) were tested for resistance to several concentrations of FPA in MM by growth of point inocula of conidia. By this test, as well as by measuring the growth of heterozygous diploids on’MM and MM f 0.01% FPA, fpuDl2 and fpaD56 were found to be dominant and fpaC43 was found to be semidominant (Table 1 ) . f p a A l is a recessive mutation (SINHA 1967c) and has been included for comparison. All the haploid resistant strains grew at the highest concentration of FPA (0.1
%)
tested.Non-recovery of FPA sensitive recombinants from crosses of the types ( 1 ) fpuD56 X fpaDII, (2) fpaD56
x
fpaC43 and ( 3 ) fpaDll x fpaC43 (Table 2)TABLE 1
Dominance tests of FPA-resistant mutants
Concentration of FPA (in % ) required to inhibit the growth of a colony
Colony diameter (mm) 48 hrs after inoculation of heterozygous diploids Mutants
tested
Heterozygous Haploid diploid
on MM
+
on MM O . O l q & FPAfpaDii
>
0.1 0.1fpaD56
>
0.1 0.1fpaC43
>
0.1 0.05fpaAi
>
0.1 0.00521.5 20.5
21
.o
19.516.5 10.0
20.0 0.0
TABLE 2
Nonrecovery of FPA-sensitjve recombinants from crosses between fpaDl1, fpaC+3 and fpaD56
Segregation of markers FPA-resistant : Green :
FPA-sensitive Yellow Cmss G n o t y p s of parents
-
fpaDS6 X fpaDii bii;fpaD56 X pabai,adl7,y;fpaDl~ 203 : 0 92 : 111 194 : 0 93 : 101
fpaDil
x
fpaC43 riboi,bil;fpaDli X pabai,adl7,y;fpaC43 208 : 0 113 : 95A M I N O A C I D U P T A K E I N A S P E R G I L L U S
TABLE 3
Meiotic location of fpaD11, fpaC43 and fpaD56
49 7
Percentage recombination fraction between
Number of br42-fpaD ni7-fpaD fpaD-arg3
Cross progeny tested ni7-br42 br42-fpaC ni7-fpaC fpaC-arg3
bil;arg3 x pabal,adI7,y;fpaC43 239 16.7 i. 2.2
4.9i.1.5 12.6-+2.3 . . .
bif;br42 X y;ni7,fpaC43 206 . . .
. . . 18.322.8 riboI,bii;fpaDII
x
y;ni7 186. . .
bil;ni7 x pabai,adl7,y;fpaC43 195 . . 22.0k2.9 . . .
biI;fpaD56 x y;ni7 203 . . . 17.7k2.7 . . .
. . .
suggested that the isolates f p a D l l , fpaC43 and fpaD56 are either allelic to each other or are located very close to one another.
A heterozygous diploid between ribol,
bil;
fpaDll and ‘Master strainI?
was synthesised. It was haploidised on F’PA and showed that the strain ribol, bil; fpaDll is free of translocations and the locus fpaD is in linkage group VIII.By appropriate crosses the mutants fpaDl1, fpaC43 and fpaD56 were located at about the same place in the ni7-arg3 interval (Table 3 ) . In view of the mapping results these three mutants can be assigned to a single locus-fpd. These results differ from those of MCCULLY (DORN 1967) in that fpaDll is not in linkage group I11 and that mutants fpaDll and fpaC43 are probably allelic.
