Copyright 0 1989 by the Genetics Society of America
Genetic Analysis of Olfactory Behavior in Drosophila:
A New Screen
Yields the
ota
Mutants
Craig Woodard, Thomas Huang, Henry Sun,' Stephen L. Helfand and John Carlson
Department of Biology, Yale University, New Haven, Connecticut 0651 1 Manuscript received March 3 1, 1989
Accepted for publication June 12, 1989
ABSTRACT
A simple means of measuring Drosophila olfactory response is described, and the behavior which
it measures is characterized. The assay was used to screen for X-linked mutants defective in olfactory function. Six ota mutants were isolated and characterized (ota = olfactoy trap abnormal). Four of the mutants were found to be abnormal in another chemosensory behavior as well. Two of the mutant phenotypes extend to include another sensory system: they are defective in visual system physiology. All were normal, however, in a test of giant fiber system physiology. Two of the mutations are dominant, and the recessive mutations define two complementation groups. Mutations representing each complementation group, as well as one of the dominant mutations, were mapped. For the mutants with defective visual system physiology, the visual defects were shown to cosegregate with olfactory phenotypes.
D
ROSOPHILA is exquisitely sensitive to a wide variety of volatile chemicals, which it senses primarily with its antennae (BARROWS 1907; VENARD and PICHON 198 1). The third segment of each an- tenna is covered with approximately 500 sensory hairs, which can be divided into three morphologically distinct classes: sensilla basiconica, sensilla coelocon- ica, and sensilla trichodea. Each class is distributed in a characteristic pattern on the antennal surface (VEN-KATESH and SINGH 1984), and each hair contains
up
to four neurons, which project directly to the antennal lobes of the brain.
Little is known about sensory transduction, func- tional organization, or development of the Drosophila olfactory system. Although belief in the existence of olfactory receptor molecules has enjoyed common currency in the field of chemosensory physiology for many years (SHEPARD 1983), receptor molecules have not yet been identified or characterized in any higher organism. The initial steps of olfactory transduction can be elucidated only after characterization of those membrane components with which chemical stimuli interact. The logic by which Drosophila or other animals encode complex olfactory information into a form interpretable to the brain is poorly understood; the organization of olfactory systems has not been described in the detail provided by HUBEL, WIESEL, and others for the mammalian visual system (HUBEL and WIESEL 1977). Finally, little is known about the principles by which genes specify the construction of the system in Drosophila nervous system develop- ment.
' Present address: Institute of Molecular Biology, Academia Sinica, Nan- kang, Taipaei, 11529 Taiwan, Republic of China.
The publication costs of this article were partly defrayed by the payment of page charges. This article must therefore be hereby marked "aduertismncnc"
in accordance with 18 U.S.C. 61734 solely to indicate this fact. Genetics 123 315-326 (October, 1989)
Genetic analysis of mutants defective in visual be- havior (BENZER 1967) has been instrumental in inves- tigating both the mechanisms by which Drosophila responds to visual information and the means by which the visual system develops. Comparable genetic analysis of olfactory mutants has been pioneered by RODRIGUES AND SIDDIQI (1 978), who have described the isolation of several olfactory system mutants [SID-
DIQI (1987); see also HELFAND and CARLSON (1989)
and MCKENNA et al. (1989)l. Progress in this field, however, has been constrained by lack of convenient methods of measuring olfactory behavior. To screen for mutants, RODRIGUES AND SIDDIQI used a Y-maze
assay, in which a population of adult Drosophila makes a choice between two air streams, one containing an odorant, the other a control. Such assays enjoy impor- tant advantages, but also suffer certain limitations: theoretical limitations are imposed by the binary- choice design, practical limitations are imposed by the testing of large populations of animals, and technical limitations are imposed by the need for fine manipu- lation of airflows.
We have developed a new means of measuring the response of Drosophila to airborne chemicals. The method is simple, requires few animals, and yields a high signal-to-noise ratio. We describe here the olfac- tory behavior which it measures and a screen for mutants in which this behavior is defective. Six mu- tants defective in olfactory response were isolated and characterized.
MATERIALS AND METHODS
Drosophila melanogaster stocks and mutagenesis: Can-
from a Canton-S stock obtained from 0. SIDDIQI, Tata Institute, Bombay. CS-5 was generated from this Canton-S stock and from an FM7 balancer stock whose autosomes were of Canton-S origin, obtained from S. BENZER, Califor- nia Institute of Technology. The CS-5 stock was mutagen- ized by means of EMS (LEWIS AND BACHER 1968), X- irradiation (4300 rad), or hybrid dysgenesis. In the case of EMS or X-rays, mutagenized males were allowed to mate en masse with attached-X females for a period ending 4 days after the beginning of the mutagenesis procedure. F1 males were then individually mated to attached-X females to es- tablish lines, which were then screened in the FS or in subsequent generations. The mutagenized Fo males were limited to a 4-day mating period so that lines established from different F1 males would contain independently de- rived mutagenized X chromosomes. Attached-X virgin fe- males were obtained originally from a C ( I ) A y stock and subsequently from a C( I ) A y/shi" stock, both in a Canton-S background and both from D. KANKEL, Yale University. A single parental X chromosome and autosomes from a de- fined background were used in these screens in order to reduce variation in olfactory response due to genetic back- ground effects (MONTE et al. 1989). For hybrid dysgenesis, CS-5 was used as an M strain. We confirmed its cytotype to be M by the snw test (ENGELS 1979), and it was shown to be free of P elements by genomic blotting. It was crossed to five P strains which we confirmed to contain transposase activity: Harwich, obtained from S. ARTAVANIS-TSAKONAS, Yale University, and r2, 78-61, 8-31-15, and Inbred Cage 3, from W. ENGELS, University of Wisconsin. The duplica- tion y2Y611 (CRAYMER AND ROY 1980) was obtained from R. FLEMING, Yale University. Dp(l;f)fJ9 (HARDY et al. 1984) was obtained from A. SCHALET, Yale University.
Cultures were grown in half-pint milk bottles with card- board stoppers or in cylindrical vials with cotton stoppers. Drosophila were kept on cornmeal-molasses-agar medium (DOANE 1967), supplemented with dry active yeast. The animals were grown in 25 O incubators in the dark.
