JOURNAL OFCLINICALMICROBIOLOGY, Mar. 2004, p. 1265–1269 Vol. 42, No. 3
0095-1137/04/$08.00⫹0 DOI: 10.1128/JCM.42.3.1265–1269.2004
Copyright © 2004, American Society for Microbiology. All Rights Reserved.
Comparative Analysis of
agr
Locus Diversification and Overall Genetic
Variability among Bovine and Human
Staphylococcus aureus
Isolates
Philippe Gilot
1* and Willem van Leeuwen
2Unite´ de Pathologie Infectieuse et Immunologie, Institut National de la Recherche Agronomique, F-37380
Nouzilly, France,1and Erasmus MC, Rotterdam, The Netherlands2
Received 9 July 2003/Returned for modification 23 September 2003/Accepted 26 November 2003
The accessory gene regulator (agr) is a central system that controls the expression ofStaphylococcus aureus
virulence factors. In this study, the distribution of agr alleles, defined by agr restriction fragment length
polymorphism withinagrinterference groups, among S. aureusisolates from bovine and human origin was
measured with PCR-based techniques. Statistically highly significant associations of someagralleles with the
infection of a specific host were found. The genetic difference between the twoS. aureuspopulations was further
stressed after analysis of their genetic background by binary typing.
Staphylococcus aureusis an important cause of a variety of
diseases in humans and animals and is a large economic prob-lem worldwide (9, 14). Whereas the mechanism of pathoge-nicity ofS. aureusis still not completely understood, the bac-terium is hypothesized to invade the tissue by starting to adhere directly to epithelial cells (13, 18). The expression of such adhesive proteins, which recognize macromolecules of the host tissue, is globally controlled by theagrlocus (17). Theagr
operon encodes a two-component signaling system, which is driven by a quorum-sensing autoinducing peptide (AIP), en-coded by theagrDgene.S. aureusisolates can be classified in four majoragrinterference groups on the basis of AIP speci-ficity for its signal receptor (AgrC) (4, 6, 7). In a laboratory setting, Ji et al. showed that AIP from strains belonging to a specificagrgroup activate each other’sagrresponse, whereas AIP from strains from differentagrgroups compete with each other at the level ofagrexpression, since each AIP inhibits the
agr response in strains from other groups (7). This type of bacterial interference is unusual, because it affects the expres-sion of a subset of genes rather than inhibiting growth. Diverse works have noted the existence of distinctagralleles, as defined withagrrestriction fragment length polymorphism, withinagr
groups both in clinicalS. aureusisolates that originated with humans and in those from cows (2, 15, 16, 19, 24, 29). In a recent study, one of us identified 12 distinct agralleles in an epidemiologically unrelated collection of bovine mastitis iso-lates. The majority of strains from this collection are neverthe-less represented by one particularagrallele (R III A1) fromagr
group 1 (2). These observations suggest the occurrence of host-adapted (or tissue-adapted)S. aureusstrains in which the
agrrestriction type (allele) may play a significant role. In this study, we compared the distribution ofagralleles in relation to the genotype amongS. aureusstrains of human and bovine origin by using restriction endonuclease-PCR of theagr
locus (agrRE-PCR) and binary typing (BT).
agrpolymorphism.Two hundred fifty-one humanS. aureus
isolates, originating in diverse collections, were analyzed byagr
RE-PCR. The results were compared with those obtained from a collection of bovine mastitis isolates, previously analyzed with the same method (Table 1). In brief, a 1,070-bp variable region of theagr operon (a sequence encodingagrD, the N-terminal two-thirds ofagrC, and the C-terminal part ofagrB) was amplified by PCR, as described previously (2). PCR am-plifications of all tested human isolates resulted in amplicons of the expected molecular weight. After restriction withAluI or
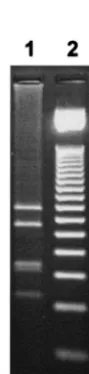
RsaI, these amplicons were electrophoresed through a 3% agarose gel, giving nine (A1, A2, A4, A5, A7 to A10, and A13) and five (R I⬘and R III to R VI) different profiles, respectively. Most of these patterns were previously identified after restric-tion of the same region of theagr locus ofS. aureus bovine mastitis isolates (2). Three of them (A9, A10, and A13) were uniquely found in human isolates. Patterns A9 (502, 244, 177, and 149 bp) and A10 (772, 144, 74, 73, and 11 bp) correspond to restriction profiles of theagrlocus fromagrgroup reference strains and from strains whose genomes were sequenced, all isolated from humans (A9:agrgroup 2 reference strain with GenBank accession no AF001782, strains N315 and Mu50; A10:agrgroup 3 reference strain with GenBank accession no AF001783, strain MRSA-252 [2]). Pattern A13 (338, 274, 168, 155, 113, and 25 bp) was newly identified during this study (Fig. 1).
