1
ABSTRACT 2
OLSON, ERIC LEONARD. Characterization of Stem Rust Resistance in US Wheat 3
Germplasm. (Under the direction of Gina Brown-Guedira.) 4
5
In 1999 in Uganda a race of stem rust, Puccinia gramins f. sp. tritici was 6
identified with virulence to Sr31. This race, designated as TTKS based on the North 7
American nomenclature system, combined Sr31 virulence with virulence to the majority 8
of Triticum aestivum L. derived stem rust resistance genes. The development of resistant 9
cultivars is needed as TTKS may reach global dispersal due to its unique virulence to 10
multiple known and unknown resistance genes and widespread cultivar susceptibility. 11
The ability to detect the presence of specific stem rust resistance genes using molecular 12
markers presents a viable method for identifying resistance to race TTKS in the absence 13
of the pathogen itself. The frequency of DNA markers associated with resistance genes 14
Sr24, Sr26, Sr36, and Sr1RSAmigo which confer resistance to TTKS was assessed in 15
diverse wheat cultivars and breeding lines from breeding programs throughout the United 16
States. The reliability of these markers in predicting the presence of the resistance genes 17
in diverse germplasm was evaluated through comparison with phenotypic data. 18
Introgression of undeployed seedling resistance genes is necessary to improve the 19
availability of resistance to TTKS. The stem rust resistance gene Sr22 confers resistance 20
to TTKS. Sr22 is present on a chromosomal translocation derived from Triticum 21
boeoticum Boiss. which is homoeologous to the A genome of T. aesitivum Linkage 22
analysis of SSR loci on 7AL was done to identify the loci most closely linked to Sr22. 23
populations of crosses between the germplasm stock Sr22Tb and the hard winter wheat 1
lines 2174 and Lakin. From analysis of F3:4 populations derived from F2 recombinants, 2
F3:4 individuals with further reduced translocation segments have been identified. 3
Recombinant lines with reduced translocations will provide a more agronomically 4
desirable source of Sr22 stem rust resistance in hard winter wheat germplasm that can be 5
readily deployed utilizing molecular markers. The identification of molecular markers 6
efficacious for the selection of genes for resistance to TTKS will hasten the development 7
by 3
Eric Leonard Olson 4
5
A thesis submitted to the Graduate Faculty of 6
North Carolina State University 7
in partial fulfillment of the 8
requirements for the Degree of 9
Master of Science 10
11 12
Crop Science 13
14 15
Raleigh, North Carolina 16
17 18
2009 19
20 21
APPROVED BY: 22
23 24 25 26
_______________________ _________________________ 27
Dr. David Marshall Dr. James B. Holland 28
29 30 31 32 33 34
__________________________ 35
Dr. Gina Brown-Guedira 36
1 2
BIOGRAPHY 3
4
Eric Leonard Olson was born in Dodgeville, WI in 1980 to Leonard and Catherine 5
Olson. Eric, the oldest of three siblings, lived and worked on the family dairy farm for 6
many years. The best years of his life were spent working beside his brother, father and 7
grandfather on the farm. Attending the University of Wisconsin in Platteville, Eric 8
developed a love of science and a desire to make meaningful contributions to agriculture 9
through science. Opportunity for graduate studies at North Carolina State University was 10
available and in 2007 Eric began work on an MS degree with Dr. Gina Brown-Guedira. 11
In January of 2009 Eric will begin a Phd. program at Kansas State University working 12
with Dr. Michael Pumphrey and Dr. Bikram Gill. 13
1 2
ACKNOWLEDGEMENTS 3
4
I would like to thank most of all those few who let me believe that graduate 5
school was a possibility. Thank you to my family for teaching me how to work hard and 6
for being there for me, always. Thank you to Dr. Gina Brown-Guedira for the 7
opportunity to do challenging and meaningful work. I am grateful to Jared Smith and 8
Kim Howell for sharing their valuable technical expertise. I sincerely thank Dr. Michael 9
Pumphrey for contributing populations and providing phenotypic evaluations. Thank 10
you to my committee members Dr. Jim Holland and Dr. David Marshall. A special 11
thanks to Dr. Gina Brown-Guedira and Dr. David S. Marshall for the opportunity to 12
travel to Kenya. Many thanks to all who took time to listen and helped me learn through 13
meaningful discussion. 14
1
TABLE OF CONTENTS 2
3
Page 4
LIST OF TABLES………...vi 5
6
LIST OF FIGURES………..…...vii 7
8
CHAPTER I. Literature Review.……….1 9
Importance of Wheat Cultivation to Humans….……… 2 10
Significance of Wheat………...2 11
Wheat Evolution and Cytogenetics………...4 12
Origins of modern wheat………...4 13
Allopolyploidy……….………7 14
The Use of Wheat Relatives in Breeding for Disease Resistance………...8 15
Wheat germlplasm resources...………...………...…8 16
Introgression Methods……….9 17
Ph1 Mutants……….9 18
Gametocidal Genes………..………..10 19
Radiation………11 20
The StemRust Pathogen……….12 21
Historical Impact………12 22
Life Cycle………...13 23
Infection Process………16 24
Physiologic Races………..17 25
Population Genetics and Evolution………18 26
New Highly Virulent Pgt Race………..19 27
Stem Rust Resistance……….22 28
Sr and Avr gene interaction………22 29
Stem Rust Resistance Genes………..………22 30
Resistance to Ug99………..…. 24 31
Development of Resistant Germplasm………...………...23 32
References………..………28 33
34
CHAPTER II. Genotyping of U.S. Wheat Germplasm for Presence of Stem Rust 35
Resistance Genes Sr24, Sr36 and Sr1RSAmigo 36
Abstract………..47 37
Introduction………48 38
Materials and Methods………...51 39
Marker Analyses………52 1
Phenotypic Analysis………...54 2
Results……….………...55 3
Discussion………..61 4
References………..…..…….…….65 5
6 7
CHAPTER III. Genetic Characterization of Stem Rust Resistance Gene Sr22 8
Abstract………..…..………..……76 9
Introduction………..…..……....…....77 10
Materials and Methods………..…..…….…...…...81 11
Plant Materials………..…..…………...…81 12
Stem Rust Evaluations………..…..…………...83 13
Molecular Marker Analyses………..…..………...84 14
Results………..…..………86 15
Phenotypic Evaluation…………...………..…..………86 16
Genetic and Physical Mapping of Sr22Tb Introgression.………...…...…87 17
Linkage Analysis of F3:4 Recombinant populations…………...…………90 18
Identification of Recombinants………...93 19
Discussion………..94 20
References……….……….98 21
LIST OF TABLES 1 2
CHAPTER II. Genotyping of U.S. Wheat Germplasm for Presence of Stem Rust 3
Resistance Genes Sr24, Sr36 and Sr1RSAmigo 4
5
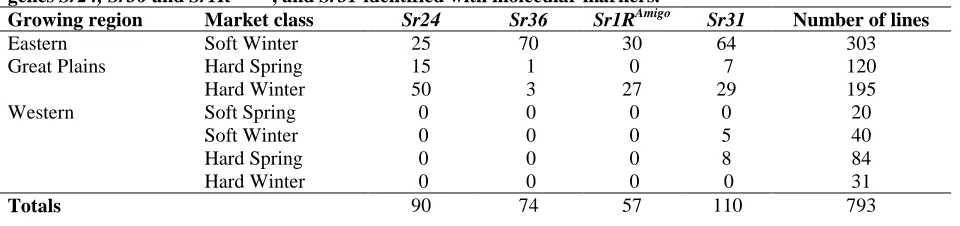
Table 1. Number of U.S. wheat lines from different regions and market classes 6
having stem rust resistance genes Sr24, Sr36 and Sr1RAm , and Sr31 identified 7
with molecular markers.……….………71 8
9
Table 2. Species of origin, chromosomal location, diagnostic markers and 10
expected size in base pairs of amplified fragments for selection of Sr24, Sr36, 11
Sr1RAmigo, and Sr26.…….……….…...……..………71 12
13
Table 3. Effective Sr1RAmigo resistance in the presence 14
of Sr24 virulence………...……….…71 15
16
Table 4. Stem rust resistance gene pyramids present………72 17
18
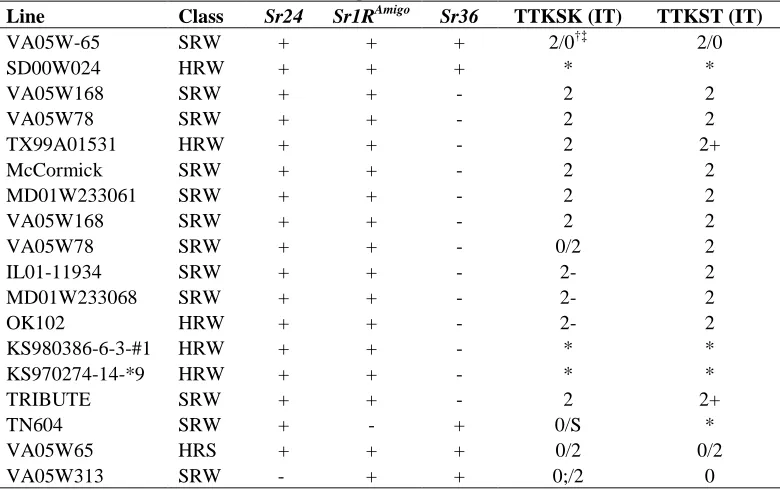
Table 5. Lines resistant to TTKSK and TTKST without marker alleles for 19
Sr24, Sr36 or Sr1RAmigo...72 20
21
CHAPTER III. Genetic Characterization of Stem Rust Resistance Gene Sr22 22
23
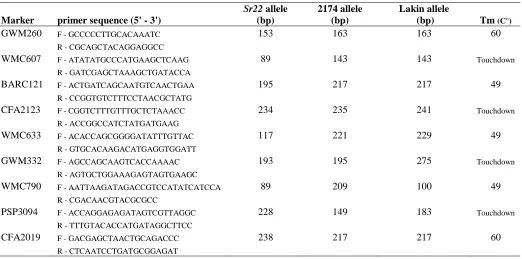
Table 1. Markers used for linkage analysis of Sr22, primer sequences, parent 24
allele sizes in base pairs and primer annealing temperatures (Tm) in 25
degrees Celsius ………..…102 26
27
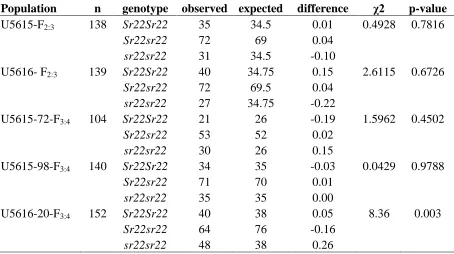
Table 2. Segregation of Sr22 in F2:3 and F3:4 populations…….………103 28
29
Table 3. Alleles of the Sr22Tb donor parent, the cultivar Steinwedel and 30
the hard winter wheat cultivars 2174 and Lakin for six markers...104 31
LIST OF FIGURES 1 2
CHAPTER II. Genotyping of U.S. Wheat Germplasm for Presence of Stem Rust 3
Resistance Genes Sr24, Sr36 and Sr1RSAmigo 4
5
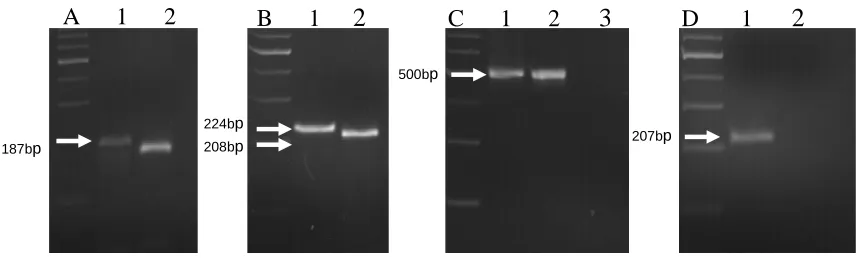