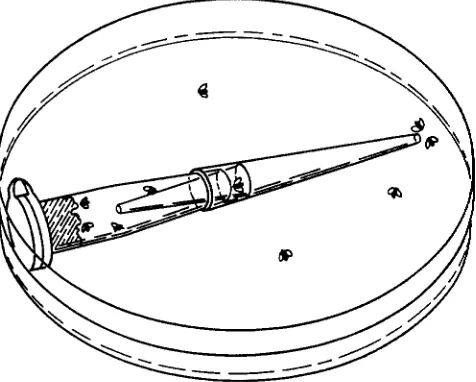
Trap assay: Trap assays were conducted in a Petri dish (Falcon: 100 mm X 20 mm) containing 10 ml of 1% agarose. Traps were constructed from a microfuge tube (Marsh Biomedical: 1.5 ml polypropylene, with flat attached cap) and two yellow plastic micropipette tips (USA/Scientific Products USA 1 10: yellow, 1-200 PI). The end of the micro- fuge tube was severed with a razor blade approximately 2.5 mm from its terminus so as to create an aperture of internal diameter 4 mm. When testing males, the terminal portion of each micropipette tip was removed by severing at a distance 0.8 cm from the end to create an internal diameter of -1.3 mm. When testing females, the distance was in- creased to 0.9 cm to create a larger aperture. One micro- pipette tip was further severed at the junction between the smooth and ridged portions of the tip, i e . , at a distance of 1.6 cm from the large end. This twice-cut tip was inserted snugly into the microfuge tube with its small end inside; the larger tip was fitted tightly over this with its small end outside (Figure 1). Approximately 400 PI of olfactory at- tractant was placed inside the microfuge tube: Drosophila culture medium was added in solid form simply by opening the microfuge cap and inserting with a small spatula; 0.5% agarose was added in liquid form through the hole made at the distal end, prior to the addition of the micropipette tips. When a pure chemical was used as an attractant, it was mixed with molten agarose maintained at 50". The mixture was then immediately transferred into the microfuge tube.
Following light anesthesia with COS, ten flies of the same sex and genotype were introduced into a Petri dish contain- ing a trap, and the number of trapped flies was determined
periodically until a score was finally determined at approx- imately 60 hr in the case of males and 85 hr in the case of females. The possibility that the C o n treatment might affect olfactory response was tested, and no adverse effect was found. During the course of the assay it was common for a small number of flies to die before entering the traps, presumably due to starvation. In one experiment, in an attempt to reduce the death rate, 1 % sucrose was added to the plates. However, this procedure was found, interest- ingly, to reduce markedly the number of flies entering the traps, from 8.5
*
0.4 to 2.7 & 0.6 ( N = 10; P < 0.001; the attractant was culture medium), as if the olfactory attraction response were decreased by the availability of local food resources. Viability of females was found to be much higher than for males, in that a much higher fraction of those females remaining outside a set of ethyl acetate traps at 60 hr was still alive (P < 0.001).The rate of exit from traps has been determined to be null or very low in experiments of three different designs. In all of these experiments, male flies were placed in empty traps and their rate of egress into the external chamber was then measured as a function of time. In one such experi- ment, a single fly was placed in each of a set of traps, in the second experiment ten flies were placed in each of a series of traps, and in the third experiment ten visibly marked flies
(w, sn) were placed in traps and ten wild-type flies were placed in the Petri dish outside the traps. At the end of a -60 hr test period, all of the singly trapped flies in the first experiment were still trapped ( N = lo), an average of 8.2 +: 0.5 flies were still trapped in the second experiment ( N =
5), and an average of 9.8 f 0.2 marked flies were still trapped in the third experiment ( N = 6). Since observation of these experiments was not continuous, but rather periodic, we cannot exclude the possibility that flies often exited from the traps and reentered thereafter. Arguing against this possibility, however, is the following fact: of a total of 159 observations made in these experiments on 21 traps, in oniy two cases was the number of experimental flies outside a trap observed to be less than at the time of the previous observation.
Age was determined not to be a significant factor in response to culture medium among six sets of males varying in age from 18 f 6 hr to 2 weeks at the onset of the assay. In most experiments, animals less than 1 week in age were used.
Manipulation and examination of antennae: A micro- pipette tip was placed under a dissecting microscope with the tapered end upward, in the beam of a light source positioned above (R. AYER, P. MONTE and J. CARLSON, in press). A fly, inserted into the base of the micropipette tip by means of a standard mouth aspirator, walked upward, presumably driven by positive phototaxis and negative geo- taxis. Air pressure was then applied with the mouth aspirator in order to lodge the fly at the top of the pipette tip, its head protruding through the opening of the tip and its thorax resting beneath the aperture. In this position, anten- nae or third antennal segments were removed with fine forceps. Also in this position, ota mutants were scored for gross defects in antennal structure.
Enrichment for olfactory mutants: Mutageneses I1 and
Genetics of Drosophila Olfaction 317
FIGURE 1 .-The trap assay. Ten flies are placed in the Petri dish along with a trap containing an olfactory attractant. The trap is constructed from a microfuge tube and two micropipette tips. The number of flies in the trap is counted as a function of time.
to reach the stimulant. After 10 h in the dark, approximately 90% of the larvae were found in the small dish; the larvae remaining outside were collected. Those individuals that pupated and eclosed, approximately 10,000 total for both mutageneses, were placed in population cages (12 cm di- ameter X 14 cm high), each containing a culture bottle. Flies could gain access to the medium in the culture bottle oniy by passing through a funnel whose terminus was 2 mm in diameter. Flies found outside the bottle after approxi- mately 3 days were subjected to another cycle of this pro- cedure, and attached-X lines were established from the 300 male flies failing to become trapped in both cycles. In the case of the dysgenic lines, males were backcrossed for three generations to an attached-X stock confirmed to be of P cytotype.
Physiology: Electroretinograms (see P A K AND GRA-
BOWSKI 1978) were recorded from flies attached to a wire
screen following light anaesthesia with ether. Glass record- ing and reference electrodes-both containing 3 M KC1 in initial experiments and Ringer's solution in subsequent ex- periments-were placed slightly below the cornea and in the thorax, respectively. Ag-AgCI electrode holders were used to connect the recording electrode to a high impedance preamplifier and the reference electrode to a ground. A Tektronix 5 1 13 or 2220 oscilloscope visualized the voltage signal. A quartz halogen illuminator and an electronic shut- ter were used to deliver single light pulses of 0.5-sec dura- tion.
Giant fiber pathway physiology was recorded as described by TANOUYE AND WYMAN (1980), except that flies were
immobilized using a small suction tube. The electrical stim- ulus delivered by the electrodes in the brain was 10-1 5 V. The electrical responses from the dorsal longitudinal and tergotrochanteral muscles were measured on a storage os-
cilloscope. Electrodes were made of tungsten. Three indi- viduals of each genotype were examined.