The combination of theAluI andRsaI restriction patterns allows the construction of restriction maps and the definition ofagrrestriction types (alleles) (2). Elevenagrrestriction types were defined among the strains isolated from humans (Fig. 2). Two of these types, type R I⬘-A4 (a R I⬘-A1 type which has lost theAluI site at position 437) and type R I⬘-A13 (a R I⬘-A1 type which has lost theAluI site at position 890) were newly iden-tified during this study. agr restriction types were previously classified into an evolutionary scheme, which allows them to be placed in one of the fouragrinterference groups (2). By this kind of analysis, we found that the majority of strains isolated from humans fall into the classificationagr group 1 (61.7%) and group 2 (33.5%). Group 3 contains only 4.8% of the analyzed strains, and no group 4 strains were identified in our collection of strains isolated from humans. The above agr
* Corresponding author. Present address: Unite´ Bio-Agresseurs-Sante´-Environnement, Institut National de la Recherche Agrono-mique, 37380 Nouzilly, France. Phone: 33-2-47 42 75 52. Fax: 33-2-47 42 77 74. E-mail: [email protected].
1265
on May 15, 2020 by guest
http://jcm.asm.org/
group classification of strains was confirmed by anagr group-specific multiplex PCR (2). The repartition of human isolates in theagrinterference group is quite similar to what was found for bovine mastitis isolates (69%agrgroup 1 strains, 23.9%agr
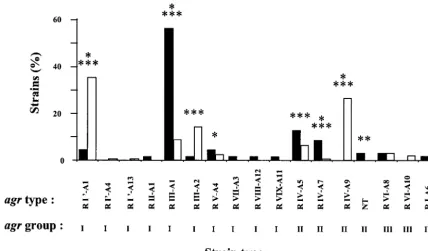
group 2 strains, 2.8%agrgroup 3 strains, and 1.4%agrgroup 4 strains) (2). Whereas the repartition of strains inagrgroups is quite similar in both populations, the 17 differentagrtypes identified are very differently distributed in each of them. The most striking differences (statistically highly significant) were found in type R I⬘-A1 (35.4% of strains of human origin but only 4.2% of strains of bovine origin), type R IV-A9 (26.7% of strains of human origin but 0% of strains of bovine origin), type R III-A1 (8.8% of strains of human origin but 56.3% of strains of bovine origin), and type R IV-A7 (0.4% of strains of human origin but 8.4% of strains of bovine origin). Some types, containing only a few strains, were also found to be unique either to the population of human origin or to the population of bovine origin (Fig. 2).
The distribution ofagrrestriction types suggests that the two populations are different and indicates that some types prob-ably expand in one or the other population due to their pos-session of unique genetic characteristics.
BT.The genetic difference between the S. aureus popula-tions of bovine origin and those of human origin was analyzed in more detail by BT, a procedure involving strain-differenti-ating DNA probes which were initially identified on the basis of RAPD analysis of a large collection of S. aureus strains isolated from humans (28). The method was proved to be able to elucidate overall genetic polymorphisms and clonal related-ness among S. aureus strains and was extensively described elsewhere (30). In brief, 12 cloned strain-specific DNA frag-ments (AW1 to AW9, AW11, AW14, and AW15) were used as probes and were hybridized to total genomic DNA of the 71 strains isolated from cows with mastitis previously analyzed by
agrRE-PCR. Hybridization of the probes was scored with a 1 or a 0 according to the presence or absence of a hybridization signal, resulting in a 12-digit binary code for each S. aureus
isolate.