Figure 1. PCR amplification of markers for selection of 6
Sr36, Sr1RAmigo, Sr24 and Sr26…………....………..73 7
8
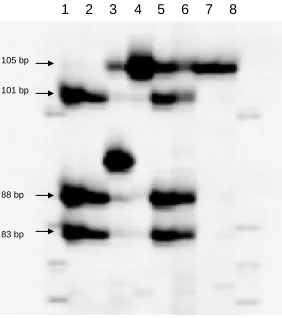
Figure 2. PCR amplification of BARC71 for the identification of Sr24 and 9
differentiation between the 3DL/3Ae translocation derived from ‘Agent’ and the 10
1BL∙1BS-3Ae translocation derived from ‘Amigo’ of Sr24……...………...74 11
12
CHAPTER III. Genetic Characterization of Stem Rust Resistance Gene Sr22 13
14
Figure 1a-b.Genetic linkage of U5615 and U5616 F 2:3 populations 15
segregating for stem rust resistance from Sr22 on wheat chromosome 7AL…105 16
17
Figure 2a-c. Genetic linkage of F3:4 populations derived from recombinant F2 18
individuals in the U5615 and U5616 populations………106 19
20
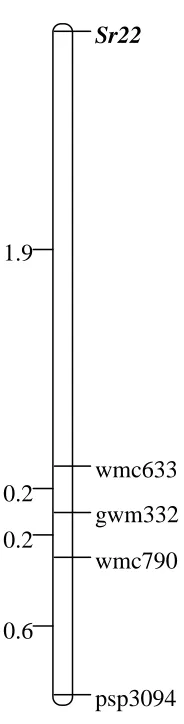
Figure 3.Genetic linkage map of SSR loci linked to Sr22 and segregating in 21
F3:4 populations U5615-72 and U5615-98………107 22
23
Figure 4. Physical map of SSR loci linked to Sr22 on the long arm of 24
chromosome 7A……….…..108 25
26
Figure 5. Physical maps of the long arm of chromosome 7A showing Triticum 27
boeoticum chromatin in recombinants identified from 28
2174/Sr22Tb (U5615) and Lakin/Sr22Tb (U5616) F2:3 and F3:4 29
Chapter 1 1
Literature Review 2
Importance of Wheat Cultivation to Humans 1 2
Significance of Wheat 3
The hunter-gatherer behavior of early societies began to change near the end of 4
the Pleistocene era due to several factors including climate change, availability of game 5
resources and increasing population densities (Diamond, 2002). With larger populations, 6
the more accessible and available foods including fruits and large game became more 7
scarce, leading to a selection of foods requiring more processing like grinding or 8
leaching. Cereals were ideal for this purpose and underwent selection and domestication 9
as population densities increased and the cultivation of cereals became a means of 10
providing a stable, surplus of food resources. 11
The domestication of cereal crop species including barley, rye and wheat was a 12
principal step in the formation of modern agrarian societies. The sowing of grasses 13
allowed for the production of an annual supply of food in one location instead of 14
migrating seasonally to sources of food or continuously hunting and gathering. Wild 15
wheat relatives were abundant in Mesopotamia. Population densities rose beyond the 16
capacity of the native species leading to migrations into more marginal areas less suited 17
for wheat relatives resulting in active cultivation of cereals and the advent of agriculture 18
Larger groups of individuals could congregate in a single location, and as food 1
availability became more secure, individuals within groups were allowed freedom for 2
activities other than food acquisition. Specialized trades and development of new 3
technologies became possible, as individuals were not constrained to the immediate 4
necessity of finding food. The complexity of groups and specialization of trades within 5
groups increased with a sedentary lifestyle and the availability of a stable food supply 6
(Bender, 1978). 7
Climate changes, which led to the migration of groups from more arid regions, 8
resulted in a wider dispersal of cereals. Wheat is a robust cereal which can be grown in 9
environments experiencing abiotic and biotic stresses. Wheat is a broadly adaptable crop 10
that can be grown in environments unsuitable for some other staple crops. It can be 11
produced in arid and semi-arid regions which experience little annual rainfall and high 12
winds, such as Kazakhstan, where nearly 12 million hectares of wheat are now grown 13
(Meng, 2000). Wheat underwent selection for adaptation to many environments as 14
migrations continued. 15
Wheat is a major staple food crop in modern societies, providing 20% of the 16
caloric intake globally (Porter et al., 2007). The wide adaptation of wheat permits its 17
cultivation from the equator to 60°N and 44°S and at elevations from sea level to 3000 m 18
and greater. The leading wheat producing nations are China, India, and the United 19
of 600 million tons of wheat are produced annually, a large part of which is from 1
developing countries. In 2008, 671 million tons were produced globally (Vocke and 2
Alan, 2008). Only 10% of wheat produced is exported, with developing counties 3
consuming most exports (Aquino et al. 2002). Increasing modernization and 4
industrialization on a global scale has changed the diets of developing countries, leading 5
to increased consumption of grains and an increased global demand (Brown, 2004). 6
7
Wheat Evolution and Cytogenetics 8
9
Origins of modern wheat 10
Cultivated wheat, Triticum aestivum L., is an allohexaploid species (2n = 6x = 42) 11
derived from the union of three separate diploid genomes composed of a base 12
chromosome number of seven. The Triticum genus is assembled into three major 13
taxonomic groups: einkorn, emmer or durum wheat, and modern common wheat, based 14
on chromosome number. Einkorn wheats are diploid (2n = 2x = 14), emmer wheat is 15
tetraploid (2n = 4x = 28) and modern common wheat is hexaploid (2n = 6x = 42) 16
(Bonjean, 2001). 17
The three separate genomes of common wheat were derived from distinct 18
ancestral species which diverged from a common progenitor 2.5 to 4.5 million years ago 19
Triticum urartu Thum. ex. Gandil with the AA genomic constitution. The original 1
hybridization event leading to tetraploid wheat combined the A genome of T. urartu with 2
another species closely related to the modern Aegilops speltoides Taush. (BB), resulting 3
in a fertile tetraploid (AABB). The specific donor of the B genome has not been 4
identified due the extinction of the original donor or the possibility of multiple 5
hybridization events (Schneider et al. 2008). The resulting wild tetraploid species, 6
Triticum turgidum L. ssp. dicoccoides (Körn. ex Aschers. & Graebn.) Thell. was 7
subsequently domesticated to become emmer wheat of the species Triticum turgidum L. 8
ssp. diccoccum Schrank ex Schübler. The tetraploid possessed the genomic resources of 9
both diploid ancestors and therefore exhibited greater vigor, and was more adaptable to a 10
broader array of environments than the original diploid progenitors. These traits allowed 11
emmer wheat to be grown across the climates of the Mediterranean as human populations 12
spread beyond Mesopotamia (Zohary and Hopf, 1993). 13
A second hybridization event occurred ~7000 years ago that led to the 14
development of modern bread wheat (Zohary and Hopf, 1993). The cultivation of T. 15
turgium northward out of the Mesopotamia brought emmer wheat into contact with the D 16
genome donor, Aegilops taushii Boiss., with the genomic constitution of DD. The 17
hybridization event leading to modern hexaploid wheat was between the tetraploid 18
emmer wheat (AABB) and the diploid Ae. taushii (DD). The resulting hybrid was the 19
Under normal reproductive conditions involving two haploid gametes from the 1
the tetraploid parent and the diploid parent, the resulting triploid progeny would be 2
sterile. Two conditions exist in which viable progeny are possible from the hybridization 3
of the tetraploid and diploid. One is the possibility the union of 2n gametes from both 4
parents. A 2n gamete that is AABB could be fertilized by a 2n DD gamete to form the 5
tetraploid. The reciprocal cross is possible but less likely due to the potential genetic 6
benefits of AABB maternal cytoplasm. Another possibility is the self fertilization of 7
unreduced gametes in pollen and egg of the triploid progeny of the union of AB and D 8
gametes. Both pollen and egg would be ABD resulting in an AABBDD diploid 9
individual. An additional possibility is the somatic doubling of chromosomes in the 10
triploid resulting from the union of the AB and D haploid gametes. If a mitotic mis- 11
