Role of a-Factor and the MFal a-Factor Precursor in Mating in Yeast
Shari Caplan’ and Janet Kurjan2
Department of Biological Sciences, Columbia University, New York, New York 10027 Manuscript received July 12, 1990
Accepted for publication October 18, 1990
ABSTRACT
The peptide pheromones secreted by a and a cells (called a-factor and a-factor, respectively) are each encoded by two structural genes. For strains of either mating type, addition of exogenous pheromone does not alleviate the mating defect of mutants with disruptions of both structural genes. I n addition, a particular insertion mutation in the major a-factor structural gene ( M F a l ) that should result in an altered product inhibits a mating. These results suggested that the pheromone precursors (the M F a l pro region in particular) might play a second role in mating separate from the role of
pheromone production. T o analyze the role of a-factor and the M F a l precursor in a mating, we have constructed two classes of mutants. The mating defects of mutants that should produce the M F a l pro region peptide but no a-factor could not be alleviated by addition of exogenous a-factor in crosses to a wild-type a strain, indicating that the previous results were not due to an inability of the disruption mutants to produce the pro region peptide. Mutants able to produce a-factor, but with a variety of
alterations in M F a l precursor structure, mated at levels proportional to the levels of a-factor produced, suggesting that the only role of the a-factor precursor in mating is to produce a-factor. Both of these results argue against a role for the M F a l pro region separate from its role i n a-factor production. We also describe results that show that in vivo production of a-factor’ (the form of a-
factor encoded by one of the two a-factor repeats of M F a 2 ) is equivalent to the major form of a- factor for induction of all responses necessary for mating. We discuss the implications of these results on the role of the pheromones in mating.
M
ATING in the yeast Saccharomyces cerevisiae in- volves secretion of and response to peptide pheromones (reviewed by CROSS et al. 1988). a Cells secrete a-factor and respond to a-factor, and a cells secrete a-factor and respond to a-factor. Response to pheromone elicits arrest in the G I phase of the cell cycle, cell wall changes, morphological alterations, and induction of genes that encode products involved in aspects of mating or pheromone response. Mutants defective in pheromone production or response are sterile (HARTWELL 1980; KURJAN 1985; MACKAY and MANNEY 1974; MANNEY and WOODS 1976; MICHAE- LIS a n d HERSKOWITZ 1988).a-Factor is a 13-amino acid peptide that is encoded
by two unlinked structural genes, M F a l a n d M F a 2 .
These genes encode precursors of 165 and 120 amino acids, respectively (KURJAN a n d HERSKOWITZ 1982;
SINCH et a l . 1983). Both precursors include a signal sequence, a pro region with three sites for N-linked glycosylation, and tandem a-factor repeats each of which is preceded by a spacer peptide (Figure 1).
M F a l contains four spacer-a-factor repeats, all of
which encode a peptide identical to the a-factor se- quence determined by analysis of secreted a-factor
School of Medicine, New Haven, Connecticut 06510. CJ~liversit! of Vermont. Burlington, Vermont 0540.5. ( ~ c w t i c s 127: 299-307 (February, 1991)
’
(:urrcnt ;~ddt-ess: Department of Human Genetics, Yale University’ (;urrent address: I k p a r t m e n t of Microbiology arid Molecular Genetics,
(STOTZLER, KILTZ a n d DUNTZE 1976; TANAKA et al.
1977). M F a 2 contains two spacer-a-factor repeats; one repeat encodes a-factor (Gln-5,Lys-7 a-factor) and one repeat encodes a peptide that differs from a-factor by
two amino acid substitutions (Asn-5,Arg7 a-factor; to
be referred to as a-factor’). T h e M F a l precursor is processed in several well characterized steps to pro- duce mature a-factor (reviewed by FULLER, STERNE and THORNER 1988).
M F a l is responsible for the majority of a-factor produced (KURJAN 1985), providing an explanation for the fact that amino acid sequencing of secreted a- factor identified only a-factor and not a-factor’ (STOT- ZLER, KILTZ a n d DUNTZE 1976; TANAKA et al. 1977). Analysis of synthetic a-factor’ indicated that this pep- tide is equivalent to a-factor in the induction of several aspects of pheromone response, including cell cycle arrest, shmoo formation and induction of agglutina- tion (KURJAN a n d LIPKE 1986; RATHS et al. 1986).
300 S. Caplan and J. Kurjan
TABLE 1
Yeast strains
Strain Genotype Source
W303-1B“ MATa ade2-1 canl-100 R. ROTHSTEIN
leu2-3,112 his3-11,15 trpl-1 ura3-1
RC618 MATa ade2-1 canl cyh2 his6 R. CHAN
metl ural rme
RC629 MATa sstl-2 ade2-1 canl R. CHAN
cyh2 his6 metl u r a l rme
RC63 1 MATa sst2-1 ade2-1 canl R. CHAN
cyh2 his6 metl u r a l rme
227 MATa cry1 lysl I . HERSKOWITZ
” All mfa mutant constructs are isogenic to this strain. exogenous a-factor does not alleviate the mating de- fect of an a mfal mfa2 strain (MICHAELIS and HER-
SKOWITZ 1988). One possible explanation for these
results is that the M F a and MFA precursors play multiple roles in mating, a role in pheromone produc- tion and a separate uncharacterized role. For the M F a genes, a second result seems consistent with this hy- pothesis. One particular m f a l mutant (mfal::URA3A) shows an unexpected phenotype, a 1 0-fold lower level of mating than an m f a l null mutant containing a deletion of the entire M F a l gene (KURJAN 1985). T h e mfal::URA3A mutant encodes a hybrid protein con- sisting of the M F a l signal sequence, the pro region, and the first spacer sequence, followed by several amino acids encoded by the U R A 3 fragment (Figure
1). The phenotype of the mfal::URA3A mutant indi- cates that this hybrid gene product inhibits a mating, suggesting that mfal::URA3A is a change-of-function mutation, and therefore that the wild-type M F a l pro region might play a positive role in a mating.