All bovine mastitis isolates but two were typeable by the BT method. All typeable isolates were found to hybridize with at least 3 and up to 11 of the tested probes. Probe AW9 hybrid-ized to all typeable bovine mastitis isolates and, as suggested by a previous work, did not contribute to the discrimination power of the method for strains of bovine origin (31). BT was found to be very discriminatory and allowed the division of the collection of bovine mastitis isolates into 33 different binary types. Allagrtypes comprising more than one strain could be subdivided with BT. The highly prevalent R III-A1 type was divided into 15 (16 if nontypeable strains were included) dif-ferent binary types, whereas the following other two prevalent types, R IV-A5 and R IV-A7, were subdivided into eight and four BT types, respectively. The frequency of most BT types is only one or two strains, except for six BT types containing nine, eight, seven, five, three, and three strains (Fig. 3).
The binary types of the strains isolated from cows with mas-titis were compared to the binary types of 50 strains isolated from humans, picked randomly among all strain collections analyzed by RE-PCR of theagr locus (28). These randomly selected strains represent all major agr types as mentioned before (20 strains of the R IV-A9 type, 12 strains of the R I⬘-A1 type, 6 strains of the R III-A2 type, 5 strains of the R III-A1 type, 3 strains of the R IV-A5 type, 2 strains of the R VI-A8 type, and 2 strains of the R V-A4 type). All of theagr
[image:2.603.46.542.82.175.2]types found among these human isolates were also subdivided by BT. The prevalent R I⬘-A1 and R IV-A9 types were divided into 11 and 16 BT types, respectively. Types R III-A2, R III-A1, R IV-A5, R VI-A8, and R V-A4 contain six, five, three, FIG. 1. Identification of a novel restriction pattern in theagr
[image:2.603.141.186.492.680.2]vari-able region. The amplified varivari-able region of the agr operon was digested with AluI and electrophoresed on a 3% agarose gel. The newly identified type A13 is shown in lane 1. Molecular weight markers (50-bp DNA ladder; Promega) are shown in lane 2.
TABLE 1. Strains analyzed in this study
Collection Geographic origina No. of strains Origin Descriptionb Reference
1 Worldwide 55 Human MRSA strains 10
2 New York City Hospital 64 Human MRSA outbreak 27
3 USA 49 Human Multicenter collection of MRSA
and MSSA strains 25
4 Denmark 34 Human Nasal carrier 26
5 The Netherlands 30 Human MRSA 28
19 Human MSSA
6 France (mainly), USA, UK, Japan 71 Bovine Mastitis isolates 2
aUSA, United States; UK, United Kingdom.
bMRSA, methicillin-resistantS. aureus; MSSA, methicillin-susceptibleS. aureus.
on May 15, 2020 by guest
http://jcm.asm.org/
two, and two different BT types, respectively (Fig. 3). No bi-nary type common to strains of human and bovine origin was found.
Hierarchical clustering.Genetic relatedness between strains
was analyzed by hierarchical clustering of binary types using the squared Euclidian distance of absolute value (weights of 0 and 1 were identical), and a dendrogram was constructed (SPSS 9.0 program for Windows software) (Fig. 3). The overall clonality of all strains (except for one) based on BT results was shown by their clustering into one group for a relative genetic similarity of 79%. At a genetic similarity of 83%, three clusters, which could be linked to host specificity, were defined (Fig. 3, roman numerals). Two human isolates (RIVM IV and K2-02) do not belong to any these three clusters. The majority (84%) of the strains isolated from humans were found in clusters I and III. Both clusters also contained four strains of bovine origin (strains 125 and 130 in cluster I; strains 23 and 57 in cluster III). One of us previously reported that strains 125 and 130, isolated in Japan by Takeuchi et al., harbor a distinctagr
locus. The receptor-encoding genes (agrC) of strains 125 and 130 are highly similar to those of the agr group 1 strains, whereas their propeptide-encoding genes (agrD) are highly similar to those of groups 2 and 3, respectively. Both strains are classified in the agr group 1 by the restriction map analysis method but either in group 2 (strain 125) or in group 3 (strain 130) by the multiplex PCR method. Since the propeptide and its receptor belong to different interference groups, these two strains should be impaired in the activation of RNA III by the
agrsystem. These strains are probably in the process of
evolu-tionary development, moving fromagrgroup 1 to groups 2 and 3, respectively (2). Cluster II contains 92% of the strains of bovine origin and also includes six human isolates.