division took place early in embryo development, or in meristematic tissue giving rise to 12
gametic cells, and the diploid chromosomes were doubled, the result would be an 13
AABBDD cell that would give rise through meiosis to gametes with even chromosome 14
numbers that would produce fertile AABBDD progeny. 15
The introgression of the D-genome conferred multiple beneficial traits to 16
hexaploid wheat. Ae. tauschii was adapted to the more continental climate of central 17
Asia expanded the geographical adaptation of hexaploid wheat beyond that of tetraploid 18
wheat (Zohary and Hopf, 1993). The D-genome carries alleles for friabilin proteins 19
and glutenin proteins that trap CO2 during yeast fermentation. The combination of these 1
traits made hexaploid wheat suitable for the baking of leavened bread (Chantret et al., 2
2005). Factors including improved geographical adaptation and improved end use 3
characteristics have allowed for the modern wide scale cultivation of hexaploid wheat. 4
5
Allopolyploidy 6
Major genomic changes took place during the course of polyploidization 7
involving genetic and epigenetic changes. Structural, genetic alterations occurred to 8
genomic DNA sequences and chromosome structure. These structural alterations led to 9
functional, epigenetic changes in gene expression (Levy and Feldman, 2004). 10
Regions of duplicated function underwent large scale deletion. Feldman et al. 11
(1997) and Liu et al. (1997) identified homologous sequences present in diploid wheat 12
relatives that are present in only a single genome of tetraploid or hexaploid wheat, 13
suggesting deletion of some duplicated sequences upon polyploidization. 14
The formation of tetraploid wheat was also accompanied by translocation events. 15
In the A genome diploid ancestor or early in the tetraploid ancestor, chromosomes 4AL 16
and 5AL exchanged terminal segments. In a later translocation in the tetraploid genome 17
a segment of 5AL present on 4AL was exchanged with a terminal segment of 7BS, 18
Genome-wide methlyation patterns influence gene expression and gene silencing. 1
Studies of methylation patterns of cysteine residues indicated changes in methylation 2
status at 13% of genomic loci analyzed through methylation or demethylation (Shaked et 3
al., 2001). Kakush et al. (2002) observed novel gene expression between diploid parents 4
and synthetic allotetraploid progeny, with 48 observed transcripts present in the diploid 5
parents and absent in the allotetraploid. In the tetraploid, 12 transcripts were present that 6
were absent in the diploids. The silencing was associated in part, but not completely, 7
with methylation. 8
9
The Use of Wheat Relatives in Breeding for Disease Resistance 10
11
Wheat germlplasm resources 12
Gerplasm resources available for trait introgression include a wide array of 13
species within Poaceae with a base chromosome number of seven including the several 14
hundred genera within the Triticeae. The genepool available for trait introgression 15
include the species itself, T. aestivum, related species with which T. aestium can be 16
crossed readily to produce fertile offspring, and species to which wide crosses can be 17
made but require special techniques such as embryo rescue to obtain progeny. These 18
resources comprise the primary, secondary and tertiary gene pools, respectively (Harlan, 19
The genomes of wheat progenitor species are a reservoir of valuable alleles that 1
may be introgressed into modern cultivated wheat. Multiple traits have been introgressed 2
including stress tolerance and robustness, drought tolerace (rye translocations) (Villareal, 3
1990), grain quality (Pina, Pinb) (Bonafede et al., 2007), seed storage proteins (high 4
molecular weight glutenins, Payne et al., 1982), and disease resistance. As abiotic and 5
biotic stresses present in wheat growing regions change, the diversity of genetic resources 6
in the primary gene pool of T. aestivum can be improved with alleles for the traits of 7
interest from secondary and tertiary germplasm. 8
9
Introgression Methods 10
Ph1 Mutants 11
The Ph1 gene present on chromosome 5BL is the principal factor responsible for 12
the diploid meiotic behavior of hexaploid wheat. Under normal meiotic conditions in the 13
presence of the Ph1 gene, homoeologous chromosomes do not readily form chiasmata or 14
undergo recombination events (Riley &Chapman 1958, Sears 1977). A mutant stock of 15
the cultivar Chinese Spring designated ph1b has a 70 Mb deletion in the region of 5BL 16
carrying the Ph1 gene (Gill and Gill, 1991), in which pairing of homoeologous 17
chromosomes is evidenced by the formation of trivalents and higher order associations 18
(Riley, 1960). Another deletion of Ph1 exists as ph1c mutants of tetraploid T . turgidum 19
(Riley et al. 1968). The inhibition of Ph1 effects allows for homoeologous 1
recombination, which is a desireable method of alien chromosome integration, due to the 2
compensating effects of the resulting recombinants where wheat chromatin is replaced by 3
alien chromatin. 4
The technique of Ph1 inhibition has been used to introgress multiple disease 5
resistance genes and agronomic traits into T. aestivum germplasm. Zhang et al. (2005) 6
were able to separate the leaf rust seedling resistance gene Lr19 from the yellow pigment 7
gene Y present on chromosome 7E from the Lohopyrum ponticum (Podp.) using ph1b 8
mutants. Kuraparthy et al. (2007) used a PhI stock to minimize Ae. geniculata chromatin 9
from chromosome 5D to less than 5% of the chromosome arm. Lukaszewski (2000) used 10
ph1b mutants to develop recombinant 1RS∙1BL rye chromosomes that do not carry the 11
Sec-1 locus, thereby ameliorating the negative effects of the 1RS rye chromosome on 12
bread-baking quality. Bonafede et al. (2007) used the ph1b mutant to reduce the alien 13
chromatin of chromosome 5A introgressed from T. monococcum containing the Ha locus 14
which carries alleles for grain softness, including the Pina-Am1, Pinb-Am1, and GSP- Am1. 15
16
Gametocidal Genes 17
Random breakage and reformation of chromosomal segments can be induced with 18
the gametocidal genes (Gc) from Aegilops species (Masoudi-Nejad et al. 2002). 19
substitution and addition lines, in which certain chromosomes were maintained during 1
backcrossing between Aegilops species and T. aestivum (Maan, 1975). Chromosomes 2
carrying Gc genes ensure their transmission by causing chromosomal breakage in 3
gametes not carrying the Gc genes. Their activity can range from complete lethality to 4
semi-lethality (Endo, 1990). The female gametes with chromosomal breakage can be 5
fertilized to produce offspring with chromosome aberrations that are stabilized in 6
subsequent generations (Endo, 1988). 7
Gametocidal chromosomes are derived from three diploid genomes, C, S, or M, 8
and are in homoeologous groups 2, 3, or 4 (Endo, 2007). The chromosomal aberrations 9
caused by Gc chromosomes can be highly beneficial as research tools. Deletion stocks 10
have been created for physical mapping of chromosomes allowing for approximations of 11
genes of interest (Endo and Gill, 1996). Further, the disruption of chromosomes allows 12
for breakage and fusion necessary to break up large linkage blocks of introgressed alien 13
chromatin carrying disease resistance genes. Masoudi-Nejad et al. (2002) used a Gc 14
system to recover 1RS chromosomes possessing reduced rye chromatin while 15
maintaining the locus carrying Sr31, Yr9, and Lr26. 16
17
Radiation 18
A physical method of disrupting chromosome segements is the use of ionizing 19
irradiated inducing chromosomal breakage and fusion. Radiation frequently induces 1
reciprocal translocation between the alien chromosome and the wheat chromosome 2
(Badaeva et al., 2007). By inducing breakage of the alien chromosome and fusion of a 3