The results of five crosses presented in Table
4
show that there is some inter- action between the FPA-resistance of a mutant at the fpaD locus and PHE or methionine requirements of relevant auxotrophs. The apparent distortion for arg2 in the third cross is most probably due to its tight linkage to the meth2 marker. There is no interaction between FPA-resistance and riboflavin or biotin requirements or between FPA-resistance and inability to utilize nitrate or sul-TABLE 4
Segregation of nutritional markers with respect to fpaD and fpaC
Cross
Number of Pairs of Segregation classes PW5WY markers
tested considered
+ +
+ - - +
-- bil;cnxH4 x pabal,adl7,y;fpaD11 208 fpaDll -cnxH4 (63) 51 50 (44) y ; d 2 x ribo1,biI;fpaDll 201 fpaD11-sI2 (61) 29 67 (44) fpaDl I-ribol 51 (39) (66) 45 fpaDII -bil 45 (42) (4t) 67 y;argZ,methZ x ribo1,biI;fpaDlI 198 fpaDlI-arg2 (75) 47 71 (05)fpaDlI-meth2 (71) 51 72 (04) fpaDII-ribol 61 (61) (48) 28
f paDl I - biz 51 (71) (36) 40 y;methZ,phenAZ x riboI,bil;fpaDlI 208 fpaDlI-meth2 (59) 80 51 (18)
fpaDII-phenA2 (68) 71 69 (00) biI;arg3 X paba1,adf 7,y;fpaC43 299 fpaC43-arg3 (23) 136 113 (30)
498 U M A K A N T S I N H A
FIGURE 1.-Segregants from a cross y;meih2,phenA2
x
ribo1,bil;fpnDll on MM -k PHE -k methionine+
riboflavin+
biotin. Arrows point to melh2;fpoDi 1 recombinants.phate. Very few meth2; fpaDll recombinants were obtained. Going back to the colonies recovered by plating the ascospores of a hybrid perithecium from the cross y; meth2, phenA2 X ribol,
hil;
fpaDll on MMf
all the required growth factors, it was noticed that even after 72 hrs of incubation meth.2; fpaDll re- combinants formed very small, poorly sporulating and cottony colonies (Figure 1 ).
On prolonged incubation such colonies often produced well sporulating and normally growing sectors which turned out to be methionine-independent and FPA-resistant. Perhaps poor growth, poor sporulation and difficulty in transfer to the tester media tend to eliminate the meth2; fpaDll recombinants at each of the successive steps of the master plate preparation, transfer to the tester media and scoring. On the other hand the recombinant phenA2; fpaDll was not re- covered at all. Numerical results of Table 4 strongly suggest that the absence of this recombinant class is due to its inviability on MM f required growth factors rather than due to suppression ofPHE
requirement.All three mutants at the fpaD locus isolated so far were found to be resistant to FPA, ethionine and L-3-aminotyrosine
+
phenylanthranilic acid, whereas theA M I N O ACID UPTAKE I N ASPERGILLUS 499
100
h
-I
a
2 80
>
nz 3
Y,
w 6o
s
Iz
5
2 0(3
5
40 w0
0 1 2 3 4 5 6
F P A in mg Iml.
FIGURE 2.-Percentage survival of conidia of f p ~ & , b i i ( 0 ), riboi,bii;fpaDil (O), bii; fpaD56 (H) and n'bol,bil;fpaC43 (0) on MM f biotin
+
riboflavin-+
different concentrations of FPA after 48 hrs of incubation.sine
f
phenylanthranilic acid has enabled CALVORI and MORPURGO (1966) to select a class of mutants resistant to these antimetabolites as well as to FPA. The properties of these mutants, so far investigated, appear to be similar to thoseof the mutants at the fpaD locus.