Identical to theagrtyping results, hierarchical clustering of binary types confirms that the strain populations of bovine origin and those of human origin are genetically different. Bacterial host specificity was also described by other studies using different typing techniques to analyze the genetic back-ground ofS. aureuspopulations (8, 31). Diverse studies showed divergent data for prevalence of S. aureusstrains within the differentagr interference groups (5, 12, 15, 20–22), while we showed that the agr interference group I has spread at the expense of the other agr groups, both in the population of strains isolated from cows as well as in the population of strains isolated from humans. When allelic variations within agr
groups were analyzed, a more precise scheme emerged; human isolates with the allelic variant R I⬘-A1 and R IV-A9 and bovine isolates with the allelic variant R III-A1 were predom-inant (Fig. 2). If the allelic variants R I⬘-A1 and R III-A1 belong toagr group 1 and are not very divergent (they were differentiated by the presence or absence of one restriction site), the RIV-A9 variant belongs toagrinterference group 2 (2). This indicates that in the human population, not only has
agrinterference group 1 spread at the expense of the otheragr
groups, but also one particular allelic variant of theagrgroup 2 has spread at the expense of other groups. BT, detecting DNA elements with a relative high evolutionary clock speed, reflects changes arising in other parts of the genome of each of theseagrrestriction types. Both methods produced results con-FIG. 2. Distribution ofS. aureusstrains isolated from cows with mastitis and from human patients.agrgroups andagrtypes of 71 strains isolated from cows with mastitis (solid bars) and from 251 strains of humans patients (open bars) were determined and used to subdivide the populations. NT indicates strains not typeable by the restriction endonuclease-PCR of theagrlocus. Chi-square tests (carried out on the actual number of occurrences) using fourfold tables and the correction of continuity devised by Yates (23) were used to analyze the significance of the differences in the distribution of strains by population origin. Quadruple asterisks,Pvalue of⬍0.001; Triple asterisks,Pvalue of⬍0.01 and⬎0.001; double asterisks,Pvalue of⬍0.10 and⬎0.05; asterisk,Pvalue of⬍0.50 and⬎0.10.
VOL. 42, 2004 NOTES 1267
on May 15, 2020 by guest
http://jcm.asm.org/
[image:3.603.78.507.73.324.2]FIG. 3. Dendrogram showing the clustering of 50 unrelated humanS. aureusisolates and 69 unrelated bovineS. aureusisolates on the basis of hybridization scores after BT with 12 DNA probes. Isolates numbers,agrtypes,agrgroups, binary codes, and origins of strains are shown for all isolates. Roman numerals indicate the three main clusters of strains defined at a genetic similarity of 83% (arrow). The scale indicates the level of genetic relatedness within this collection of strains.
1268
on May 15, 2020 by guest
sistent with the host specificity concept amongS. aureusstrains and showed that the transfer of strains between humans and cows is a possible but infrequent event. We believe that all strains are putatively able to infect one or the other host, but our typing results indicate that most of them are much more adapted for the infection of one host versus the other. At this moment, the underlying genetic basis for the variation among
S. aureusstrains from both populations is unknown, but at least
two hypotheses can be conceived: (i) theagrallele of a strain is directly implicated in the regulation of the expression of genes implicated in the tropism of the strain for a particular host; (ii) theagrallele is not directly involved in host specificity, but it reveals the overall genetic evolution ofS. aureus after its ad-aptation (host-adapted lineage[s]) to one or the other host by the acquisition (or the loss) of some virulence genes and/or pathogenicity islands. The University of Minnesota is sequenc-ing the entire genome of a common bovineS. aureusstrain. A comparative genomic analysis of a shotgun sequence of around 10% of the genome of this bovine mastitis-associated clone and the genomes of the recently sequenced human-associated strains Mu50 and N315 has already identified numerous unique sequences (3, 11). A recent study also identified a novel putative pathogenicity island, SaPIbov, in the genome of this bovine isolate (1). For these reasons it can be expected that the identification of lineages dominant in a particular host or dis-ease, and further comparative genomic analysis will provide important insights regarding disease pathogenesis and lead to the development of new prophylactic and diagnostic tools.