Statistical analysis: Statistical significances of differences between two means were determined using Student's t test, two-tailed. ANOVA was used for comparison of more than two means and was also two-tailed. Trap assay scores and chemosensory jump scores were arcsine transformed prior
TABLE 1
Summary of trap assay results
Time Experimental Control Signal: Series Stimulus (hr) score' score",* Noise
I Medium 72 8.2 f 0.3 (65) 0.77 f 0.3 (35) 10.6
I1 Various' -60 7.3 f 0.3 (63) 0.76 f 0.2 (21) 9.6
HId Medium 60 7.8 f O . l (413) IV Medium 60 7.1 f 0.2 (227)
Vd EtAc 60 5.9 f 0.2 (500) 0.40 f 0.4 (5) 14.8
a Mean f SEM (no. of trials). All flies tested were male.
Control traps contained agarose in series I and V; they were empty in series 11. Flies used in control experiments in series V were not mutagenized.
' Included varying combinations of medium and yeast, natural food sources, and virgin females.
Screening of lines established following EMS mutagenesis. Screening of lines established following mutagenesis with X-
irradiation or hybrid dysgenesis and enrichment (see text).
to comparison. All indicated experimental errors represent SEM.
RESULTS
Rationale in design of paradigm: A major consid- eration in the design of the paradigm described here is the signal-to-noise ratio. If signal and noise are defined as the fraction of animals responding in the presence and absence of stimulus, respectively, then the maximal signal-to-noise ratio obtainable with a binary choice assay is 1.0/0.5 =
2;
that is, whereas under optimal circumstances 100% of the animals will respond to a stimulus, in the absence of a stimulus 50% of the animals will make the appropriate choice by chance. Assays with higher signal-to-noise ratios offer several advantages, including the possibility that they may be adapted to efficient enrichment proto- cols. In a conventional Y-maze assay (e.g., RODRICUES and SIDDIQI 1978), for instance, at least 50% of mu- tants will be lost in each cycle of enrichment. By varying the parameters of the signal-to-noise ratio, however, enrichment protocols may be made more efficient.The trap assay was designed with the intention of providing the animals with a challenge resembling one for which the olfactory system presumably evolved. The system plays a role in the location of food re- sources, and the assay represents an effort to simulate this activity as closely as possible within practical limits. The physical dimensions of the assay and the materials chosen for it were selected so that it could be conven- iently used as the basis of a screen for mutants.
Llght Dark
TIME (HRSI TIME (HRS) TIME (HRS) TIME (HRS)
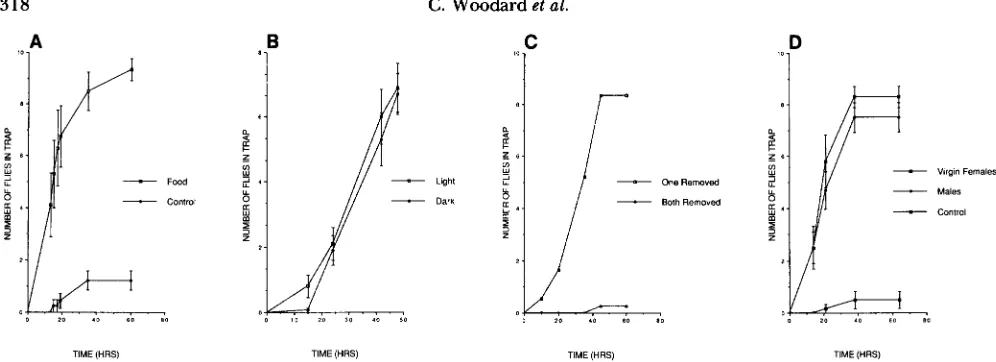
FIGURE 2.-Characterization of response in trap assay. Tested flies are Canton-S males in groups of ten. Error bars represent SEM. A) Response depends on presence of olfactory stimulant. Traps contain either Drosophila culture medium (Food) or no stimulant (Control). N
= five groups of ten flies. B) Response to culture medium does not require visual input. Assays were conducted under continuous light ( N = 15) or darkness ( N = 14). Time points for dark assays were taken in dim red light. Scores are not significantly different by Student's t-test. C) Response to culture medium requires antennal input. Third antennal segments were removed either bilaterally (Both Removed; N = 4) or unilaterally (One Removed; N = 2). D) Flies as attractants. Ten virgin females or males were used as attractants; they were marked with
w , sn so they could be distinguished easily from tested males and had been aged 2 days, 12 hr, at 18" at the onset of the experiment. During the course of the experiment, 2 of 120 w , sn animals, both male, were found to have escaped from the traps; one subsequently reentered. Control traps contained no attractant. In each of the three cases, N = 6.
which holds olfactory attractant. Flies can reach the attractant only by passing through a narrow orifice: the small end of the microfuge tube is cut off and a truncated pipette tip is placed with its small end inside the microfuge tube. Another truncated pipette tip is placed over the first with its small end outside. The pipette tip with the small end outside makes it difficult for a
fly
to enter the trap, and the pipette tip with the small end inside makes it hard for the fly, once en- tered, to exit. Response is measured by counting the number of flies in the trap as a function of time. The number of trapped flies is the integral of the entry rate minus the exit rate:Number trapped =
J &
(entry rate-
exit rate) d t .The exit rate is kept very low by the small aperture of the internal pipette tip, thus the number of trapped flies is a good measure of the entry rate (see MATE- RIALS AND METHODS for documentation). The signal is increased by allowing the flies an extended period of time in which to respond to the attractant, and the noise is kept low by virtue of the small aperture of the external pipette tip. The signal-to-noise ratio and ki-
netics of response in a typical experiment is shown in Figure 2A. We note that assays of a related design, although of much larger scale, have been used previ- ously (BARROWS 1907).
Characterization of behavior in the trap assay: A data base has been collected from four different series of assays and is shown in Table 1. The data show that on average approximately eight of the ten tested male flies are found in the trap at the end of the test period when Drosophila culture medium is used as the olfac- tory attractant. In series I, 8.2 A 0.3 (SEM; N = 65)
flies were found in food traps, whereas in a control set of traps containing only agarose, only 0.8 A 0.3 flies were found, indicating a signal-to-noise ratio of 10.6. These results are very similar to those obtained by pooling the data from a series of pilot experiments (series 11) in which various experimental conditions were used and in which the control traps were com- pletely empty. The largest data sets come from screens of lines established following EMS mutagenesis. Al- though the flies used in these assays had been heavily mutagenized, the average score was 7.8 (series 111).