We have constructed new m f a l mutations and tested their effect on a-factor production and mating to further elucidate the possible role of the precursor in mating separate from its role in a-factor produc- tion. Two complementary approaches were taken. T h e first approach involved constructing mutants that should allow production of the pro region but no a- factor and testing for alleviation of the resulting mat- ing defect by addition of exogenous a-factor. T h e second approach involved an attempt to identify pro region mutants that allowed a-factor production but resulted in a mating defect. T h e results do not support the hypothesis that the M F a I precursor plays a role in mating separate from its role in a-factor produc- tion. We therefore favor an alternative hypothesis, that secretion of an appropriate concentration or gra- dient of pheromone in the vicinity of a mating partner is critical for efficient mating.
MATERIALS AND METHODS
Strains, plasmids and media: Yeast strains are listed in Table 1. All of the mutants were constructed in
strain W303-1B. Mutants mfal::URA3A, mfaI::LEUBC, mfa2::URA3 and mfa2::LEU2C, also constructed in this strain, were described previously (KURJAN 1985). The mfal::LEU2C, mfa2::URAjr and mfa2::LEU2C mutations are deletions of most or all of the MFal and MFa2 coding regions and therefore are null mutations. Escherichia coli strain RRI was used for all plasmid constructions and 7 1.18 for M 13 cloning and mutagenesis.
Plasmids pHK2 and pBH3 contain one and two copies of the 1.7 kb MFal EcoRI fragment, respectively, and YIp30 contains the URA3 gene. pUC18::LEU2 (constructed by J. HILL) is pUCl8 with the LEU2 BglII fragment inserted into the BamHI site of pUC18. YEp6 is a high-copy-number plasmid containing the HIS3 gene. pJKl contains the 1.7- kb MFal EcoRI fragment subcloned into the EcoRI site of pIH3 (constructed by I. HERSKOWITZ), which is YIp5 with the HindIII site eliminated.
Media were as described previously (KURJAN 1985). Construction of altered MFaZ genes: Elsewhere, we de- scribe the construction of several types of structural altera- tions of the M F a l gene, with which gene replacements were made and used to test effects on a-factor secretions and precursor build-up (CAPLAN et al. 1991). These constructs include MFal genes that differ with respect to the number of spacer-a-factor repeats; the mfal-al, mfal-a2 and
mfal-a3 genes contain one, two and three tandem spacer- a-factor repeats, respectively. Other alterations include mu- tation of N-glycosylation sites, and in-frame deletions and insertions in the pro region.
The 1.5-kb mfal-a1 EcoRI fragment was subcloned into M 13mplO for use as the template for site-directed mutagen- esis (ZOLLER and SMITH 1983). T h e oligonucleotide, 5’-
GCTTGGCATTGGTTGAACTTAASACCCG-3’ (nu-
cleotide changes from the wild-type sequence are under- lined), which alters the single a-factor sequence to an a- factor’ sequence, was synthesized on an Applied Biosystems model 380A DNA synthesizer by the phosphoramidite method and used to produce mfal-al‘. The presence of the nucleotide changes were confirmed by dideoxy sequencing (SANGER, NICKLEN and COULSON 1977) using the oligonu- cleotide 5’-TTTATAAATACTGCTATT-3’, which primes within the MFal pro region.
The mfal-pro3 gene was constructed by insertion of BamHI linkers at AluI sites at amino acids 19 and 37 and deletion of the small fragment between the linkers (as de- scribed for the construction of mfal-pro2; CAPLAN et al. 1991). This construction results in an in-frame deletion of 17 amino acids and an insertion of three amino acids at the beginning of the pro region.
The MFal gene contains four closely spacedHindIII sites, separated by 63 bp, at each of the spacer-a-factor boundaries (KURJAN and HERSKOWITZ 1982; KURJAN 1985; see Figure
and to determine the number of 63 bp a-factor-spacer HindIII fragments present at the ends of the inserted frag- ment.
Gene replacements: Plasmids or M 13 DNAs containing mfa constructs that did not contain a selectable marker were cleaved with EcoRI and the EcoRI fragments were cotrans- formed along with YEp6 [HIS?] into a MATa mfal::SUP4 mfa2::LEUZ canl-I00 strain (CAPLAN et al. 1991). SUP4 is able to suppress the canl-100 ochre mutation, resulting in canavanine sensitivity; therefore, Hisf transformants were plated on canavanine plates to select for canavanine-resistant transformants in which the mfal::SUP4 gene had been re- placed by the new construct (ROTHSTEIN 1983). Replace- ments were confirmed by Southern analysis. Gene replace- ments with mfal constructs containing selectable markers were made using the one step gene disruption technique (ROTHSTEIN 1983) and were confirmed by Southern analy- sis.
a-Factor secretion, mating and a-factor addition assays:
Levels of a-factor secreted were determined by halo assays. More quantitative estimates were obtained by making serial dilutions of cells and spotting on supersensitive lawns of a
cells as described previously (KURJAN 1985). Mating was assayed by patch tests in which MATa strains to be tested were replicated to minimal media on which a lawn of MATa
227 tester cells (Table 1) had been spread and the presence of prototrophs scored. Quantitative efficiency-of-mating ex- periments were done as described by DUTCHER and HART-
WELL (1982). To test alleviation of the mating defects by addition of exogenous pheromone, a-factor (Sigma) was spread on buffered minimal plates with adenine (KURJAN
1985), and mating patch tests were done as described above with the tester lawns RC6 18, RC629 and RC63 1 (Table 1).
DNA sequencing: T h e sequences of the mfal hybrid carboxy termini were determined by the Sanger dideoxy terminator method. A 17-nucleotide oligomer (5’-
TGCCATTTTCCAACAGC-3’), complementary to a re- gion within the MFal pro region, was synthesized using an Applied Biosystems DNA synthesizer and used to sequence the mutant plasmids (SANGER, NICKLEN and COULSON 1977).