This work was supported by a grant (AIP P00060, P00223) from the French association Bureau des Ressources Ge´ne´tiques.
We thank Martine Braibant for critical reading of the manuscript.
REFERENCES
1. Fitzgerald, J., S. Monday, T. Foster, G. Bohach, P. Hartigan, W. Meaney, and C. Smyth.2001. Characterization of a putative pathogenicity island from bovineStaphylococcus aureusencoding multiple superantigens. J. Bacteriol.
183:63–70.
2. Gilot, P., G. Lina, T. Cochard, and B. Poutrel.2002. Analysis of the genetic variability of genes encoding the RNA III-activating components Agr and TRAP in a population ofStaphylococcus aureusstrains isolated from cows with mastitis. J. Clin. Microbiol.40:4060–4067.
3. Herron, L., R. Chakravarty, C. Dwan, J. Fitzgerald, J. Musser, E. Retzel, and V. Kapur.2002. Genome sequence survey identifies unique sequences and key virulence genes with unusual rates of amino acid substitution in bovine
Staphylococcus aureus.Infect. Immun.70:3978–3981.
4. Jarraud, S., G. Lyon, A. Figueiredo, G. Lina, F. Vandenesch, J. Etienne, T. Muir, and R. Novick.2000. Exfoliatin-producing strains define a fourthagr
specificity group inStaphylococcus aureus.J. Bacteriol.182:6517–6522. 5. Jarraud, S., C. Mougel, J. Thioulouse, G. Lina, H. Meugnier, F. Forey, X.
Nesme, J. Etienne, and F. Vandenesch.2002. Relationships between Staph-ylococcus aureusgenetic background, virulence factors,agrgroups (alleles), and human disease. Infect. Immun.70:631–641.
6. Ji, G., R. Beavis, and R. Novick.1995. Cell density control of staphylococcal virulence mediated by an octapeptide pheromone. Proc. Natl. Acad. Sci. USA92:12055–12059.
7. Ji, G., R. Beavis, and R. P. Novick.1997. Bacterial interference caused by autoinducing peptide variants. Science276:2027–2030.
8. Kapur, V., W. M. Sischo, R. S. Greer, T. S. Whittam, and J. M. Musser.1995. Molecular population genetic analysis ofStaphylococcus aureusrecovered from cows. J. Clin. Microbiol.33:376–380.
9. Kluytmans, J., A. van Belkum, and H. Verbrugh.1997. Nasal carriage of
Staphylococcus aureus: epidemiology, underlying mechanisms, and associ-ated risks. Clin. Microbiol. Rev.10:505–520.
10. Kreiswirth, B., J. Kornblum, R. Arbeit, W. Eisner, J. Maslow, A. McGeer, D. Low, and R. Novick.1993. Evidence for a clonal origin of methicillin resis-tance inStaphylococcus aureus.Science259:227–230.
11. Kuroda, M., T. Ohta, I. Uchiyama, et al.2001. Whole genome sequencing of methicillin-resistantStaphylococcus aureus.Lancet357:1225–1240. 12. Lina, G., F. Boutile, A. Tristan, M. Bes, J. Etienne, and F. Vandenesch.2003.
Bacterial competition for human nasal cavity colonization: role of staphylo-coccalagralleles. Appl. Environ. Microbiol.69:18–23.