Biochemical investigations: The total absence or marked deficiency, in genetic analyses, of the classes phenA2; f p a D l l and met&?; fpaDI1 and resistance of fpaD mutants to FPA, ethionine and L-3-aminotyrosine
+
phenylanthranilic acid suggested that they may well have an impaired capacity for uptake of PHE and related compounds from the medium. Direct measurements of the uptake of3H-
amino acids by wild type (ribol, b i l ) and mutant (ribol, bil; fpaDl1) strains established that this was the case. As shown in Figure 3, the wild-type mycelium takes up
PHE
very rapidly. The internal concentration reaches a maximum after about 50 min. PHE uptake by the mutant proceeds at a very low rate. Three independently isolated fpaD mutants ( f p a D l l , fpaC43 and fpaD56) were com- pared to the wild type (ribol, b i l ) for PHE uptake. All had rates less than 15% of that of the wild type. Suspension of hyphae in 1 0 - 2 ~ diamino-ethane-tetraacetic acid for 90 min had no effect on the uptake of PHE in any of the strains tested. Several other amino acids appear to be taken up by the same uptake system as PHE. The results given in Table5
show that a mutation at the locus fpaD dis- tinctly retards the uptake of tyrosine, tryptophan, methionine, leucine and as- partic acid and slows down the uptake of histidine, glycine, glutamic acid and lysine by about one fourth the rate in the wild type. Table 6 shows the blocking of uptake of 3H-PHE by wild type with other amino acids which are unlabelled. It can be seen that the most effective blocking amino acids are the same ones which depend on the presence of the fpD+ allele for their own uptake.500 UMAKANT S I N H A
TIME (minub)
FIGURE 3.-Time plot of the uptake of PHE by hyphal suspensions of ribo1,bil ( 0 ) and
ribo1,bil;fpaDll (U).
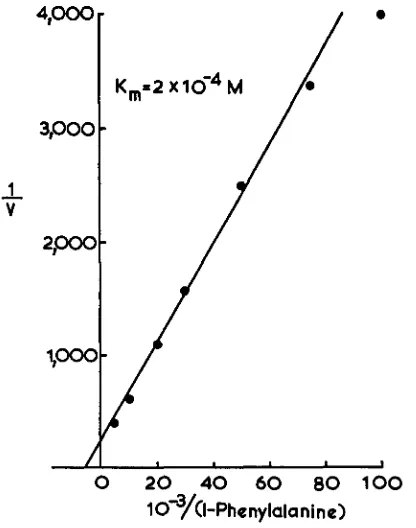
20 min following the addition of the amino acid. Initial velocities of the entrance of 3H-PHE were, therefore, studied for the first 10 min of incubation after the addition of the isotope. Such studies in the strain ribol, bil appear, on a Line- weaver-Burk plot, to indicate that the uptake of PHE is dependent on only one function and is saturable. This suggests that PHE enters the hyphae by only one system, the
K ,
and VJ1lUx of which would be about 2 x IO-'M and 52 x pmoles/mg dry weight/minute, respectively (Figure4).
Similar studies with the mutant ribol, bil; f p a D l l suggest that the velocity of entrance of "-PHE isTABLE 5
Uptake of various amino acids By ribo1,bil;fpaDll (Percentage of ribo1,bil)
Amino acid Percentage uptake
phenylalanine 11
tyrosine 20
tryptophan 12
aspartic acid 12
glycine 21
glutamic acid 2.3
lysine 25
methionine 16
leucine 16
histidine 30
valine 80
threonine 83
A M I N O A C I D U P T A K E IN A S P E R G I L L U S
TABLE 6
Effects of p-fiuorophenylalanine, ethionine and other amino acids on the
uptake of phenylalanine b y ribo1,bil
501
Blocking agent
Percentage uptake of
phenylalanine at concentrations
I O - 4 ~ 2x 1 0 - 4 ~
none p-fluorophenylalanine ethionine glutamine alanine aspartic acid asparagine tryptophan serine threonine leucine tyrosine glycine methionine histidine lysine valine arginine isoleucine glutamic acid proline cysteine 100 8 10 9 9 10 12 17 17 17 1 7 17 19 20 31 41 43 52 60 66 1 1 1 3 4 100 31 47 46 73 31 67 59 53 74 5 4 80 50 77 69 82 84 65 77 83 94 62
Hyphae were in SH-PHE for 10 min before sampling, The concentration of the unlabelled blocking amino acid analogue or amino acid was 4 x 1 0 - 3 ~ .
greatly reduced in this strain (Figure
5 ) .