segment carrying the alleles of interest with a broken wheat chromosome, a reduction in 4
alien chromatin can be achieved. Sears (1956) achieved a successful transfer of leaf rust 5
resistance from Aegilops umbellata using ionizing radiation. Sears (1972) also produced 6
lines carrying Sr24/Lr24 from the 3D/3Ae#1 translocation from Agropyron elongatum. 7
The Sr24/Lr24 translocation has been one of the most widely deployed translocations 8
developed by ionizing radiation. 9
10
The Stem Rust Pathogen 11
12
Historical Impact 13
Wheat stem rust caused by the fungus Puccinia graminis f. sp. tritici has been a 14
threat to wheat production and food security for as long as wheat has been cultivated by 15
human agrarian societies. Passages from the bible refer to rusts, and smut epidemics as 16
punishments on the Israelites from God for their sins (Chester, 1946). The festival of 17
Robigalia was celebrated annually by the Romans around 700 A.D. to pacify the rust 18
Major stem rust epidemics have occurred in all of the major wheat producing 1
countries. China experienced epidemics in 1948, 1951, 1952, and 1956 due to higher 2
than average temperatures and rainfall which led to ideal conditions for infection (Roelfs, 3
1977). 4
In the United States stem rust has affected primarily the spring wheat growing region. 5
One of the worst recent stem rust epidemics in the United States occurred in 1935 when 6
50% of the crop in Minnesota and North Dakota was lost to stem rust (Leonard, 2001). 7
8
Life Cycle 9
Puccinia graminis f. sp. tritici is a heteroecious fungus that requires two hosts, a 10
primary host and an alternate host to complete its life cycle. The life cycle consists of 11
multiple spore stages involving both monokaryotic and dikaryotic nuclear conditions. 12
The primary host of Pgt is T. aestivum and the alternate hosts are of the genus Berberis, 13
primarily common barberry (Berberis vulgaris). The sexual stage of the life cycle takes 14
place on the alternate host and asexual reproduction takes place on the primary host 15
(Leonard and Szabo, 2005). 16
Teliospores overwintering on infected straw germinate annually in conjunction 17
with the development of new growth of leaves of the barberry host (Roelfs, 1985). Each 18
teliospore consists of two cells each containing two haploid nuclei that undergo 19
arrested in diplonema during dormancy (Boehm et al., 1992). Both cells germinate to 1
produce a basidum to which the four haploid nuclei migrate upon completion of meiosis. 2
Within the basidum, the four nuclei are separated by three transverse septa. From each 3
basidum a sterigma elongates, through which the haploid nuclei migrate into the 4
developing basidiospore as it forms at the tip of the sterigma (Roelfs, 1985). In the 5
basidiospore the haploid nuclei undergo mitosis to produce two identical haploid nuclei. 6
Basidiospores are ejected and carried by air currents to the barberry host, on 7
which they infect younger leaves. The structure produced from infection is a flask 8
shaped pycnia on the adaxial leaf surface. Two gametic cells of the pycnia are involved 9
in sexual recombination between the + and – mating types. The male gametes are the 10
pycniospores which are extruded from the pycnium in a drop of nectar, making them 11
available for dissemination among pycnia by insects and rain. The female gametes are 12
flexuous hyphae that extend out of the top of the pycnium. The contact of a pycniospore 13
with the nectar of an opposite mating type induces the formation of a pyncial cap of the 14
pycniospore (Anikster, 1999). When the pycniospore contacts a flexuous hypha, fusion 15
of the cells occurs and the haploid nucleus migrates through flexuous hypha , then 16
through the monokaryotic hyphae to the cells at the base of the pycnium (Johnson and 17
Newton, 1946). The dikaryotic state is established with the division and subsequent 18
union of + and – gametes. The result of this union is the production of a dikaryotic 19
through which chains of dikaryotic aeciospores are produced which are capable of 1
infecting the wheat host (Roelfs, 1985). 2
Aeciospores infecting the wheat host produce a dense mat of hyphae below the 3
host epidermis. From these, hyphae sporophores emerge to produce dikaryotic 4
urediniospores, leading to the formation of the visible infection structure known as the 5
uredinium. Infections generally take place on the stems and leaf sheaths of the wheat 6
host. Urediniospores then re-infect wheat hosts, causing secondary infections on the 7
same plants or primary infections on other plants. As wheat host plants begin to senesce, 8
the uredinia cease uredinospore production and produce teliospores. From then on, the 9
infection structure is called a telium (Cummins and Hiratsuka, 2003). 10
The uredial stage is able to persist throughout the year on susceptible wheat 11
varieties beginning on winter wheat in the southern Great Plains moving to winter wheat 12
of the northern Great Plains and on into spring wheat of the upper Midwest. With the 13
eradication of the alternate hose, barberry (Berberis vulgaris L.), urediniospores and not 14
aeciospores have become the source of primary inoculum in the United States. Uredinia 15
that persist on winter wheat grown in Texas and Gulf Coast states produce the 16
urediniospores that act as the primary inoculum. These are carried via air currents 17
northward and eastward on what is known as the Puccina pathway to spring wheat 18
growing regions (Stakman and Lambert, 1928). 19
Infection Process 1
The infection process of the uredinial stage begins with the landing of a 2
urediniospore on a stem or leaf surface. Spore germination takes place if it is in contact 3
with a film of water. The germinating urediniospore produces a germ tube which extends 4
its growth perpendicular to the long axis of epidermal cells of the stem or leaf, thereby 5
orienting itself towards the parallel rows of stomata. 6
Migration of the germ tube takes place until a stomate is reached and appressoria 7
formation is induced. Chemical and physical stimuli are inducers of appressoria 8
formation (Read et al. 1997; Collins et al. 2001). When the specific spacing of the 9
intercellular junctions of epidermal cells adjacent to stomata is encountered by the germ 10
tube, the induction of an appressoria above the stomatal opening is induced (Read et al. 11
1997). Other chemical factors may be involved in the signaling of appressoria formation 12
including the leaf alcohols cis-3-hexen-1-ol and trans-2-hexen-1-ol (Collins et al. 2001). 13
The two nuclei of the uredeniospore migrate from the germ tube to the 14
appressorium, where they undergo mitosis and are subsequently separated from the germ 15
tube by a septum. The appressoria forces a penetration peg through the stomata and an 16
elongate substomatal vesicle develops. Another round of mitosis takes place producing a 17
hypha from one of the substomatal vesicle. A pair of nuclei then migrate into the 18
developing infection hypha. Upon contact with a host cell, the infection hypha develops 19
which contains two to four nuclei, enzymatically degrades the cell wall and causes an 1
invagination of the host cell membrance. Within the periplasmic space of the host cell, 2
the haustorium enlarges. It is through haustoria that fungal hyphae are able to extract 3
nutrients from host cells (Chong, 1985). 4
5
Physiologic Races 6
Within the classification of formae specialis of Pgt exists further subdivision of 7
the pathogen at the level of physiologic race. The differentiation of races of Pgt follow 8
observations based on the gene for gene concept of H.H. Flor, in which the resistance 9
gene in the host recognizes an avirulence target in the pathogen (Flor, 1955). The 10
development of virulence occurs when the avirulence target is modified so as to become 11
undetectable by the cognate recognition factor in the host. 12