RESULTS
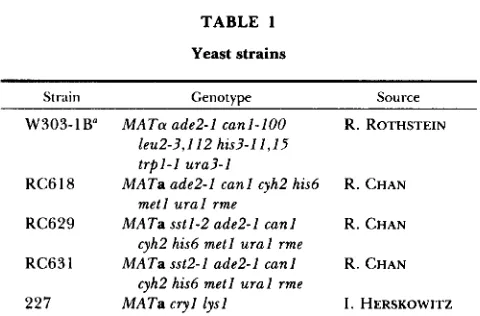
Inhibition of mating is specific to the mfal::URA3A carboxy terminus: T h e mfal::URA3A gene should encode a hybrid product with the carboxy terminus encoded by one end of the URA3 fragment (KURJAN 1985; Figure 1, A and C). T h e inhibition of mating by the mfal::URA3A product could be specific to the particular carboxy terminus of this hybrid or could result from any abnormal carboxy terminus at this position. A new set of disruptions were made in which different sequences were inserted at the same position (Figure 1). T h e mfal::URA3D construction has the same URA3 fragment inserted into M F a l as mfal::URA3A, but in the opposite orientation. T h e carboxy termini of the mfal::URA3A and mfal::URA3D hybrids are 20 and 30 amino acids, respectively, encoded by opposite ends of the URA3 fragment (Figure 1 C). T h e mfal::LEU2E construction has a LEU2-containing fragment inserted at the same position. T h e carboxy terminus of the mfal::LEU2E hybrid is seven amino acids encoded in the multiple cloning site at the end of the inserted LEU2 fragment.
A
MFal L l l l ,
MF a2 h x x x r
mfal -a3 k,,,
mfal-a2 V , , ”
mfal -a1 P , ~ A
-
mfal-al’ V / / A I,:-
mfal ::URA3A k , , 1
-
mfal ::URA3D G z I n
mfa1::LEUZE P
mfal::URA3C ” ” ’ -.__I
mfal::URA3E V ’ r n
mfal ::LEUZF o I I I
,.
B
Pheromonea-factor W H W L Q L K P G Q P M Y a - f a c t o r ’ WHWLNLRPGQPMY
c
Carboxy termini of hybridsm f a l : : U R A 3 A , mfal: :URA3C F Q F I I F F L F F F L I S L S L K F F
mfnl::URA3D, m f a l : : U R A 3 E FSFQFFFFRHYKNHYDRDSRVITDIIKLKL
m f a l : :LEU2E, m f a l : :LEUZF C M P A G R L
FIGURE 1.-MFal constructs. A) The MFal and MFa2 precur- sors (KUR,JAN 1985; SINCH et al. 1983) are diagrammed at the top. T h e signal sequences are indicated by diagonal lines, the pro region (within which are three sites for N-glycosylation) by open rectangles (or a lightly dotted rectangle for M F a Z ) , spacer sequences by spotted rectangles (or zig-rag rectangles for MFa2), a-factor se- quences by black rectangles, and a-factor’ sequences by black rectangles with dots. There is a Hind111 site at each of the spacer a-factor boundaries in M F a l . Constructs with reduced numbers of spacer-a-factor repeats include mjal-al, mfal-a2, and mfal-a3, which were constructed by deleting the small Hind111 fragments consisting of a-factor-spacer sequences. T h e a-factor sequence in
mfaul-a1 was mutated to the MFa2-encoded a-factor’ sequence to produce mfal-al‘. T h e remaining mutants were constructed by inset-tion of a URA3 fragment in either orientation or a LEU2 fragment into the HindIII sites after either the first or second spacer. T h e resulting disruptions should encode the M F a l sequence throughout the first or second spacer and read into the end of the inserted fragment until a termination codon is reached to produce hybrid carboxy termini. T h e mfaI::URA3A and mfal::URAjC ter- mini are indicated by rectangles with horizontal lines, the
mfaI::URA3D and mjaI::URA3F: termini by rectangles with widely spaced vertical lines, and the mjal::LEU2E and mfal::LEU2F ter- mini by rectangles with closely spaced vertical lines. B) T h e amino acid sequences of a-factor and a-factor’ are shown using the single letter code. C) T h e amino acid sequences of the hybrid carboxy termini encoded by disruption constructs diagrammed in (A) are shown.
T h e effects of these mutations were tested in strains that contain wild-type MFa2 and therefore produce some a-factor and are able to mate (KURJAN 1985). A newly constructed a mfal::URA3A mutant had the same phenotype as seen previously (KURJAN 1985), a
302 S. Caplan and J. Kurjan
TABLE 2
Mating of disruption mutants
Strain" Mating
M F a 1 1
.oo
mfa1::LEUZC 0.45
mfa::URA3A 0.02
mfaI::URA?D 0.47
mfa1::LEUZE 0.43
mfa1::URAjrC 0.42
" All disruptions were constructed in W303-1B and therefore
contain a wild-type MFa2 gene. The rnfa1::LEUZC mutation results in deletion of the entire M F a l coding region (KURJAN 1985). T h e twnaining constructs are diagrammed in Figure 1.
mfal::URA3A product inhibits a mating. T h e a mfal::URA3D and mfal::LEU2E mutants, however, have the same phenotype as the mfal::LEU2C null mutant, indicating that the inhibitory phenotype is specific to the particular carboxy terminus of
mfal::URA3A. T h e mfal::URA3A MFa2 and
mfal::LEU2C MFa2 strains produced similar levels of a-factor (KURJAN 1985), suggesting that the
mfal::URA3A product did not inhibit processing or secretion of a-factor from the MFa2 precursor. Given the very low level of a-factor produced by MFa2, however, moderate inhibition of a-factor production could have been missed. Production of a-factor from a wild-type M F a l gene is reduced in strains also ex- pressing mfal::URA3A (J. KURJAN, unpublished re- sults), suggesting that the inhibitory activity of the mfal::URA3A mutation may result from reduced ef- ficiency of a-factor processing or secretion of a wild- type MFa precursor expressed in the same cell.
The aberrant carboxy termini of hybrids can af- fect a-factor production: T h e mfaI::URA3D and mfal::LEU2E mutants potentially represent strains in which to test whether the mating defect of mutants able to produce the pro region peptide but no a-factor can be alleviated by addition of exogenous a-factor. T h e pro region peptides produced by these mutants, however, would have aberrant carboxy termini if processing is inefficient, as described below. Because it is difficult to follow the pro region biochemically due to the lack of cysteine and methionine residues, a genetic approach was taken to get some information on processing of the hybrid proteins.