This could be due to a passive o r re- tarded uptake system. Figure 6 shows the effect of varying the external PHE concentration on the rate of its uptake in a ribol, bil strain.D I S C U S S I O N
The results presented here indicate that three independently isolated FPA- resistant mutants f p D l l , fpaC43 and fpaD56 are allelic to each other and the locus fpaD is in the ni7-arg3 interval in linkage group VIII.
502 UMAKANT S I N H A
IO-?( I
-Phenylanlne)
FIGURE 4.-Lineweaver-Burk plot of the uptake of PHE by hyphal suspensions of ribo1,bil. U in pmoles per mg dry weight per min.
4,000
-
e1
V
/
t 1 I0 2 0 4 0 6 0 8 0 1 0 0 1 04/(l-Phenylalani ne)
FIGURE 5.-Lineweaver-Burk plot of the uptake of PHE by hyphal suspensions fpaD11. U in pmoles per mg dry weight per min.
A M I N O A C I D U P T A K E I N A S P E R G I L L U S
503
'10.0
:
2.5 'J
e
8
+I
c
w
U
c
1.25
n
0 1 0 2 0 3 0
TIME (minutes)
FIGURE 6.-Time plot of the uptake of PHE by hyphal suspensions of ribo1,bil in the presence of several external PHE concentrations.
f p a D l l takes up practically no 3H-PHE. Kinetic parameters of PHE uptake in the wild type as well as in the mutant support this contention. Mutations with similar broad specificities exist in yeast (SURDIN et al. 1965) and Neurospora (JACOBSON and METZENBERG 1967).
It is not clear at this stage whether f p a D is the structural gene for a permease, and, if not, what is its relation to the uptake system. The structural gene hy- pothesis is favoured by the qualitative alterations of the uptake system controlled by different mutations at the f p a D locus. But dominance or semidominance of such mutants in heterozygous diploids point to a regulatory role of this locus.
CALVORI and
MORPURGO
(1966) observed that suppressors of pfp-r ( f p a A ) can be distinguished from their true revertants (1) by crossing the former (pfp-r, su-pfp-r) with a wild-type strain and selecting FPA-resistant segregants or (2)by examining diploids of the general structure
PfP-rx
+
su-pfp-rx+
PfP-rv+
which, on the assumption that the suppressor is recessive, should be FPA-resistant or (3) by their capacity to grow on media supplemented with either FPA or L-3- aminotyrosine
+
phenylanthranilic acid. Similar results will be obtained if one assumes that their su-pfp-r mutants are in fact mutants at the f p a D locus. Un- fortunately mapping results of su-pfp-r mutants are not available to verify this suggestion.5 04 UMAKANT S I N H A
with the results of MCCULLY (unpublished results) and DORN (1967). However, the locus symbol
‘D’
has been retained and in view of the fact that fpaDll and fpaC43 appear to be allelic, it is suggested that the isolate fpaC43 also should be given the locus symbol ‘D’ instead of ‘C’ which was proposed by MCCULLY (un- published results). Semidominance of fpaC43 in a heterozygous diploid and its lesser degree of FPA-resistance (as compared to fpaDII and fpaD56) can be at- tributed to an incompleteness of the genetic block in this mutant.Mutants of A. nidulans at loci fpaA ( t y r A ) and fpaE ( t r v p A ) have been in- terpreted to be FPA-resistant due to an oversynthesis of
PHE
(SINHA 1 9 6 7 ~ ) . Loss or retardation of the uptake of amino acids and their analogues appears to be another mechanism for FTA-resistance in this mould. Most of the mutants iso- lated so far as resistant to FPA alone are probably excretor mutants (SINHA1 9 6 7 ~ ) . Amino acid permease mutants can probably be better selected on a me- dium containing more than one amino acid analogue.