The designation of races within Pgt is dertermined by specificities of avirulence 13
and virulence to a defined set of stem rust resistance genes present in a differential set of 14
host cultivars (Roelfs, 1988). The differential set consists of cultivars possessing single 15
dominant stem rust resistance genes to which the avirulence and virulence of a stem rust 16
isolate determines the race classification. 17
In the current nomenclature system, the presence of a high or low infection type 18
(IT) is determined for a race to four sets of genes consisting of four genes each. A letter 19
by its specific avirulence/virulence profile. A new race of Pgt can be designated upon the 1
development of a novel virulence/avirulence profile. The development of a new race of 2
Pgt in Eastern Africa and the subsequent development of novel virulences in subsequent 3
races derived from the race designated as TTKS has prompted a proposal to add a fifth 4
set of genes to the current nomenclature system (Jin and Szabo, 2008). 5
6
Population Genetics and Evolution 7
The deployment of single genes for resistance can lead to profound changes in the 8
population structure of Pgt populations. The large scale cultivation of wheat lines 9
carrying single genes for resistance deployed on a large scale places tremendous 10
directional selection pressure on stem rust pathogen populations towards the 11
predominance of pathotypes virulent to the resistance gene (Van der Plank, 1968). 12
The large scale deployment of a highly efficacious single gene effective against a 13
large fraction of the pathogen population and the subsequent evolution of the pathogen 14
population towards virulence is known as the ‘boom and bust’ cycle (Sun and Yang, 15
1999). The inefficacy of the resistance gene is not due to changes in the gene itself but to 16
the proliferation of mutants in the pathogen population with an aberrant avirulence gene. 17
These individuals are able to proliferate on hosts carrying the cognate resistance gene for 18
population, as they are the only individuals able to proliferate on the widely deployed 1
host carrying the defeated gene. 2
With the near eradication of barberry in the United States, the opportunity for 3
sexual reproduction by Pgt has been greatly minimized. Without the opportunity for 4
sexual union of mating types and recombination during meiosis, most common genotypes 5
of Pgt have adapted to strictly asexual reproduction (Zambino et al. 2000). In this 6
adaptation they have lost the ability to produce teliospores and induce recombination 7
through meiosis. Sexual recombination is no longer a principal source of genetic 8
variation in Pgt populations in US populations. In asexual reproduction, the main source 9
of variation is mutation (McDonald and Linde, 2002). From Pgt isolates collected in 10
Minnesota, the greatest diversity of races in aeciospores and urediniospores were from 11
times prior to Barberry eradication (Peterson et al., 2005). 12
13
New Highly Virulent Pgt Race 14
A race of Pgt emerged in Uganda in 1998 which was identified in 1999 as the 15
only global race to possess virulence to Sr31 (Pretorius, 2000) present on the 1BL∙1RS 16
translocation derived from ‘Petkus’ rye (Secale cereale L.) (Zeller, 1983). This 17
translocation is the source of stem rust resistance in approximately 30% of the advanced 18
lines from CIMMYT (Singh, 2008). The race originally called Ug99 was designated as 19
1988). This designation indicates the race elicits a high IT to all genes in the first two 1
sets, a low IT to Sr36 in the third set and SrTmp in the fourth set. The original race 2
TTKS has now been designated TTKSK based on the addition of a fifth set of 3
differentials (Jin and Szabo, 2008). This new race combines Sr31 virulence with 4
virulence to the majority of T. aestivum derived stem rust resistance genes (designated 5
“Sr”genes). Since its identification, new variants with additional virulence, such as 6
virulence to Sr24 (Jin et al. 2008) have been identified in Kenya. TTKS is now divided 7
into two races, TTKSK and TTKST with avirulence and virulence to Sr24, respectively 8
(Jin et al. 2008). The expanded virulence adaptation of race TTKS further increased the 9
genetic vulnerability of wheat. 10
This virulence to Sr31 in concert with virulence to most genes derived from T. 11
aestivum and virulence to Sr38 present on a translocation from T. ventricosum is unique. 12
The development of Sr24 virulence indicates the potential for the TTKS lineage to 13
develop more complex virulence as the population size increases and additional selection 14
pressures are presented in the form of resistant varieties (Singh, 2008). 15
The highlands of East Africa are ideal for the development of new races of rust 16
(Saari and Prescott, 1985). The year round cultivation of susceptible wheat varieties 17
under ideal conditions that promote disease development will hasten the spread of TTKS 18
and its variants. Emergence of the virulent strains of Pgt from the East African countries 19
peninsula to Yemen and as far east as Iran. This is the same path observed for the Yr9- 1
virulent race of stripe rust (Puccinia striiformis Westend f.sp. striiformis) that originated 2
in the East African highlands and migrated across the Middle East through West Asia to 3
East Asia (Singh, 2004). The movement of TTKS into Yemen is of particular concern, as 4
seasonal airborne trajectories present in the country regularly favor a north-easterly 5
movement of inoculum. A buildup of urediniospores in Yemen will provide a continuous 6
source of inoculum (Singh, 2008). 7
International attention and support for the development of resistant cultivars is 8
needed as TTKS may reach global dispersal due to its unique virulence to multiple 9
known and unknown resistance genes (Singh, 2006). The majority of current cultivars 10
grown on 90% or more of the acreage in the migration path are susceptible to TTKS 11
(Singh, 2006). Approximately 1 billion people reside in the predicted path of TTKS. 12
Many of the people present in this region are in countries that consume all the wheat 13
produced within their borders. World stocks of wheat are at record lows due to poor 14
harvest in the largest producing countries and higher consumption in countries 15
undergoing industrialization (Brown, 2004). These scenarios and the potential for TTKS 16
to cause widespread losses of wheat yields provide the conditions for great social unrest 17
and personal hardship. 18
Stem Rust Resistance 1 2
Sr and Avr gene interaction 3
Resistance to Pgt is conferred by genes that interact with pathogen virulence 4
genes in a gene-for-gene manner (Flor, 1955). It is in this relationship a particular stem 5
rust resistance gene present in the host is cognate to an avirulence gene in the stem rust 6
pathogen. Several hypotheses regarding the physical interaction between resistance gene 7
products and avirulence gene products (Jones and Dangl, 2006). In a recptor ligand 8
relationship, the pathogen effector molecule (avirulece gene product) interacts directly 9
with the host recognition protein (resistance gene product) (Martin et al. 2003). Another 10
well supported model is the guard hypothesis, in which the host resistance protein 11
recognizes the perturbation of another host factor by the pathogen effector (Bent and 12
Mackey, 2007). In this model the host resistance protein detects the avirulence protein 13
indirectly. It is by these relationships that a mutation in an avirulence gene leads to 14
virulence in the pathogen due to the subsequent inability of the resistance gene product to 15
detect the presence of the avr gene product and induce a defense response. 16
17
Stem Rust Resistance Genes 18
Stem rust resistance genes have been derived from T. aestivum itself, members of 19