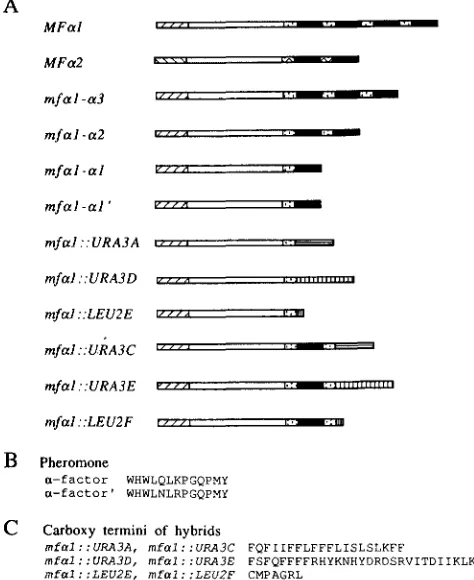
The first step involved in processing of the M F a l precursor is cleavage by the endopeptidase Kex2 at Lys-Arg sequences at the amino termini of each of the M F a l spacer sequences (Figure 2A; ACHSTETTER and WOLF 1985; JULIUS et al. 1984). Kex2 cleavage pro- duces spacer-a-factor intermediates that are processed further to produce mature a-factor (FULLER, STERNE and THORNER 1988); another product of this cleavage should be the MFaI pro region, although this product has not been detected.
The mfal::URA3A, mfal::URA3D and mfal::LEU2E
constructions contain the first M F a l spacer sequence and therefore, the first Lys-Arg sequence (Figure 1A). If Kex2 cleavage at this sequence occurs, the MFa1 pro region peptide should be produced as well as a
polypeptide consisting of the remainder of the first spacer and the hybrid carboxy terminus provided by the end of the URA3 or LEU2 fragment (Figure 2C). If so, the small carboxy terminal peptide, possibly with the spacer residues removed, must account for the inhibition of mating by the mfal::URA3A gene. If transport of the hybrids through the secretory path- way or Kex2 cleavage is inefficient in the context of a hybrid carboxy terminus, however, an unprocessed product consisting of the MFa1 pro region and first spacer followed by the hybrid carboxy terminus would be produced (Figure 2C). If so, the inhibition of mating by mfal::URA3A would result from this un- processed hybrid.
An additional set of constructions provided some information on transport and/or processing of the hybrid proteins. T h e same URA3 (mfal::URA3C and
mfal::URA3E) and LEU2 (mfal::LEU2F) fragments
were inserted into the second MFaI Hind111 site, resulting in fusions containing the first spacer-a-factor repeat followed by the second spacer and the hybrid carboxy termini (Figure 1A). Levels of a-factor were determined in strains also containing an mfa2 null mutation, so that all a-factor is provided by the m f a l gene. T h e control strain contained the mfal-a1 con- struction in which a single a-factor repeat is present within its normal structural context. Transport and processing of the m f a l - a 1 precursor is relatively effi- cient (CAPLAN et al. 1991).
T h e mfaI::URA3C mfa2-null and mfal::URA3E mfa2-null mutants secreted 10-20-fold less a-factor, respectively, than the mfal-a1 mfa2-null mutant (data not shown). This reduction could be due to either inefficient transport or processing of the hybrid pre- cursors. Because the mfal::URA3A and mfaI::URA3D hybrids have the same carboxy termini as the mfal::URA3C and mfal::URA3E hybrids, respectively, it is likely that transport or processing of these hybrids is also inefficient, resulting in production of an un- processed hybrid product (Figure 2C). If so, the un- processed mfaI::URA3A hybrid inhibits mating, but the unprocessed mfal::URA3D hybrid does not.
A
Signal Peptidase
+
c
KexP Endopeptidase +,1
c
Kexl Carboxypeptidase+ + + +
""-
c
I -" "
c
7""
c
Role of a-factor and its precursor
Addition of exogenous a-factor does not alleviate the mating defect of pro-region-containing mutants: Previous results showed that addition of exogenous a- factor to a m f a l m f a 2 double null mutants did not alleviate the mating defect of these mutants in crosses to wild-type a strains (KURJAN 1985), although some alleviation was seen in crosses to supersensitive mu- tants (see DISCUSSION). If the inability of exogenous a- factor to alleviate the a m f a l m f a 2 mating defect is due to a separate role for the pro region itself in mating, then exogenous a-factor should alleviate the mating defect of constructs able to produce the pro region but no a-factor. Because mfal::LEU2E is the most likely hybrid to be transported and processed efficiently, resulting in production of a wild-type pro region product, it provides the best strain with which to test alleviation by exogenous a-factor.
Mating of the a mfal::LEU2E mfa2 in the presence of exogenous a-factor gave results identical to the previous results with null mutants (Table 3; KURJAN
1985). N o alleviation of the mating defect by addition of exogenous a-factor was seen in the crosses to the wild-type a strain. In crosses to an a sstl mutant (CHAN and OTTE 1982), which is supersensitive to phero- mone due to a defect in a secreted activity that de- grades a-factor, some alleviation of the mating defect by addition of exogenous a-factor was seen for both mfal::LEU2E and the null mutant (Table 3). An a sst2
C
+
c
c
c
f"
c
FIGURE 2.-Wild-type and hybrid precursor processing. T h e wild-type and disruption constructs are dia- grammed as in Figure 1 except that the carboxy termini shown in (B) and (C) are illustrated by crosshatched rectangles to represent any of the carboxy termini in the hybrids. Ar- rows indicate positions of processing by the enzyme indicated. Spotted ar- rows indicate sites within aberrant precursors or intermediates, which might therefore be processed ineffi- ciently. T h e a-factor processing path- way has been reviewed by FULLER, STERNE and THORNER (1988). T h e signal sequence is removed from pre- pro-a-factor during transport into the endoplasmic reticulum. T h e KexP endopeptidase cleaves after Lys-Arg residues at the N termini of the spacer peptides. The Kexl car- boxypeptidase removes the C-termi- nal Arg and Lys residues remaining. T h e S t e l 3 diaminopeptidase re- moves the remaining spacer residues by step-wise removal of Glu-Ala or Asp-Ala dipeptides from the N ter- minus of a-factor. (A) Wild-type
M F a l . (B) Disruption constructs with one a-factor repeat. (C) Disruption constructs with no a-factor repeat.