Genetics of Drosophila Olfaction 319
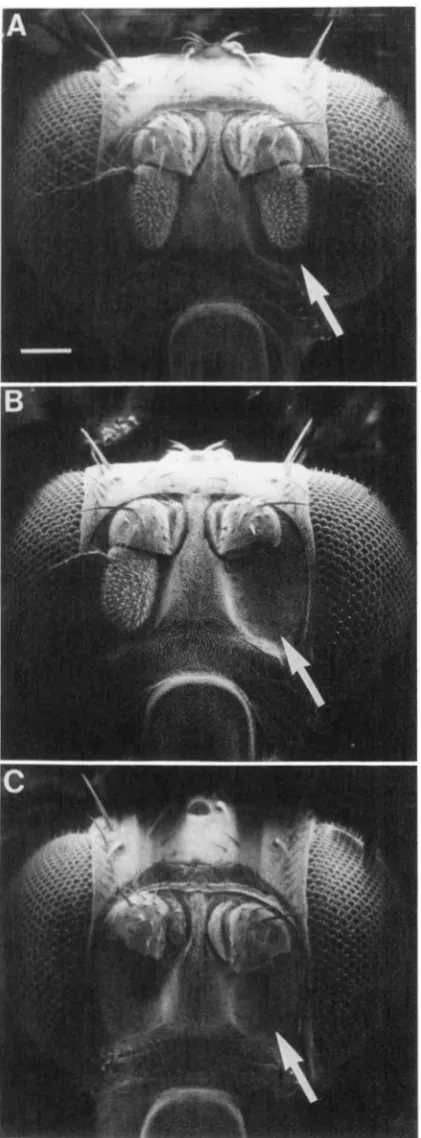
FIGURE 3.--Scanning electron micrograph of Drosophila head. Arrow indicates position of the third antennal segment. A) U n o p erated; B) following surgical removal of left third antennal segment; and C) following removal of both third antennal segments. White bar in (A) represents 100 pm.
Pure chemicals may also be used as olfactory attrac- tants in this assay. Ethyl acetate at a series of concen- trations was added to agarose and was found to attract
TABLE 2
Isolation of trap assay mutants
lines No. Mutants
Screen Mutagen Enrichment Stimulus screened obtained
I EMS
-
Medium 413 otalI 1 X-rays
+
Medium 34 ota2I11 Hybrid
+
Medium 193IV EMS
-
Ethyl acetate 500 otajr, 4 , 5 , 7Drosophila in a dose-dependent manner. T h e dose having the strongest effect was 0.5%, which gave a score of 5.9 f 0.2 in a screening of 500 mutagenized lines (Table 1). Other pure chemicals, including ace- tone, propionic acid, acetic acid, and ethanol also attract flies in this assay, although less effectively
u.
RIESCO-ESCOVAR andJ.
CARLSON, in preparation).Can the assay be used to detect a response to other flies, as might be expected if Drosophila secreted an attractive pheromone? [For a review of currently char- acterized Drosophila pheromones, see Jallon (1 984); see also BARTELT, SCHANER and JACKSON (1985).] Figure 2D shows that ten virgin females placed in the traps elicited a strong attractive effect on the tested males; however, trapped males also exerted an attrac- tive effect on tested males. T h e attractive effect of trapped organisms in the assay has not been examined in detail, and we know neither the nature of the stimulus nor whether it has ecological significance. These results do suggest, however, that behavior of different individuals during the course of an assay may not be independent, since the stimulus would effectively change as the traps accumulate animals. An attractive effect of trapped organisms would serve, then, to amplify the signal generated by the original chemical stimulus.
Isolation of mutants defective in the trap assay. Four screens have been performed using the trap assay (Table 2). T h e first was a screen of 41 3 lines each containing an individual
X
chromosome muta- genized with EMS, using Drosophila culture medium as the olfactory attractant. From this screen one mu- tant was recovered, otal (ota = olfactory trap abnormal;the numeral refers to the mutant and is not meant to designate a genetic locus). T w o other screens were performed on sets of lines mutagenized by means of X-irradiation or hybrid dysgenesis. As these methods generally induce mutations at low frequencies, an enrichment procedure was carried out prior to estab- lishment of these lines (see MATERIALS AND METHODS). One mutant, ota2, was isolated from among the X- irradiated lines (Figure 4A). No mutants were isolated from the dysgenic lines.
Examination of otal revealed that it was defective not only in entering traps containing medium, but also in entering traps containing ethyl acetate (Figure
A
B
0
TIME (HRS)
Control
O h 2
" Control
-c Olal
2 0 4 0 6 0 80
-c Olal
L
2 0 4 0 6 0 80
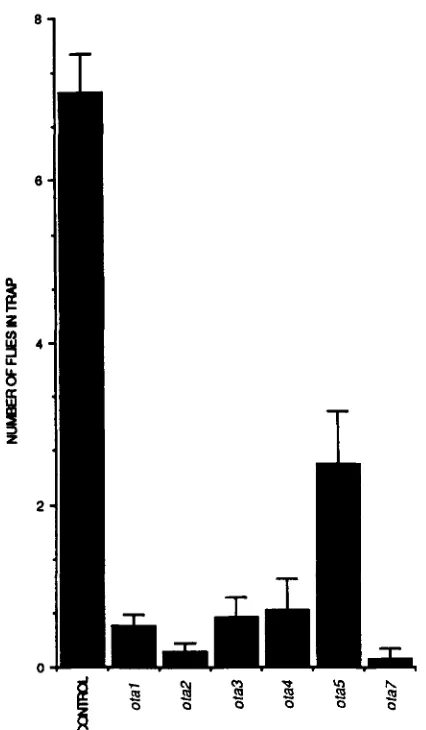
4B). Moreover, the data suggested a more severe phenotype in the case of the pure stimulus. In light of these results, a fourth set of 500 lines each containing an X chromosome mutagenized with EMS was screened using ethyl acetate as a test stimulus; these lines were independent of those used in the prior EMS screen. From this screen four new ota mutants were obtained (Figure 5 ) .
Morphological examination of otu Mutants: No cuticular abnormalities were detected among any of the six ota mutants. Special attention was paid to the antennae, which appeared normal in structure and size when examined at 128X magnification under a dissecting microscope. In no case does the reduced rate of trap entry appear to be due to an increase in body dimension.
Behavioral characterization of otu mutants: As is the case with many behavioral screens, defective re- sponse in the trap assay can result from lesions at any of a variety of levels, including sensory transduction, central processing, and motor function. One method which may be useful in distinguishing mutants with defects in olfactory transduction or processing from those whose defects affect exclusively motor or other functions, is to test them in a second chemosensory paradigm of very different design. The chemosensory jump response seems particularly appropriate: it meas- ures a different motor activity and operates on a time scale almost five orders of magnitude shorter than that of the trap assay. It will be described in detail
TIME [HRS)
FIGURE 4.-Response of otal and ota2 males in trap assay. A) Time course of ota2 entry into traps. Dro- sophila culture medium was the at- tractant. (SEM; N = 10 for ota2 and for control CS-5) E) Time course for otal entry into traps. Ethyl acetate was the stimulus. (SEM; N = 20 for
otal and for control CS-5).
elsewhere (MCKENNA et al. 1989); briefly, flies are stimulated to jump by exposure to high concentrations of any of a variety of volatile chemicals, and this response can be quantitated accurately and efficiently with the aid of a simple apparatus constructed for this purpose.