been designated, with three gene loci having multiple alleles (McIntosh et al. 1995) and 1
other stem rust resistance genes exist with temporary designation status. 2
Several of the genes derived from wild relatives present on Robertsonian 3
translocations or small chromosomal introgression segments have been relied upon in 4
breeding programs and have been deployed commercially including Sr24, 25, 31, 36 ,38, 5
and Sr1RAmigo. Many undeployed stem rust resistance genes are present on introgression 6
segments comprising large segments or entire chromosomes. These large introgressions 7
carrying substantial amounts of alien chromatin are associated with high levels of linkage 8
drag and decreased agronomic performance. Examples of these introgressions include 9
Sr32, Sr39 and Sr40 (Singh, 2008). 10
The majority of Sr genes confer seedling resistance, which is effective in both 11
seedlings and adult plants to varying degrees. Seedling resistance genes confer a range of 12
resistance phenotypes. Several genes confer complete hypersensitive immunity 13
evidenced by the absence of any symptoms of infection or minute hypersensitive 14
flecking. Examples of genes conferring a hypersensitive phenotype include Sr5, 17, 27, 15
and 36 (Singh, 2008). 16
Several genes confer an adult plant resistance that does not entirely prevent 17
infection by Pgt but slows the development of symptoms, so as to maintain normal plant 18
function through maturity. One of the most widely utilized adult plant resistance genes is 19
with small effects comprising the “Sr2-Complex” (McIntosh, 1988). Resistance from Sr2 1
in the cultivar “Hope” and other emmer-derived resistance in the cultivar “Thatcher” 2
provided a foundation for stem rust resistance in spring wheat germplasm of the United 3
States and widely adapted lines developed by Dr. N. E. Borlaug (Hare and McIntosh, 4
1979). 5
6
Resistance to Ug99 7
The unique virulence profile of Ug99 (Pgt race TTKSK and derivatives) makes it 8
a tremendous threat to wheat production worldwide. For many years Sr31 provided 9
seemingly durable resistance globally but inevitably selection pressures led to the 10
development of virulence in Ug99. Developing lines with adequate and durable 11
resistance to Ug99 has presented unique and challenging problem to wheat scientists 12
worldwide with the majority of genes conferring resistance coming from wild relatives. 13
Many of the effective resistance genes are present on large translocations and are 14
associated with linkage drag. 15
Field evaluations in Kenya and greenhouse evaluations at the USDA Cereal 16
Disease Lab have elucidated Sr genes effective against Ug99 (Jin et al. 2007). These 17
include Sr13, 22, 24, 25, 26, 27, 28, 32, 33, 35, 36, 37, 39, 40, 44, Sr Tmp, and Tt-3 and 18
genes, Sr24, Sr26, Sr36 and Sr1RSAmigo have been deployed in wheat cultivars in some 1
countries. 2
Virulence exists in other races endemic to particular growing regions of the world 3
to seven of the Sr genes effective against Ug99, including SrTmp, 13, 36, 24, 27, 29 and 4
Sr1RSAmigo (Singh et al., 2008). Virulence to the remaining effective genes does not exist 5
for several reasons. Some have proven durable over time despite deployment singly over 6
large acreages. More commonly, many Sr genes including Sr32, 37, 39, 40 and 44, have 7
not been deployed in modern cultivars due to their presence on large alien translocations 8
that have deleterious effects on important agronomic characteristics. 9
10
Development of Resistant Germplasm 11
Reducing the amount of alien chromatin associated with undeployed Sr genes 12
effective against Ug99 is necessary to ensure lines carrying the genes are able to meet the 13
needed annual increase in yields to meet growing demands. One example of yield 14
depression due to alien chromatin is in CIMMYT attempts to introgress Lr35 linked to 15
Sr39, resulting in a 15-20% yield detriment (Singh, unpublished data). In the original 16
release of germplasm carrying Sr26, a 10% yield penalty was associated with the gene 17
located on a Thinopyron elongatum translocation segment on chromosome 6DL (Friebe 18
Methods involved in the introgression of alien chromosome segments can be 1
employed in reducing the size of alien chromosome segments carrying resistance to 2
Ug99. Homoeologous recombination between alien chromosome segments and wheat 3
chromosomes can be induced using ph1b and PhI lines to generate critical recombinants, 4
followed by subsequent backcrosses to recover the majority of the recurrent parent 5
genome (Qi et al., 2007). 6
Diagnostic molecular markers linked to effective resistance genes are available 7
for Sr24, Sr26, Sr36 and Sr1RSAmigo, each of which was transferred to wheat from non- 8
homologuous or partially homologous genomes (Saal and Wricke, 1999; Mago et al., 9
2004; Tsilo et al., 2008). The ability to detect the presence of specific stem rust resistance 10
genes using molecular markers presents a viable method for identifying resistance to 11
TTKS in the absence of the pathogen itself. Markers will also facilitate the identification 12
of recombinant lines carrying reduced translocation segments. 13
Benefits of durability of resistance can be realized from the pyramiding of Sr 14
genes effective against TTKS. The hypothesis of the efficacy of resistance gene 15
pyramids is based on the low probability of a pathogen developing virulence to two or 16
more genes simultaneously. Multiple mutations to virulence must also not be 17
accompanied by any significant reduction in pathogen fitness (Ayliffe et al., 2008). 18
can potentially lead to residual effects of pyramiding defeated Sr genes (Ahmed et al., 1
1997; Kousik and Richie, 1998). 2
Several of the difficulties of developing lines with resistance to TTKS can be 3
circumvented with the use of molecular markers. Marker loci linked to effective Sr genes 4
can be used for selection in breeding populations. Screening for resistance genes with a 5
molecular assay allows for the screening of greater numbers in less time than using 6
phenotypic greenhouse assays or field evaluations. Molecular markers can aide in the 7
identification of lines currently carrying resistance to Ug99 and will be useful in the 8
future development of resistant cultivars. 9
Pyramiding multiple Sr genes in a single line utilizing specific 10
avirulence/virulence specificities of Pgt isolates can be complicated and time consuming 11
and nearly impossible for undeployed genes to which no virulence exists. The 12
pyramiding of multiple resistance genes in a single line becomes possible using 13
molecular markers by selecting for loci linked to the genes of interest. Pyramiding 14
multiple genes in linkage blocks in coupling phase becomes possible with sufficient 15
population sizes and the identification of critical recombinants. With the improved 16
capability of deploying resistance gene pyramids, the durability of resistance to Ug99 17
will be greatly enhanced. 18
References 1
Ahmed H.U., Finckh M.R., Alfonso R.F., Mundt C.C. (1997) Epidemiological effect of 2
gene deployment strategies on bacterial blight of rice. Phytopathology 87:66-70. 3
4
Anikster Y., Eilam T., Mittelman L., Szabo L.J., Bushnell W.R. (1999) Pycnial nectar of 5
rust fungi induces cap formation on pycniospores of opposite mating type. 6
Mycologia 91:858-870. 7
8
Ayliffe M., Singh R., Lagudah E. (2008) Durable resistance to wheat stem rust needed. 9
Current Opinion in Plant Biology 11:187-192. 10
11
Aquino P., Carrion, F., and Calvo, R. (2002) Selected wheat statistics. In “CIMMYT 12
2000-2001 World Wheat Overview and Outlook: Developing No-Till Packages 13
for Small Scale Farmers” (J.Ekboir, ed.) pp. 52-56. CIMMYT, Mexico D.F. 14
15
Badaeva E.D., Dedkova O.S., Gay G., Pukhalskyi V.A., Zelenin A.V., Bernard S., 16
Bernard M. (2007) Chromosomal rearrangements in wheat: their types and 17