TABLE 3
a-Factor addition experiments
Mating in presence of a-factor (M)"
MATa testerb Strain o 5 X 5 x 10-8 5 x 10"
Wild-type M F a I
+++
+++ +++
+++
mfal::LEU2C - -
-
-mfa1::LEUZE - - -
-
mfal::URA?C
+
+
+
+
sstl-2 M F a l
+++
+++
+++
++
mfal::LEU2C -
-
P+
mfaI::LEU2E - - P
+
mfaI::URA3C
+
+
+
+
a Mating was tested by patch tests.
+++,
wild-type mating;++,
somewhat lower mating;+,
low level of mating; p, papillation-very low level of mating; -, no detectable mating.T h e wild-type strain was RC618 and the sstl-2 strain was RC629.
strain, which shows greater supersensitivity to phero- mone and is defective in desensitization to pheromone (CHAN and OTTE 1982; DIETZEL and KURJAN 1987), was able to mate at a low level to the mfal::LEU2E and null mutants and in both cases, the level of mating was increased by addition of exogenous a-factor (data not shown).
S. J.
and it is possible that transport through the secretory pathway is necessary for the proposed pro region function. Because the a mfal::URA3C mfa2 strain does produce a low level of a-factor, at least a portion of the precursor must be transported and processed. Mating of this strain to the wild-type and the sstl
strains was low and not increased by addition of a- factor (Table 3). Mating to the a sst2 strain was too high in the absence of pheromone to be able to see an effect of pheromone addition. Even in a case where some precursor is transported and processed, there- fore, exogenous a-factor could not alleviate the mat- ing defect.
Structural alterations of the MFal precursor re- sult in proportional decreases in a-factor production and mating: An alternative approach to investigate the proposed second role of the M F a l precursor was to attempt to identify pro region mutants that affect the proposed second role for the M F a l precursor in a mating without eliminating a-factor production. In a separate study, a set of mutations resulting in altered precursor structure was constructed and analyzed for effects on a-factor secretion and precursor build-up (CAPLAN et al. 199 1). These alterations include elimi- nation of sites for addition of N-linked carbohydrate, in-frame deletions and insertions in the pro region, and changes in the number of spacer-a-factor repeats. Another mutant, mfal-pro3, was constructed as de- scribed in Materials and Methods. In particular, the pro region deletions (mfal-pro1 has a 35 amino acid deletion of the central pro region, mfal-pro2 has a 14
amino acid deletion near the start of the pro region, and mfal-pro3 has a deletion of 17 amino acids im- mediately following the signal sequence) might pro- vide the type of mutant described above. T h e effect of these mutations on mating was tested (Table 4).
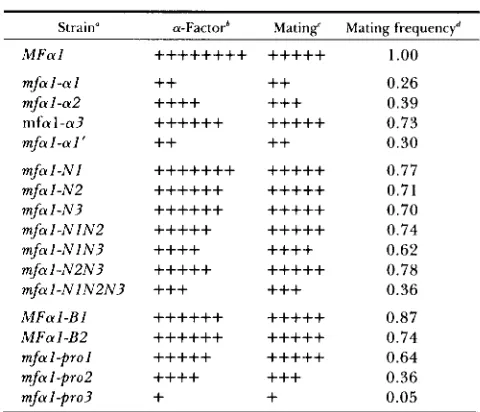
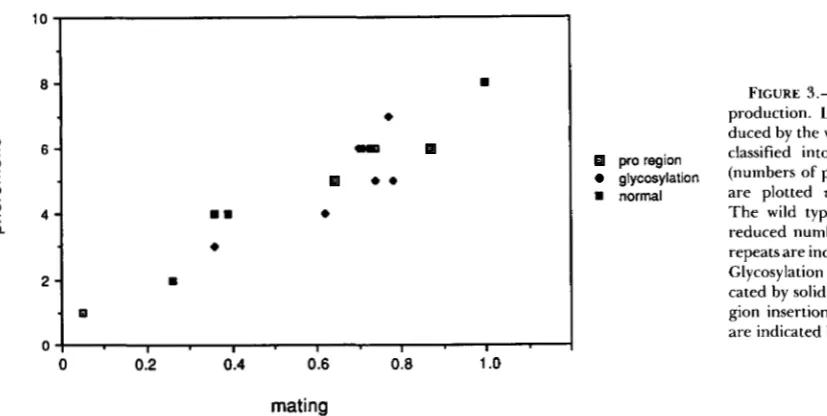
All of the mfal mutants that showed reduced mat- ing also showed reduced a-factor production. There was a striking correlation between the levels of a- factor secretion and the levels of a mating (Table 4,
Figure 3). The comparison to the constructs with reduced numbers of spacer-a-factor repeats was par- ticularly informative. T h e decreased a-factor produc- tion in these mutants can be attributed mostly to fewer copies of a-factor coding sequences in the precursor along with a minor effect due to slight precursor accumulation (CAPLAN et al. 199 1). T h e mutants with alterations of the pro region showed reduced a-factor secretion due to precursor accumulation, probably resulting from inefficient transport through the secre- tory pathway. In all cases, the level of mating was similar to the level of mating of mutants with fewer spacer-a-factor repeats that produce a similar level of a-factor. If the pro region played a separate role in mating, at least some pro region mutants would be likely to show reduced mating in comparison to other
TABLE 4
a-Factor and mating phenotypes
Strain" a-Factorh Mating' Mating frequencyd
MFal
++++++++
+++++
1.oo
mfal-a1
++
++
0.26mfal-a2
++++
+++
0.39mfa 1 -a3
++++++
+++++
0.73mfal-al'
++
++
0.30mfal-Nl
+++++++
+++++
0.77mfaI-N2
++++++
+++++
0.71mfal-N3
++++++
+++++
0.70mfal-NIN2
+++++
+++++
0.74mfal-NlN3
++++
++++
0.62mfal-NZN3
+++++
+++++
0.78mfaI-NlN2N3
+++
+++
0.36MFal-BI
++++++
+++++
0.87M F a 1-82
++++++
+++++
0.74mfal-pro1
+++++
+++++
0.64mfal-pro2
++++
+++
0.36mfal-pro3
+
+
0.05All strains are a mating type, contain the mfa2::LEU2C disrup- tion mutation (KURJAN 1985). and are isogenic to W303-1B (Table 1). T h e a MFal mfa2::LEUZC strain is used as our wild-type strain and shows a-factor expression and a mating levels similar to the isogenic a MFal MFa2 strain (KURJAN 1985). mfal-al, mfal-a2, mfal-a3, and mfal-al' have reduced humbers of spacer-a-factor repeats (Figure l), and in mfal-al', the single a-Factor repeat was mutated to encode a-factor'. Construction of mfal-pro3 is described
in MATERIALS A N D METHODS and construction of the remainder of
the mutants is described elsewhere (CAPLAN et al. 1991). Mutations N 1, N2 and N 3 eliminate sites for N-linked glycosylation by single amino acid changes in the recognition sequences for glycosylation (positions 25, 59 and 69); mutants with single, double, and triple glycosylation site alterations were tested. Other alterations of the pro region include: MFal-BZ, insertion of four amino acids between amino acids 22 and 23; MFal-B2, insertion of four amino acids between amino acids 37 and 38; mfal-prol, deletion of amino acids 29 to 63; mfal-pro%, deletion of amino acids 23 to 36 and insertion of five amino acids a t this position; and mfal-pro3, deletion of amino acids 20 to 36 and insertion of three amino acids at this position.