Kinetic parameters of
PHE
uptake in the wild type as well as in the mutant suggest that only one uptake system is responsible for concentrating PHE from the medium into the cell of A . nidulans. The same system which takes up PHE in Aspergillus is also responsible for the uptake of some other amino acids. Thus it is not surprising that fpaD mutants are resistant to a number of amino acid ana- logues.I am very grateful to Prof. G. PONTECORVO, F.R.S. for suggestions and criticisms throughout the course of this investigation and to Dr. J. M. AsHwoRrIi for providing the facilities for uptake experiments. Part of this work was done in the Department of Genetics of the University of Glasgow. This work was conducted during the tenure of a Commonwealth Scholarship and a Science Research Council Post-Doctoral Fellowship (under Grant No. B/SR/4360) both of which I acknowledge with thanks.
SUMMARY
I n Aspergillus nidulans p-fluorophenylalanine-resistant mutants at the fpaD locus have been found to be simultaneously resistant to ethionine and L-3-amino- tyrosine f phenylanthranilic acid and to be dominant or semidominant over their wild-type alleles in heterozygous diploids. The locus fpaD has been mapped in the ni7-urg3 interval of linkage group VIII. On the basis of genetic as well as b;ochemical studies it has been suggested that the mutants at this locus have an impaired capacity for the uptake of several amino acids and their analogues. The wild type has been found to transport phenylalanine at a rapid rate and this sys- tem has a K , of 2 x 1 0 4 ~ and Vma” of 52 x p moles per mg dry weight per min. A number of amino acids have been found to compete with phenylalanine for the same uptake system.
LITERATURE CITED
BRAY, G. A., 1960
CALVORI, C., and G. MORPURGO, 1966
A simple efficient liquid scintillator for counting aqueous solutions in a
Analysis of induced mutations in Aspergillus nidulans. liquid scintillator counter. Anal. Biochem. 1 : 279-285.
A M I N O ACID U P T A K E I N A S P E R G I L L U S 505
A mutational analysis of development in Aspergillus nidulans.
A revised map of the eight linkage groups of Aspergillus nidulans. Genetics
Le transport des m4tabolites-en particulier, des acides amines-a travers
A new gene which affects uptake of neutral and CLUTTERBUCK, A. J., 1967
Heredity 22: 459. DORN, G. L., 1967
56: 619-631. GRENSON, M., 1967
les membranes cellulaires. Rev. Fermentations. Bruxelles.
JACOBSON, E. S., and R. L. METZENBERG, 1967
acidic amino acids in Neurospora crassa. Biochim. Biophys. Acta 156: 140-147. KAFER, E., 1958
MCCULLY, K. S., and E. FORBES, 1965 The use of p-fluorophenylalanine with ‘master strains’ of
PONTECORVO, G., and E. KAFER, 1958 Genetic analysis based on mitotic recombination. Advan.
PONTECORVO, G., J. A. ROPER, L. M. HEMMONS, K. D. MACDONALD, and A. W. J. BUFTON, 1953
RICHMOND, M. H., 1962 The effect of amino acid analogues on growth and protein synthesis in
ROPER, J. A., 1952 Production of heterozygous diploids in filamentous fungi. Experientia 8 :
SINHA, U,, 1967a Genetics of aromatic amino acid biosynthesis in Aspergillus nidulans. Ph.D. Thesis, University of Glasgow. - 1967b Genetics of aromatic amino acid biosynthesis in Aspergillus nidulans. Heredity 22: 457. - 1967c Aromatic amino acid biosynthesis and parafluorophenylalanine resistance in Aspergillus nidulans. Genet. Res. 10 : 261-272.
Propri6tBs et controle g6n6tique du syst+me #accumulation des acides amines chez Saccharomyces cereuisiae. Biochim. Biophys. Acta. 107 : 546-566.
An 8-chromosome map of Aspergillus nidulans. Advan. Genet. 9: 105-145.
Aspergillus nidulans for the assigning genes to linkage groups. Genet. Res. 6: 352-359.
Genet. 9: 71-104.
The genetics of Aspergillus nidulans. Advan. Genet. 5 : 141-238.
micro-organisms. Bacterid. Rev. 26: 398-420.
14-15.