An example of a mutant which is defective in the trap assay but normal in the chemosensory jump assay is ota7. This mutant appears to behave sluggishly-at least when observed in the trap assay-and was sus- pected to contain a defect affecting functions other than olfactory transduction and processing. Figure 6
shows that ota7 jumps normally in response to ethyl acetate. Another mutant, otu2, also jumps normally to ethyl acetate. The other four mutants, however, o t a l , 3, 4 and 5 , are defective in jump response to ethyl acetate, a stimulus to which they also respond defec- tively in the trap assay.
Genetics of Drosophila Olfaction 321
8
IT
FIGURE 5.-Trap assay phenotypes of otu mutant males. Ethyl acetate was the stimulus in all cases except for ota2, in which culture medium was used. (SEM; 17 5 N 5 20 for otu mutants, N = 77 for
control CS-5).
Of the two mutants which jump normally to ethyl acetate, both jump normally in response to propionic acid and to benzaldehyde as well (Figure 6). The four
otu mutants which are defective in jump response to ethyl acetate, however, divide into two classes. One class, consisting of a single mutant, otul, is also defec- tive in response to propionic acid and benzaldehyde. The second class, comprising otu3, 4 and 5 , is also defective in response to propionic acid, but responds normally to the tested concentration of benzaldehyde. Physiological defects in the otu mutants: As a third approach to characterizing the mutants, we have ex- tended the analysis from the behavioral level to the physiological level. How many of the ota mutants have electrophysiological defects? Two types of electro- physiological recording have been used to investigate the nature and scope of the otu defects. First, electro- retinograms (ERGS) were recorded because of their sensitivity as a means of detecting physiological ab- normalities and because of their convenience; they also address the question of whether the otu pheno-
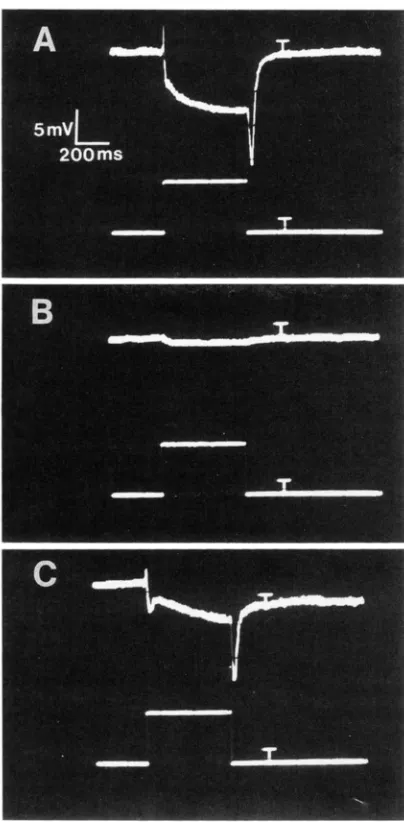
types extend to include another sensory modality, the visual system.
Two of the otu mutants exhibit abnormalities in visual system physiology (Figure
7).
otul exhibited defective retinal depolarization in all of 30 individuals examined. The characteristic “on” and “off” tran- sients, which in wild-type reflect postsynaptic activity in the neurons of the lamina (GOLDSMITH and BER-NARD 1974; COOMBE 1986), are also missing in otul.
otu7 also showed an abnormal ERG waveform in 38 of
43
animals tested, with the remaining five animals appearing normal. The otu7 individuals displaying the mutant phenotype exhibit an apparently normal “on” transient, and then, as in wild-type, their recorded potential becomes negative. However, unlike wild- type controls tested in parallel under the same condi- tions, their potential then abruptly begins a return toward baseline, but then once again becomes increas- ingly negative until a normal “off” transient occurs at the end of the stimulus. This abnormality could reflect a defect either in the physiology of the photoreceptor cells or in the neurons of the lamina.The physiology of the giant fiber system was also examined (WYMAN et al. 1984). This system, which drives an escape response consisting of a jump and flight initiation in response to a visual stimulus, was tested by stimulating the brain electrically and record- ing from the tergotrochanteral muscle-an extensor muscle of the mesothoracic leg-and the dorsal-lon- gitudinal muscles, which are wing-depressor muscles used in flight. All six mutants performed normally, suggesting normal propagation of action potentials through at least four defined types of neurons, normal transmission at five synapses-both electrical and chemical-and normal electrical response of two mus- cle types. The bendless mutant of THOMAS AND WY-
MAN (1984) was used as a negative control and was
clearly distinguished as abnormal.
The phenotypes of the otu mutants are summarized in Table 3.
Genetic characterization of the otu mutants: In order to determine whether the trap assay might be a convenient tool for genetic analysis of X-linked olfac- tory mutations, it was necessary to determine whether females would respond in the assay. We found that when the dimensions of the trap were enlarged slightly
so as to accommodate their larger size, Drosophila females gave a strong response. Moreover, homozy- gous females of all the otu lines were found to give significantly lower scores than wild-type ( P
<
0.05)except otu5/otu5, which, interestingly, behaved nor- mally and whose genetics are not considered further here.
T
FIGURE 6.-Chemosensoryjump phenotypes of ota
mutants. A detailed description of the jump assay will appear elsewhere (MCKENNA et ul. 1989). Briefly, a fly is placed in a vertically oriented plastic t u b e and allowed to crawl halfway up the side. An airstream is
Ethyl Acetate pulled through the tube-which has a small hole at the top-at a rate of 1 liter/min. The introduction of an odorant into the airstream induces the fly to jump. A positive response is scored if the fly jumps off the wall of the tube and lands at the bottom of the tube Benzaldehyde within 3 sec after introduction of the odorant. Flies were tested once and then discarded. Mutants were tested with ethyl acetate and propionic acid, both used undiluted, and ben~aldehyde, diluted 1 : l O in paraffin oil. Error bars represent SEM; N = 5 assays for all mutants except N = 1 for otu2 and o l d . Each assay consisted of trials of 30 male flies (50 flies in the case of otal). See Table 3 for statistical significance.
Propionic Acid
also have allowed isolation of dominant mutations, whether X-linked or autosomal. As a first step in characterizing the genetic basis of the ota defects, we sought to determine whether they were X-linked. Testing of attached-X females from the otal, 2, 3, 4
and 7 attached-X lines showed that none gave scores significantly lower than that of control attached-X females. These are the results that would be expected
if all these ota lesions were X-linked.