*
a-Factor levels as determined by halo and dilution assays on an a sst2 strain (RC63 1). In the dilution assays, serial dilutions of wild- type and mutant strains are spotted on lawns of the tester strain. T h e dilutions of the wild-type strain necessary to give a growth arrest spot similar to the mutant strain were:++++++++,
1 X ;+++++++,
4X;++++++,
6X;+++++,
8X;++++,
16X;+++,
3 2 ~ ;
++,
64X; +, >lOOX. Because the values vary depending on the tester lawn used and tester lawn thickness, they give good estimates of relative levels of a-factor produced but cannot be interpreted as accurate a-factor levels.Qualitative mating levels as determined by patch tests. Quantitative mating assays. T h e frequencies are an average of three experiments.
types of mutants producing similar levels of a-factor. This correlation between a-factor and mating levels for mutants with a quite wide range of structural alterations strongly suggests that the only role of the
M F a l precursor is to produce a-factor, and that the function of the pro region is to mediate transport and processing.
Strains producing a-factor or a-factor' show
Role of and precursor 305
I I
8 - 8
e
6 - a
e .
4 - e
e
2 - 8
- 0
0 I I I I I
0 0.2 0.4 0.6 0.8 1 .o
mating
(Figure 1A) and a-factor (Gln-5,Lys-7 a-factor) had similar activities with respect to several aspects of pheromone response that could be assayed by addition of exogenous pheromone (KURJAN and LIPKE 1986; RATHS et al. 1986). To determine the relative activi- ties of these two peptides produced in vivo, constructs allowing synthesis of either a-factor or a-factor’ from
M F a l were constructed. T h e mfal-a1 gene, which encodes an a-factor precursor with only a single M F a l
spacer-a-factor repeat, was mutated to mfal-a1
’,
which is identical except that the a-factor sequence encodes a-factor’ (Figure 1). These constructs were tested in strains containing an mfa2 null mutation, so all a-factor is provided by mfal-a1 or mfal-al‘. Se- cretion of a-factor and mating by the two strains were similar (Table 4), indicating that a-factor’ is as effi- cient as a-factor in eliciting all aspects of pheromone response required for mating.
DISCUSSION
The two forms of a-factor are functionally equiv- alent: Two forms of a-factor are encoded by the
M F a I and M F a 2 genes (Figure 1). T h e minor form (a-factor’) is encoded only by one of the two M F a 2
repeats (KURJAN and HERSKOWITZ 1982; SINGH et al. 1983) and was not detected by amino acid sequence analysis of secreted a-factor (STOTZLER, KILTZ and DUNTZE 1976; TANAKA et al. 1977). Synthetic a-
factor‘ was shown to be equivalent to a-factor in eliciting several aspects of pheromone response (KUR- JAN and LIPKE 1986; RATHS et al. 1986); however, to
investigate the actual function during the mating proc- e s s , it is necessary to examine mating of cells able to secrete only one of the two forms. We found that strains with all of the pheromone provided by an
MFaI gene encoding one copy of either a-factor
(mfal-al) or a-factor’ (mfal-al’) showed no signifi- cant difference in a-factor production or mating
FIGURE 3.-Mating us. a-factor production. Levels of a-factor pro- duced by the various constructs were pro region classified into eight relative levels
glycosylation (numbers of plusses in Table 4) and
rn normal are plotted us. mating frequency.
The wild type and constructs with reduced numbers of spacer-a-factor repeats are indicated by solid squares. Glycosylation site mutants are indi- cated by solid diamonds, and pro re- gion insertion and deletion mutants are indicated by open squares.
(Table
4).