distribution. Genome 50:907-926. 18
19
10:204-222. 1 2
Bent A.F., Mackey D. (2007) Elicitors, effectors, and R genes: The new paradigm and a 3
lifetime supply of questions. Annual Review of Phytopathology 45:399-436. 4
5
Binford L.R. (1968) Post-Pleistocene Adaptations. In New Perspectives in Archaeology 6
(Binford S.R. and Binford L.R., eds.) pp. 313-341. Aldine, Chicago. 7
8
Boehm E.W.A., Wenstrom J.C., McLaughlin D.J., Szabo L.J., Roelfs A.P., Bushnell 9
W.R. (1992) An ultrastructural pachytene karyotype for Puccinia graminis fsp. 10
tritici. Canadian Journal of Botany-Revue Canadienne De Botanique 70:401-413. 11
12
Bonafede M., Kong L.R., Tranquilli G., Ohm H., Dubcovsky J. (2007) Reduction of a 13
Triticum monococcum chromosome segment carrying the softness genes Pina and 14
Pinb translocated to bread wheat. Crop Science 47:821-828. 15
16
Bonjean A.P., Angus W.J. (2001) The world wheat book: a history of wheat breeding 17
Intercept, London; Paris. 18
19
water tables and rising temperatures. Earth Policy Institute. W.W. Norton & Co., 1
New York. 2
3
Chantret N., Salse J., Sabot F., Rahman S., Bellec A., Laubin B., Dubois I., Dossat C., 4
Sourdille P., Joudrier P., Gautier M.F., Cattolico L., Beckert M., Aubourg S., 5
Weissenbach J., Caboche M., Bernard M., Leroy P., Chalhoub B. (2005) 6
Molecular basis of evolutionary events that shaped the hardness locus in diploid 7
and polyploid wheat species (Triticum and Aegilops). Plant Cell 17:1033-1045. 8
9
Chester K.S. (1946) The nature and prevention of the cereal rusts as exemplified in the 10
leaf rust of wheat Chronica Botanica Co., Waltham, Mass.,. 11
12
Chong J., Harder D.E., Rohringer R. (1985) Cytochemical studies on Puccinia graminis 13
fsp tritici in a compatible wheat host. 1. Walls of intercellular hyphal cells and 14
haustorium mother cells. Canadian Journal of Botany-Revue Canadienne De 15
Botanique 63:1713-1724. 16
17
Collins T.J., Moerschbacher B.M., Read N.D. (2001) Synergistic induction of wheat stem 18
rust appressoria by chemical and topographical signals. Physiological and 19
1
Cummins G.B., Hiratsuka Y. (2003) Illustrated genera of rust fungi. 3rd ed. American 2
Phytopathological Society, St. Paul, Minn. 3
4
Diamond J. (2002) Evolution, consequences and future of plant and animal 5
domestication. Nature 418:700-707. 6
7
Endo T.R. (1988) Induction of chromosomal structural changes by a chromosome of 8
Aegilops cylindrical in common wheat. Journal of Heredity 79:366-370. 9
10
Endo T.R. (1990) Gametocidal chromosomes and their induction of chromosome 11
mutations in wheat. Japanese Journal of Genetics 65:135-152. 12
13
Endo T.R., Gill B.S. (1996) The deletion stocks of common wheat. Journal of Heredity. 14
87:295-307. 15
16
Endo T.R. (2007) The gametocidal chromosome as a tool for chromosome manipulation 17
in wheat. Chromosome Research 15:67-75. 18
19
of low-copy DNA sequences in polyploid wheat: A possible mechanism for 1
differentiation of homoeologous chromosomes. Genetics 147:1381-1387. 2
3
Flor H.H. (1955) Host-parasite interaction in flax rust - its genetics and other 4
implications. Phytopathology 45:680-685. 5
6
Friebe B., Jiang J., Knott D.R., Gill B.S. (1994) Compensation indexes of radiation 7
induced wheat agropyron-elongatum translocations conferring resistance to leaf 8
rust and stem rust. Crop Science 34:400-404. 9
10
Gill K.S., Gill B.S. (1991) A DNA fragment mapped within the submicroscopic deletion 11
of Ph1, a chromosome-pairing regulator gene in polyploid wheat. Genetics 12
129:257-259. 13
14
Giorgi, B. (1978) A homoeolgous pairing mutant isolated in Triticum 15
durum cv. Cappelli. Mutation Breeding News 11:4–5. 16
17
Hare R.A., McIntosh R.A. (1979) Genetic and cytogenetic studies of durable adult-plant 18
resistances in Hope and related cultivars to wheat rusts. Zeitschrift Fur 19
1
Harlan J.R. (1975) Crops & man. American Society of Agronomy, Madison, Wis. 2
3
Huang S., Sirikhachornkit A., Su X.J., Faris J., Gill B., Haselkorn R., Gornicki P. (2002) 4
Genes encoding plastid acetyl-CoA carboxylase and 3-phosphoglycerate kinase of 5
the Triticum/Aegilops complex and the evolutionary history of polyploid wheat. 6
Proceedings of the National Academy of Sciences of the United States of America 7
99:8133-8138. 8
9
Jin Y., Szabo L.J., Pretorius Z.A., Singh R.P., Ward R., Fetch T. (2008) Detection of 10
virulence to resistance gene Sr24 within race TTKS of Puccinia graminis fsp. 11
tritici. Plant Disease 92:923-926. 12
13
Jin Y., Singh R.P., Ward R.W., Wanyera R., Kinyua M., Njau P., Pretorius Z.A. (2007) 14
Characterization of seedling infection types and adult plant infection responses of 15
monogenic Sr gene lines to race TTKS of Puccinia graminis fsp. tritici. Plant 16
Disease 91:1096-1099. 17
18
Johnson T., Newton M. (1946) Specialization, hybridization, and mutation in the cereal 19
1
Jones J.D.G., Dangl J.L. (2006) The plant immune system. Nature 444:323-329. 2
3
Kashkush K., Feldman M., Levy A.A. (2002) Gene loss, silencing and activation in a 4
newly synthesized wheat allotetraploid. Genetics 160:1651-1659. 5
6
Kashkush K., Feldman M., Levy A.A. (2003) Transcriptional activation of 7
retrotransposons alters the expression of adjacent genes in wheat. Nature Genetics 8
33:102-106 9
10
Kousik C.S., Ritchie D.E. (1999) Development of bacterial spot on near-isogenic lines of 11
bell pepper carrying gene pyramids composed of defeated major resistance genes. 12
Phytopathology 89:1066-1072. 13
14
Kuraparthy V., Chhuneja P., Dhaliwal H.S., Kaur S., Bowden R.L., Gill B.S. (2007) 15
Characterization and mapping of cryptic alien introgression from Aegilops 16
geniculata with new leaf rust and stripe rust resistance genes Lr57 and Yr40 in 17
wheat. Theoretical and Applied Genetics 114:1379-1389. 18
19
Enemy to Modern Foe. (Peterson, P.D. ed.) pp. 1–11. The American 1
Phytopathological Society, St. Paul MN. 2
3
Leonard K.J., Szabo L.J. (2005) Pathogen profile. Stem rust of small grains and grasses 4
caused by Puccinia graminis. Molecular Plant Pathology 6:489-489. 5
6
Levy A.A., Feldman M. (2004) Genetic and epigenetic reprogramming of the wheat 7
genome upon allopolyploidization. Biological Journal of the Linnean Society 8
82:607-613. 9
10
Liu B., Segal G., Vega J.M., Feldman M., Abbo S. (1997) Isolation and characterization 11
of chromosome-specific DNA sequences from a chromosome arm genomic 12
library of common wheat. Plant Journal 11:959-965. 13
14
Lukaszewski A.J. (1990) Frequency of 1RS∙AL and 1RS∙1BL translocations in united- 15
states wheats. Crop Science 30:1151-1153. 16
17
Lukaszewski A.J. (2000) Manipulation of the 1RS∙1BL translocation in wheat by induced 18
homoeologous recombination. Crop Science 40:216-225. 19
Maan S.S. (1975) Exclusive preferential transmission of an alien chromosome in 1
common wheat. Crop Science 15:287-292. 2
3
Mago R., Bariana H.S., Dundas I.S., Spielmeyer W., Lawrence G.J., Pryor A.J., Elllis 4
J.G. (2005) Development of PCR markers for the selection of wheat stem rust 5
resistance genes Sr24 and Sr26 in diverse wheat germplasm. Theoretical and 6
Applied Genetics 111:496-504. 7
8
Martin G.B., Bogdanove A.J., Sessa G. (2003) Understanding the functions of plant 9
disease resistance proteins. Annual Review of Plant Biology 54:23-61. 10
11
Masoudi-Nejad A., Nasuda S., McIntosh R.A., Endo T.R. (2002) Transfer of rye 12
chromosome segments to wheat by a gametocidal system. Chromosome Research 13
10:349-357. 14
15
McDonald B.A., Linde C. (2002) The population genetics of plant pathogens and 16
breeding strategies for durable resistance. Euphytica 124:163-180. 17
18
McFadden, E.S. (1930) A successful transfer of emmer characteristics to vulgare wheat. 19
1
McIntosh R.A., Wellings C.R., Park R.F. (1995) Wheat rusts : an atlas of resistance genes 2
CSIRO ; Kluwer Academic Publishers, East Melbourne, Australia 3
Dordrecht ; Boston. 4
5