Testing of ota/CS-5 heterozygotes showed that the olfactory phenotypes of otal, 2 and 4 were recessive, but that those of ota3 and ota7 were at least partially dominant (ANOVA analysis of arcsine transformed trap assay scores revealed ota3/CS-5 and ota7/CS-5 to be significantly lower than CS-5/CS-5 at the P
<
0.01 and P<
0.05 levels, respectively); they will therefore be referred to henceforth as Otu3 and Ota7. The ERG phenotypes of both otal and Ota7 were found to be recessive: all of 10 otal/CS-5 heterozygotes and all of15 Ota7/CS-5 heterozygotes had normal ERGS. Complementation analysis based on all pairwise combinations of ota mutants (Figure 8) showed that the recessive mutants define two complementation groups, one comprising otal and the other comprising both ota2 and ota4. (ota2/ota4 heterozygotes gave a
score of 3.6 f. 0.9; N = 27, compared to the CS-5 value of 8.0 f 0.5; N = 27, and are thus significantly different from wild type at the P
<
0.001 level). In light of the failure of ota2 and ota4 to complement,otu4 will henceforth be referred to as 0tu2~. All Ota3 and all Ota7 heterozygotes gave trap assay scores significantly lower than wild type ( P 0.05 in all cases), consistent with our determination that both are dom- inant.
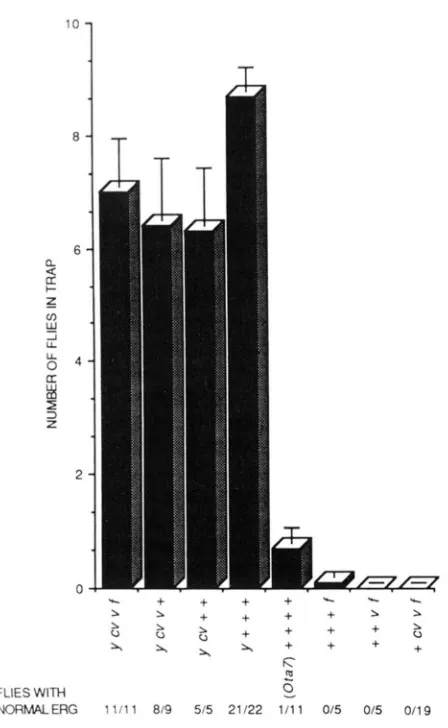
We attempted to map otal, 2, 3 and 7, in part to address the question of whether their olfactory phe- notypes arise in each case as a consequence of single mutations, but also to investigate whether the ERG phenotypes of otal and 7 cosegregate with the olfac- tory phenotypes. A stock carrying the multiply marked X chromosome y CD v f , which gives a strong response in the olfactory assay, was used for recom- binational mapping of Ota7. Flies representing each class of recombinants were tested in groups of ten for olfactory behavior, and the results (Figure 9) indicate that this phenotype maps to the region of the
X
chromosome distal to cv. The fact that the classes y+++ and
+CD
v f show phenotypes like those of the parental chromosomes y CD vfand++++,
respectively,Genetics of Drosophila Olfaction 323
TABLE 3
FIGURE 7.-Electroretinogr;1nl (ERG) analysis of ota mutants. The light stimulus is of 0.5-sec duration. The small “T” visible on the traces approximately 200 msec after the end of the flash is an artifact of the triggering system. A) CS-5. A normal “on” transient, retinal depolarization (typically 5-15 mV), “off“ transient, and return to resting potential are visible; B) otal; and C) o l d .
consistent with a map position near y, although this interpretation is complicated by the use of a popula- tion assay in which flies may not behave independ- ently.
A map position near y was also determined for the ERG phenotype of Ota7 (Figure 9). Among 41 recom- binant chromosomes with breaks in the y-co interval, in only one case did the ERG phenotype fail to seg- regate with y. More precise mapping of both pheno- types, and further evidence for cosegregation of the olfactory and ERG phenotypes, was obtained in a second experiment, in which another 20 y+++ males and another 20
+co
ufmales were mated to attached-X females to establish lines. Both olfactory and ERG phenotypes were tested among the F1 male progeny, which should contain the same recombinant X chro-
Summary of trap assay mutant phenotypes
Jump Phenotypeb
Mutant Trap score’ EtAc BT PA ERG‘
otal 0.5 f 0.1 (20)
-
-
ota2 0.2 f 0.1 (17)
+
+
+
+
otajr 0.6 f 0.2 (18)
-
+
-
+
ota4 0.7 f 0.4 (19)
-
+
-
+
ota5 2.5 f 0.7 (19)
-
+
-
+
ota7 0.1 f 0.1 (18)
+
+
+
-
CS-5 6.9 f 0.3 (77)-
-
a Mean f SEM (no. of trials). All flies tested were male. The stimulus was 0.5% ethyl acetate.
Indicates different from control CS-5, P < 0.05;
“+”
indi- cates P > 0.05. ANOVA or Student’s t-test was used after arcsine transformation of the jumping percentages shown in Figure 6. otal and CS-5 were tested in one experiment and the results wvere compared with Student’s t-test. The other ota mutants and CS-5 were tested in a separate experiment and the results were compared by ANOVA.b
.._.I
.I
.._..
indicates distinguishable from the control CS-5.
cs-5 ofa I ofa2 Ora3 ofa 4 ota 7
cs-5
ofa I
ofa 2
ota 3
ora 4
@a 7
+
+
+
-
+
-
FIGURE 8.-Complementation analysis of ofa mutants. “-” indi-
cates that response differs from that of wild type at the P < 0.05 level.
”+”
indicates complementation, i e . , response not significantly different from that of wild type. CS-5 scored 8.0 f 0.5 ( N = 27) in this experiment, and all scores represented by the ”+” designation ranged between 6.0 & 0.9 and 8.0 f 0.5. Mean scores represented by the ”-” designation were 1 4 . 4 in all cases, except that Ota7,which w e classify as partially dominant, gave scores of 4.8 f 1 .O ( N = 27). 5.0 f 1 . 1 ( N = 27), and 5.7 f 1.0 ( N = 24), when in heterozygous combination with ota2, otal, and CS-5, respectively.
N ranged from 15 to 27 for all of the genotypes shown in the matrix.
mosomes as their fathers. All 20
+co
uf
lines had abnormal ERGs, and all 20 y+++ lines had normal ERGs. Testing of olfactory behavior gave two non- overlapping distributions of trap assay scores: all the+co
uflines gave a mean score of 0.0 ( n = 3 trials perline), whereas all the y+++ lines gave scores greater than 0. Following retesting of those y+++ lines on the lower end of the distribution, all 20y+++ lines yielded average scores greater than 4.2 (3 I n 5 6). In summary, the olfactory and ERG phenotypes of Otu7
cosegregate; both map near y.
Since a number of neurological mutants reside very near they locus (JIMENEZ AND CAMPOS-ORTEGA 1987),
324
10
8
6
n
a
z w
e
v)-I
LL
LL
0 4
z
3
z
2
0
FLIES WITH
NORMAL ERG
C.