Secretion of the two forms of a-factor is therefore equivalent with regard to all responses nec- essary for mating. Because a-factor’ may be less sen- sitive to degradation than a-factor (SAMOKHIN et al. 1981), this equivalence may actually reflect lower activity combined with greater stability of a-factor’ in comparison to a-factor.The only role of the MFarl precursor is in a-factor production: Previous results showed that addition of exogenous a-factor at levels capable of inducing a cell responses was unable to alleviate the mating defect of a mfal mfa2 null mutants in crosses to wild-type a strains (KURJAN 1985). A possible explanation for these results is that the pheromone precursors play two separate roles in mating; one role is the produc- tion of pheromone, and the second role would be a separate uncharacterized role. T h e inhibition of mat- ing by the mfaI::URA3A product (Figure 1; KURJAN
1985) suggested that the M F a l pro region in partic- ular might play this second hypothesized role. In addition, it has been shown that exogenous phero- mone does allow some alleviation of the mating defect of mutants (stel3 and kex2) that do not produce a-
factor due to mutations in precursor processing en- zymes (CHAN et al. 1983). T h e results with the a-
factor structural gene mutants us. the processing en- zyme mutants suggested that exogenous a-factor can alleviate the mating defect of strains unable to pro- duce a-factor only if a-factor precursor can be made. If the inability of exogenous a-factor to alleviate the mating defect of a mfal mfa2 mutants is due to the need for a function provided by the pro region, an mfal mutant able to produce the pro region but no a-factor should show alleviation of the mating defect by addition of exogenous a-factor. T h e
mfal::LEU2E mfa2 mutant (Figure 1) should be such a mutant; it is likely to produce a pro peptide identical
306 S. Caplan and J. Kurjan
inhibit mating (Table 2). Mating assays with exoge- nous a-factor added to the a mfal::LEU2E mfa2 mu-
tant showed identical results to assays with a mfal-
null mfa2 mutants (Table 3; KURJAN 1985), ie., no alleviation of the mating defect was seen in crosses to the wild-type a strain. T h e alleviation of the mating defect in crosses to supersensitive a strains was also identical with the null mutants and with the rnfal::LEU2E mutant; implications of this result are discussed below. Alleviation assays were also done with the a mfal::URA3C mfa2 mutant, for which at least a portion of the precursor is transported through the secretory pathway and processed (because some a-factor is produced). Mating with this mutant was low in the absence of exogenous pheromone and was not alleviated by pheromone addition (Table 3). These results suggest that the pro region does not play a role separate from a-factor production that together with addition of exogenous a-factor can al- low a mating. A possible problem with this interpre- tation, however, is that we cannot be certain that transport, processing and modification of the pro region of these hybrid proteins occurs normally, and the hypothesized second role for the pro region might require that these steps be carried out efficiently.
Due to the uncertainties of the experiments de- scribed above, a second and complementary approach aimed at identifying a role for the pro region separate from a-factor production was taken. This approach involved an attempt to identify M F a I mutations that would allow a-factor production but result in a mating defect. Mutants with structural changes in the M F a I precursor were constructed and used to study the role of the a-factor precursor structure in a-factor secre- tion (CAPLAN et al. 1991). Most of these mutations cause decreased a-factor production and intracellular accumulation of precursor, suggesting that transport of the precursor through the secretory pathway is very sensitive to structural features of the precursor. In this paper, we examined the effect of these muta- tions on mating.
If the pro region plays a role in mating separate from a-factor production, some of the pro region mutations, particularly the deletions, might severely reduce mating. In particular, in a comparison of a pro region mutant and a mutant with a reduced number of spacer-a-factor repeats, both of which show similar levels of a-factor production, the pro region mutant should show a lower level of mating. In contrast, the mutants showed a striking correlation between the decreases in a-factor secretion and the decreases in mating (Table 4, Figure 3). This correlation argues against a separate role for the pro region in mating and suggests that the only role of the M F a I precursor is to produce a-factor. It remains possible that we did not alter the particular residues of the pro region
responsible for the hypothesized second role in mating in these experiments, although the deletion mutants covered most of the pro region. Given two comple- mentary sets of experiments that lead to the same conclusion, however, we conclude that it is unlikely that the pro region plays a role in mating separate from its role in transport and processing to produce a-factor. An explanation for the inhibitory effect on a mating by the mfal::URA3A gene product remains unclear (KURJAN 1985).
Implication for the role of pheromone in mating: If the a-factor precursor does not play a role separate from a-factor production, the lack of alleviation of the mfal mfa2 mating defect by addition of exogenous a-factor in crosses to wild-type strains must have an alternative explanation. Analogous results have been demonstrated for a cells; exogenous a-factor is unable to alleviate the mating defect of a mfal mfa2 null mutants (MICHAELIS and HERSHOWITZ 1988), indicat- ing that both a and a cells must be actively secreting pheromone to mate efficiently. A pair of mating a and a cells in close proximity grow towards one another (HERMAN 1971). Products involved in direct contact and fusion of the cells (the a-agglutinin and FUS gene products; Truehart, BOEKE and FINK 1987; WATZELE, KLIS and TANNER 1988) are present at high levels on the shmoo tip (site of cellular elongation) after expo- sure to the opposite pheromone and are thought to fuse at the position of elongation. Secretion of pher- omone may also occur from the shmoo tip, resulting in high localized concentrations of pheromone be- tween the fusing pair. Alternatively, a gradient of pheromone levels, with very high levels near the shmoo tip and lower levels further from the shmoo tip, might be necessary for efficient mating. Achieving either a very high concentration or a gradient of pheromone in the vicinity of the shmoo tip may be difficult by addition of exogenous pheromone.
T h e ability of exogenous a-factor to partially alle- viate the mating defect of a mfal mfa2 mutants in crosses to a sstl strains (Table 3; KURJAN 1985) is consistent with this hypothesis. The sstl (also called b u r l ) mutation causes increased sensitivity of a strains to a-factor due to a defect in a-factor degradation. Addition of exogenous a-factor in crosses of the a
mfaI mfa2 mutants to the a sstl strain should be equivalent to addition of an extremely high level of a-factor to a wild-type strain. In this situation, exog- enous a-factor may be able to provide the very high level of a-factor normally present in the vicinity of the shmoo tip in cells secreting a-factor. In the course of mating, adjacent a and a cells may be able to “sense” the mating competence of one another. Consistent with this idea, recent results (JACKSON and HARTWELL
produce a-factor makes it a better competitor, indi- cating a role for the actual secretion of pheromone in signaling between adjacent a and (Y cells.
We thank LYNN MIYAZAKI for technical assistance and Jeanne Ilirsch for comments on the manuscript. T h e work was supported by grants from the National Institutes of Health and the American < h c c r Society.
LITERATURE CITED
ACHSTETTER, T . , and D. H. WOLF, 1985 Hormone processing and tnembrane-bound proteinases i n yeast. EMBO J. 4: 173-
177.