Meng E., Longmire J., Moldashev A. (2000) Kazakhstan's wheat system: priorities, 6
constraints, and future prospects. Food Policy 25:701-717. 7
8
Naranjo T. (1990) Chromosome structure of durum wheat. Theoretical and Applied 9
Genetics 79:397-400. 10
11
Payne P.I., Holt L.M., Lawrence G.J., Law, C.N. (1982) The genetics of gliadin and 12
glutenin, the major storage proteins of the wheat endosperm. Qualitas Plantarum 13
Plant Foods for Human Nutrition. 31:229-241. 14
15
Peterson P.D. (2001) Stem rust of wheat : From ancient enemy to modern foe. APS Press, 16
St. Paul, Minn. 17
18
Peterson P.D., Leonard K.J., Roelfs A.P., Sutton T.B. (2005) Effect of barberry 19
Disease 89:935-940. 1 2
Porter J.R., Jameison P.D., andGrace P.R. (2007) Wheat Production and Global Climate 3
Change. In: Terrestrial Ecosystems in a Changing World. Springer Berlin, 4
Heidelberg. 5
6
Pretorius Z.A., Singh R.P., Wagoire W.W., Payne T.S. (2000) Detection of virulence to 7
wheat stem rust resistance gene Sr31 in Puccinia graminis fsp. tritici in Uganda. 8
Plant Disease 84:203. 9
10
Qi L.L., Friebe B., Zhang P., Gill B.S. (2007) Homoeologous recombination, 11
chromosome engineering and crop improvement. Chromosome Research 15:3-19. 12
13
Read N.D., Kellock L.J., Collins T.J., Gundlach A.M. (1997) Role of topography sensing 14
for infection-structure differentiation in cereal rust fungi. Planta 202:163-170. 15
16
Riley R., Chapman V. (1958) Genetic control of the cytologically diploid behaviour of 17
hexaploid wheat. Nature 182:713-715. 18
19
like meiotic behaviour of wheat. Nature 186:259-260. 1 2
Riley R., Chapman V., Johnson R. (1968) Incorporation of alien disease resistance in 3
wheat by genetic interference with regulation of meiotic chromosome synapsis. 4
Genetical Research 12:199 5
6
Roelfs A.P. (1977) Foliar fungal diseases of wheat in peoples republic of china. Plant 7
Disease Reporter 61:836-841. 8
Roelfs, A.P. (1985) Wheat and rye stem rust. In: The Cereal Rusts, Vol. 2 (Roelfs, A.P., 9
and Bushnell, W.R. eds.) pp. 3-37. Academic Press, Orlando, FL. 10
11
Roelfs A.P., Martens J.W. (1988) An international system of nomenclature for Puccinia 12
graminis fsp. tritici. Phytopathology 78:526-533. 13
14
Saari E.E., Prescott J.M. (1985) World distribution in relation to economic losses. In: The 15
Cereal Rusts, Vol. 2 (Roelfs, A.P.,and Bushnell, W.R. eds.) pp. 3-37. Academic 16
Press, Orlando, FL. 17
18
Saal B., Wricke G. (1999) Development of simple sequence repeat markers in rye (Secale 19
1
Schneider A., Molnar I., Molnar-Lang M. (2008) Utilisation of Aegilops (goatgrass) 2
species to widen the genetic diversity of cultivated wheat. Euphytica 163:1-19. 3
4
Sears, E.R., (1956) The transfer of leaf rust resistance from Aegilops umbellulata to 5
wheat. Brookhaven Symposium on Biology 9: 1-12. 6
7
Sears E.R., Steinitzsears, L.M. (1957) Ultraviolet and x-ray induced chromosomal 8
aberrations in wheat. Genetics 42:623-630. 9
10
Sears E.R. (1972) Agropyron-wheat transfers through induced homoeologous pairing. 11
Canadian Journal of Genetics and Cytology 14:736. 12
13
Sears E.R. (1977) Transferring genes to wheat chromosomes from those of its wild 14
relatives. Cereal Foods World 22:477-477. 15
16
Shaked H., Kashkush K., Ozkan H., Feldman M., Levy A.A. (2001) Sequence 17
elimination and cytosine methylation are rapid and reproducible responses of the 18
1759. 1 2
Singh R., Huerta-Espino J., Rosewarne G. (2004) Wheat Rust In Asia: Meeting the 3
Challenges with Old and New Technologies, International Crop Science 4
Congress, ISBN, Brisbane, Australia. 5
6
Singh R., Hodson D., Jin Y., Huerta-Espino J., Kinyua M., Wanyera R., Njau P., Ward R. 7
(2006) Current status, likely migration and strategies to mitigate the threat to 8
wheat production from race Ug99 (TTKS) of stem rust pathogen. CAB Reviews: 9
Perspectives in Agriculture, Veterinary Science, Nutrition and Natural Resources 10
1. 11
Singh R.P., Hodson D.P., Huerta-Espino Jin Y., Njau P., Wanyera R., Herrera-Foessel 12
S.A., Ward R.W. (2008) Will stem rust destroy the world's wheat crop?, p. 271 13
309 Advances in Agronomy, Vol 98, Vol. 98. Elsevier Academic Press Inc, 14
San Diego. 15
16
Stakman E.C., Lambert E.B. (1928) The relation of temperature during the growing 17
season in the spring wheat area of the United States to the occurrence of stem rust 18
epidemics. Phytopathology 18:369-374. 19
Sun P., and Yang X.B. (1999) Two properties of a gene-for-gene co-evolution system 1
under human perturbations. Pytopathology. 89:911-816. 2
3
Van der Plank J.E. (1968) Disease resistance in plants Academic Press, New York,. 4
5
Tsilo, T.J., Y. Jin, Anderson J.A. (2008) Diagnostic microsatellite markers for the 6
detection of stem rust resistance gene Sr36 in diverse genetic backgrounds of 7
wheat. Crop Sci. 48:253-261. 8
9
Villareal R.L., Rajaram S., Mujeebkazi A., Deltoro E. (1991) The effect of chromosome 10
1B/1R translocation on the yield potential of certain spring wheats (Triticum 11
aestivum L.). Plant Breeding 106:77-81. 12
Vocke G., Alan E. 2008 Wheat Outlook, A report from the USDA-Economic Research 13
Service. 14
15
Wanyera R., Kinyua M.G., Jin Y., Singh R.P. (2006) The spread of stem rust caused by 16
Puccinia graminis fsp. tritici, with virulence on Sr31 in wheat in Eastern Africa. 17
Plant Disease 90:113-113. 18
19
genes to DNA markers in the rust fungus Puccinia graminis. Phytopathology 1
90:819-826. 2
3
Zeller F.J., Fuchs E. (1983) Cytology and disease resistance of a 1A1R and some 1B1R 4
wheat-rye translocation cultivars. Zeitschrift Fur Pflanzenzuchtung-Journal of 5
Plant Breeding 90:285-296. 6
7
Zhang W.J., Lukaszewski A.J., Kolmer J., Soria M.A., Goyal S., Dubcovsky J. (2005) 8
Molecular characterization of durum and common wheat recombinant lines 9
carrying leaf rust resistance (Lr19) and yellow pigment (Y) genes from 10
Lophopyrum ponticum. Theoretical and Applied Genetics 111:573-582. 11
12
Zohary D., Hopf M. (1993) Domestication of plants in the Old World : the origin and 13
spread of cultivated plants in West Asia, Europe, and the Nile Valley. 2nd ed. 14
Clarendon Press; Oxford University Press, Oxford New York. 15
Chapter 2 1
Genotyping of U.S. Wheat Germplasm for Presence of Stem Rust Resistance Genes 2
Sr24, Sr36 and Sr1RSAmigo 3
4 5 6
Submitted to Crop Science 7
Genotyping of U.S. Wheat Germplasm for Presence of Stem Rust Resistance Genes Sr24, 1
Sr36 and Sr1RSAmigo 2
3
Eric L. Olson, Gina Brown-Guedira*, David S. Marshall, Yue Jin, Mohamed Mergoum, 4
Iago Lowe, and Jorge Dubcovsky 5
6
E.L. Olson, Department of Crop Science, North Carolina State University, Campus Box 7
7620, Raleigh, NC 27695-7620; G. Brown-Guedira, USDA-ARS Plant Science Research, 8
Department of Crop Science, North Carolina State University, Campus Box 7620, 9
Raleigh, NC 27695-7620; D.M. Marshall, USDA-ARS Plant Science Research, 10
Department of Plant Pathology, North Carolina State University, Campus Box 7616, 11
Raleigh, NC 27695-7616. Y. Jin, USDA-ARS, Cereal Disease Lab., 1551 Lindig St, 12
Univ. of Minnesota, St. Paul, MN 55108; M. Mergoum, Dep. of Plant Sciences, 270C 13
Loftsgard Hall 270C, North Dakota State University, Fargo, ND 58105-5051; I. Lowe 14
and J. Dubcovsky, Dep. of Plant Sciences, Univ. of California, Davis, CA 95616-8780 15
16
*Corresponding author E-mail: [email protected] 17
Mention of trade names or commercial products in this article is solely for the purpose of 1
providing specific information and does not imply recommendation or endorsement by 2
the U.S. Department of Agriculture. 3
4
Abbreviations: Pgt, Puccinia graminis f.sp. tritici; IT, infection type; SWW, soft winter 5
wheat; HWW, hard winter wheat; HSW, hard spring wheat; SRW, soft red winter wheat; 6
HRW, hard red winter wheat; HRS, hard red spring wheat; bp, base pairs; SSR, simple 7
sequence repeat; STS, sequence tagged site; PCR, polymerase chain reaction 8