T
11/11 8/9 515 21/22 1/11 015 015 0119
FIGURE $?.-Mapping of Ota7 olfactory and visual system phe- notypes. Ethyl acetate was the olfactory stimulus. (SEM; N = 10 trap
assays for each genotype.)
covers y and extends from the distal
tip
of theX
chromosome to approximately 1B14 on the cytoge- netic map. Since all of ten Ota7/y2Y611 individuals tested had abnormal ERGS-although all of ten sib- ling CS-5/y2Y61Z individuals tested had normal
ERGS-we conclude that the ERG phenotype does not map to this cytogenetic interval, but rather maps proximal to it within the y-n' interval.
otal was localized to the v-finterval first by recom- binational mapping of its ERG phenotype with a y n' v
f
car chromosome. All recombinant classes could be unambiguously designated as normal or abnormal except for y co v++ and+++f
car. Testing of 100 recombinants of these classes allowed us to map the ERG phenotype approximately to position 43 within the v-f interval. T h e duplication D p ( l ; f l L . 9 , which covers the cytogenetic region 12A6-10 to 13A2-5 within this interval, was found to cover the otal ERG abnormality: all of eighty co otal animals carrying the duplication had normal ERGs, whereas all of six sib- ling y co otal animals lacking the duplication had abnormal ERGs. T h e same duplication was also found to cover the olfactory phenotype:y
co otal animalscarrying the duplication had significantly higher trap assay scores than their y co otal siblings without the duplication ( P
<
0.05). Thus both olfactory and ERG phenotypes of otal cosegregate to a small region within the v-finterval of the X chromosome.ota2 was mapped to the distal portion of the
X
chromosome using they n' vfchromosome by testing flies of each recombinant class in the trap assay. As with Ota7, the results clearly indicated a map position distal to n' and again suggested a position near y; again the olfactory phenotype was not covered by the du- plication y2Y611. T h e same method of meiotic recom- bination mapping using the same multiply marked chromosome failed, however, to indicate a map posi- tion for Ota3, whose genetic origin may be more complex than a single X-linked mutation.
DISCUSSION
This article is concerned with a new means of ex- tending genetic analysis, which has proven remarkably useful in investigating the visual system of Drosophila, to a relatively unstudied sensory system, the Drosoph- ila olfactory system. A convenient means of measuring olfactory behavior is described, characterized, and used as a screen to isolate six ota mutants. T h e new mutants are characterized phenotypically with respect to several criteria, including behavior in a second chemosensory paradigm, chemical specificity, and the physiology of two other systems, including the visual system. Genetic characterization of ota mutants is also provided, demonstrating that the method developed here can be used as a tool for genetic analysis.
T h e results of our mutant analysis show clearly that the screen described here is capable of isolating mu- tants with sensory defects: two of the mutants are shown to have defects in visual system physiology, of which one shows a severe defect in retinal depolari- zation. This finding suggests the possibility that a number of genes may be used in both visual and olfactory function, a possibility we are currently in- vestigating by further analysis of these two ota mutants and by testing mutants with defined visual system abnormalities for olfactory function.
Genetics of Drosophila Olfaction 325
olfactory system, other than receptor genes, also play roles in other systems.
T h e possibility of pleiotropism complicates the interpretation of certain ota mutant phenotypes. For example, one of the mutants suspected to affect func- tions other than sensory transduction or processing is
Ota7, which shows reduced motor activity in the trap assay. A simple interpretation of the Otu7 trap assay phenotype is that its attraction response is blocked by a defect at the motor level. However, further charac- terization is required to exclude other models, such as the possibility that sluggish activity is itself a con- sequence of diminished perception of those sensory stimuli which drive normal motor activity. Consistent with this possibility, Ota7 is, in fact, one of the two mutants exhibiting defective visual system physiology. Clearly, other criteria are required to distinguish mutants with defects at the sensory and processing levels from those with defects which act exclusively at other levels. One strategy is to test response to the same stimulus in another behavioral paradigm which makes very different demands on other levels of func- tion, such as motor activity. According to this ration- ale, those mutants containing putative sensory or proc- essing defects are likely to fail both tests. Following this approach, we have shown that four of the six ota
mutants also exhibit defects in the chemosensory jump assay.
Two caveats limit the diagnostic power of this strat- egy. First, the power of the approach is limited by the extent of overlap in the motor activities required: mutants defective in any motor skills required by both assays will fail both assays. Second, a mutant with a bona fide sensory defect may perform normally in one assay and abnormally in the other if the sensory or processing tasks presented by the two assays differ. Virtually nothing is yet known about the molecular or cellular basis of the sensory perception driving either the trap or the jump assays, and we do not know that they act through the same transduction mechanism.
A second, converse, strategy to distinguish those otu
mutants with sensory defects is to retain the paradigm but to alter the stimulus. In principle, the stimulus can be altered at the level of the sensory modality: ota
mutants which can be shown to enter traps normally in response to a visual stimulus, for example, may contain lesions in olfactory transduction or processing. A finer and potentially more informative alteration is at the level of the chemical: mutations affecting re- sponse to one chemical and not to another are likely to be defective in olfactory transduction or process- ing-some could even identify receptor genes. We have investigated the chemical specificity of the ota
mutants using the chemosensory jump assay, on ac- count of its high signal-to-noise ratio for a variety of
chemicals (MCKENNA et al. 1989). Evidence presented here identifies 0 t a 2 ~ , 3 and 5 as candidates for “chem- ically specific” mutants. While a detailed description of these behavioral defects, including analysis of re- sponse to a range of chemical concentrations, is re- quired before rigorous conclusions can be drawn re- garding specificity, chemically specific mutations have been described previously by RODRICUES (1980) and by HELFAND and CARLSON (1 989).
We note that although otu2 and otu2’ appear to be allelic, the data indicate a chemosensory jump phe- notype for ota2’ but not for ota2. There are several possible explanations for this difference. It is possible that otu2’ is a more severe allele than ota2; however, this possibility does not receive support from their respective trap assay scores, both of which are very low. Alternatively, the two alleles may encode quali- tatively different forms of gene product which affect different functions to differing extents. An additional possibility is raised by the fact that the jump phenotype of ota2’ has not been mapped: we can not exclude the possibility that it arises at least in part from a second mutation at another locus.
A third strategy for distinguishing mutants with sensory defects dispenses altogether with certain other levels of function, including the motor level. Physio- logical recording from the visual system of the otu
mutants has already been informative. A direct test for defects in olfactory transduction is planned by means of physiological recording from the antenna (VENARD and PICHON 1981; BORST 1984; SIDDIQI
1984).