<:APLAN, S . , R. GREEN, J. ROCCO and J. KURJAN, 1991 Glycos- ylation and structure of the yeast M F a l a-factor precursor is itnportant for efficient transport through the secretory path- way. J. Bacteriol. 1 7 3 (in press).
CHAN, R. K., and C. A. OTTE, 1982 Isolation and genetic analysis of Saccharomyces cerevisiae mutants supersensitive to GI arrest by a-factor and a-factor pheromones. Mol. Cell Biol. 2: 11-20.
(:HAN, R. K., L. M. MELNICK, L. C. BLAIR and J. THORNER,
IS83 Extracellular suppression allows mating by pheromo- nedeficient sterile mutants of Saccharomyces cerevisiae. J. Bac- teriol. 1 5 5 903-906.
(:ROSS, F., L. H. HARTWELL, C. JACKSON and J. B. KONOPKA,
1988 Corljugation in Saccharomyces cerevisiae. Annu. Rev. Cell Biol. 4: 430-457.
hKTXEt., C., and J. KURJAN, 1987 Pheromonal regulation and sequence of the Saccharomyces cerevisiae SST2 gene: a model for desensitization to pheromone. Mol. Cell. Biol. 7: 4169-4177. I)UTCHER, S. K., and L. H. HARTWELL, 1982 T h e role of S.
cereuisiae cell division cycle genes in nuclear fusion. Genetics
100: 175-184.
I;UI.I.ER, R. S . , R. E. STERNE and J. THORNER, 1988 Enzymes required for yeast prohornlone processing. Annu. Rev. Physiol.
50: 345-362.
HARTWELL, L . H., 1980 Mutants of Saccharomyces cerevisiae un- responsive to cell division control by polypeptide mating hor- mone. J. Cell Biol. 85: 81 1-822.
H E R M A N , A. K., 1971 Sex-specific growth responses in yeasts. Antonie Leeuwenhoek 37: 379-384.
JACKSON, C. L., and L. H. HARTWELL, 1990 Courtship in Saccha-
romyces cerevisiae: an early cell-cell interaction during mating. Mol. Cell. Biol. 10: 2202-2213.
JUI.IUS, D., A. BRAKE., L. BLAIR., R. KUNISAWA and J. THORNER,
1984 Isolation of the putative structural gene for the lysine- arginine-cleaving endopeptidase required for processing of yeast prepro-a-factor. Cell 37: 1075-1089.
KURJAN, J., 1985 a-Factor structural gene mutations in Saccharo- myces cerevisiae: effects on a-Factor production and mating. Mol. Cell. Biol. 5: 787-796.
KURJAN, J., and I . HERSKOWITL, I982 Structure of a yeast pher-
omone gene ( M F a ) : a putative a-factor precursor contains four tandem copies of mature a-Factor. Cell 30: 933-943.
KURJAN, J., and P. N. LIPKE, 1986 Agglutination and nlating activity of the MFa2-encoded a-factor analog in Saccharomyces cerevisiae. J . Bacteriol. 168: 1472-1475.
MACKAY, V. L., and T . R. MANNEY, 1974 Mutations affecting sexual co~~jugation and related processes in Saccharomyces cere-
visiae. I . Isolation and phenotypic characterization o f non-
mating mutants. Genetics 76: 255-271
MANNEY, T. R., and V. WOODS, 1976 Mutants of Saccharomyces
cerevisiae resistant to the a tnating-type factor. Genetics 82:
639-644.
MICHAELIS, S., and I . HERSKOWITL, 1988 T h e a-factol- phero- mone of Saccharomyces cereuisiae is essential for mating. Mol. Cell. Biol. 8: 1309-1 3 18.
PARKER, R. C., R. M. WATSON and J. VINOCRAD, 1977 Mapping of closed circular DNAs by cleavage with restriction endonu- cleases and calibration by agarose gel electrophoresis. Proc. Natl. Acad. Sci. USA 74: 851-855.
RATHS, S., P. SHENBAGAMURTHI, F. NAIDER and J. M. BECKER, 1986 Biological activity of the Asn-5,Arg-7 tridecapeptide encoded by MFa2 of Saccharomyces cerevisiae. J. Bacteriol 168:
1468-1471.
ROTHSTEIN, R. J., 1983 One-step gene disruption in yeast. Meth- ods Enzynlol. 101: 202-21 1.
SAMOKHIN, G. P., L. V. LIZLOVA, J. D. BESPALOVA, M . I . T I T O V
and V. N. SMIRMOV, 1981 T h e effect of a-factor on the rate of cell cycle initiation in Saccharomyces cerevisiae. Exp. Cell Res.
SANGER, F., S. NICKLEN and A. R. COULSON, 1977 DNA sequenc- ing with chain-terminating inhibitors. Proc. Natl. Acad. Sci. USA 74: 851-855.
SINCH, A., E. Y . CHEN, J. M. I~UGOVOY, C. N. (:HANG, R. A. HITZMAN and P. H. SEEBURG, 1983 Saccharomyces cerevisiae
contains two discrete genes coding for the a-factor pheromone. Nucleic Acids Res. 11: 4049-4063.
STOTZLER, D., H. -H. KILTZ and W. DUNTZE, 1976 Primary structure of a-factor peptides from Saccharomyces cerevisiae.
Eur. J. Biochem. 69: 397-400.
TANAKA, T., H. KITA, T . MURAKAMI and K. NARITA, 1977 Purification and amino acid sequence of mating factor from Saccharomyces cerevisiae. J. Biochem. 82: 1681- 1687. TRUEHART, J., J. D. BOEKE and G. R. FINK, 1987 T w o genes
required for cell fusion during yeast conjugation: evidence for a pheromone-induced surface protein. Mol. Cell. Biol. 7: 2 3 16-
2328.
WATZELE, M., F. KLIS and W . T A N N E R , 1988 Purification and characterization of the inducible a ag-glutinin of Sacrharomyces cerevisiae. EMBO J. 7: 1483-1488.
ZOLLER, M. J., and M. SMITH, 1983 Oligonucleotide-directed mutagenesis of DNA fragments cloned into M 13 vectors. Meth- ods Enzymol. 100: 468-500.
131: 267-275.