In the long term, genes required specifically for response to a subset of chemicals may be particularly useful in analyzing the functional organization of the olfactory system. Molecular probes derived from such genes may allow relationships to be drawn between structure and function. It would be of interest to determine, for example, (1) whether a gene required for response to ethyl acetate but not benzaldehyde is expressed only in a particular subset of antennal sen- silla, i.e., a subset defined either on the basis of sensil- lum morphology or position on the third antennal segment;
(2)
the number of neurons within a sensillum in which the gene is expressed; (3) whether neurons in which the gene is expressed project to certain subsets of the -25 glomeruli which constitute each antennal lobe of the brain (STOCKER et al. 1983). Such genes might also be useful in addressing questions concerning olfactory system development, such as the question of when functional pathways become differ- entiated.LITERATURE CITED
AYER, R., P. MONTE and J. CARLSON, 1989 The isolation of antennal mutants and their use in Drosophila olfactory genetics, in Neurobiology of Sensory Systems, edited by R. SINGH and N. STRAUSFELD. Plenum Press, New York (in press).
BARROWS, W., 1907 The reactions of the Pomace Fly, Drosophila Ampelophila Loew, to odorous substances. J. Exp. Zool. 4: 515- 537.
BARTELT, R.,A. SCHANER, A N D L. JACKSON, 1985 cis-Vaccenyl acetate as an aggregation pheromone in Drosophila melano- gaster. J. Chem. Ecol. 11: 1747-1756.
BENZER, S., 1967 Behavioral mutants of Drosophila isolated by countercurrent distribution. Proc. Natl. Acad. Sci. USA 58:
BORST, A,, 1984 ldentification of different chemoreceptors by electroantennogram-recording. J. Insect Physiol. 3 0 507-510. COOMBE, P. E., 1986 The large monopolar cells L1 and L2 are
responsible for ERG transients in Drosophila. J. Comp. Physiol.
CRAYMER, L., and E. ROY 1980 Drosophila Inform. Serv. 55:
DOANE, W., 1967 Drosophila, pp. 219-244 in Methods in Devel- opmental Biology, edited by F. WILT and N. WESSELLS. Thomas F. Crowell, New York.
ENGELS, W., 1979 Extrachromosomal control of mutability in Drosophila melanogaster. Proc. Natl. Acad. Sci. U S A 7 6 401 1- 40 15.
GOLDSMITH, T., and G. BERNARD, 1974 The visual system of insects, pp. 165-272 in The Physiology oflnsecta, edited by M. ROCKSTEIN. Academic Press, New York.
HARDY, R., D. LINDSLEY, K. LIVAK, B. LEWIS, A. SILVERSTEN, G.
JOSLYN, J. EDWARDS and S. BONACCORSI, 1984 Cytogenetic analysis of a segment of the Y chromosome of Drosophila melanogaster. Genetics 107: 59 1-6 10.
HELFAND, S., and J. CARLSON, 1989 Isolation and characterization of an olfactory mutant in Drosophila with a chemically specific defect. Proc. Natl. Acad. Sci. USA 86: 2908-2912.
HUBEL, D., and T . WIESEL, 1977 Functional architecture of ma- caque monkey visual cortex. Proc. R. SOC. Lond. Biol. 198: 1- 59.
JALLON, J., 1984 A few chemical words exchanged by Drosophila during courtship and mating. Behav. Genet. 1 4 441-477.
JIMENEZ, F., and J. CAMPOS-ORTEGA, 1987 Genes in subdivision
1B of the Drosophila melanogaster X-chromosome and their influence on neural development. J. Neurogenet. 4: 179-200. KULKARNI, S., A. STEINLAUF, and J. HALL, 1988 The dissonance
mutant of courtship song in Drosophila melanogaster: isolation, behavior and cytogenetics. Genetics 118: 267-285.
LEWIS, E., and F. BACHER, 1968 Method of feeding ethyl methane 1 112-1 119.
A 159 655-665.
200-204.
sulfonate (EMS) to Drosophila males. Drosophila Inform. Serv. 43: 193.
MCKENNA, M., P. MONTE, S. HELFAND, C. WOODARD and J. CARL-
SON, 1989 A novel chemosensory response in Drosophila and the isolation of acj mutants in which it is affected. Proc. Natl. Acad. Sci. USA (in press).
MONTE, P., C. WOODARD, R. AYER, M. LILLY, H. SUN and J. CARLSON, 1989 Characterization of the larval olfactory re- sponse in Drosophila and its genetic basis. Behav. Genet. 19:
PAK, W., and S. GRABOWSKI, 1978 Physiology of the visual and flight systems, pp. 703-733 in The Genetics and Biology of Drosophila, Vol. 2a, edited by M. ASHBURNER and T . WRIGHT. Academic Press, London.
RODRIGUES, V., 1980 Olfactory behavior of Drosophila melano- gaster, pp. 361-371 in Development and Neurobiology of Drosoph- ila, edited by 0. SIDDIQI, P. BABU, L. HALL and J. HALL. Plenum Press, New York.
RODRIGUES, V., and 0. SIDDIQI, 1978 Genetic analysis of chemo- sensory pathway. Proc. Ind. Acad. Sci. 8 7 B 147-160. SHEPARD, G., 1983 Neurobiology. Oxford University Press, New
York.
SIDDIQI, O., 1984 Olfactory neurogenetics of Drosophila, in “Ge- netics: New Frontiers,” Proceedings of the XV International Congress of Genetics, Symposium on Neurogenetics Unipub, Ann Arbor.
SIDDIQI, O., 1987 Neurogenetics of olfaction in Drosophila mela- nogaster. Trends Genet. 3: 137-142.
STOCKER, R., R. SINGH, M. SCHORDERET, and 0. SIDDIQI, 1983 Projection patterns of different types of antennal sen- silla in the antennal glomeruli of D. melanogaster. Cell Tissue Res. 232: 237-248.
TANOUYE, M., and R. WYMAN, 1980 Motor outputs of giant nerve fiber in Drosophila. J. Neurophysiol. 44: 405-42 1.
THOMAS, J., and R. WYMAN, 1984 Mutations altering synaptic connectivity between identified neurons in Drosophila. J. Neu- rosci. 4 530-538.
VENARD, R., and Y. PICHON, 1981 Etude electro-antennograp- hique de la reponse peripherique de I’antenne de Drosophila melanogaster a des stimulations odorantes. C. R. Acad. Sci. Paris 293: 839-842.
VENKATESH, S., and R. SINGH, 1984 Sensilla on the third antennal segment of Drosophila melanogaster Meigen. Int. J. Insect Mor- phol. Embryol. 13: 51-63.
WYMAN, R., J. THOMAS, L. SALKOFF and D. KING, 1984 The Drosophila giant fiber system, pp. 133-161 in Neural Mecha- nisms of Startle Behavior, edited by R. EATON. Plenum Press, New York.
267-283.