13. Lowy, F.2000. IsStaphylococcus aureusan intracellular pathogen? Trends Microbiol.8:341–343.
14. Miles, H., W. Lesser, and P. Sears.1992. The economical implications of bioengineered mastitis control. J. Dairy Sci.75:596–605.
15. Moore, P. C., and J. A. Lindsay.2001. Genetic variation among hospital isolates of methicillin-sensitiveStaphylococcus aureus: evidence for horizon-tal transfer of virulence genes. J. Clin. Microbiol.39:2760–2767.
16. Mullarky, I. K., C. Su, N. Frieze, Y. H. Park, and L. M. Sordillo.2001.
Staphylococcus aureus agrgenotypes with enterotoxin production capabilities can resist neutrophil bactericidal activity. Infect. Immun.69:45–51. 17. Novick, R. P., H. F. Ross, S. J. Projan, J. Kornblum, B. Kreiswirth, and S.
Moghazeh.1993. Synthesis of staphylococcal virulence factors is controlled by a regulatory RNA molecule. EMBO J.12:3967–3975.
18. Novick, R.2000. Pathogenicity factors ofStaphylococcus aureusand their regulation, p. 392–407.InV. Fischetti (ed.), Gram-positive pathogens. ASM Press, Washington, D.C.
19. Papakyriacou, H., D. Vaz, A. Simor, M. Louie, and M. McGavin.2000. Molecular analysis of the accessory gene regulator (agr) locus and balance of virulence factor expression in epidemic methicillin-resistantStaphylococcus aureus.J. Infect. Dis.181:990–1000.
20. Peacock, S., C. Moore, A. Justice, M. Kantzanou, L. Story, K. Mackie, G. O’Neill, and N. Day.2002. Virulent combination of adhesin and toxin genes in natural populations ofStaphylococcus aureus.Infect. Immun.70:4987– 4996.
21. Sakoulas, G., G. Eliopoulos, R. Moellering, Jr., R. Novick, L. Venkataraman, C. Wennersten, P. DeGirolami, M. Schwaber, and H. Gold.2003. Staphylo-coccus aureusaccessory gene regulator (agr) group II: is there a relationship to the development of intermediate-level glycopeptide resistance? J. Infect. Dis.187:929–938.
22. Shopsin, B., B. Mathema, P. Alcabes, B. Said-Salim, G. Lina, A. Matsuka, J. Martinez, and B. Kreiswirth.2003. Prevalence of agr specificity groups amongStaphylococcus aureusstrains colonizing children and their guardians. J. Clin. Microbiol.41:456–459.
23. Swinscow, T. D. V.1978. The chi square tests, p. 42–53.InT. D. V. Swinscow (ed.), Statistics at square one. Dowson and Goodall Ltd., The Mendip Press, Bath, England.
24. Takeuchi, S., T. Maeda, N. Hashimoto, K. Imaizumi, T. Kaidoh, and Y. Hayakawa.2001. Variation of theagrlocus inStaphylococcus aureusisolates from cows with mastitis. Vet. Microbiol.79:267–274.
25. Tenover, F., R. Arbeit, G. Archer, J. Biddle, S. Byrne, R. Goering, G. Han-cock, G. He´bert, B. Hill, R. Hollis, W. Jarvis, B. Kreiswirth, W. Eisner, J. Maslow, L. McDougal, J. Miller, M. Mulligan, and M. Pfaller.1994. Com-parison of traditional and molecular methods of typing isolates of Staphylo-coccus aureus.J. Clin. Microbiol.32:407–415.
26. van Belkum, A., N. Riewarts Friksen, M. Sijmons, W. van Leeuwen, M. van den Bergh, J. Kluytmans, F. Espersen, and H. Verbrugh.1997. Coagulase and protein A polymorphisms do not contribute to persistence of nasal colonisation byStaphylococcus aureus.J. Med. Microbiol.46:222–232. 27. van Leeuwen, W., A. van Belkum, B. Kreiswirth, and H. Verbrugh.1998.
Genetic diversification of methicillin-resistantStaphylococcus aureusas a function of prolonged geographic dissemination and as measured by binary typing and other genotyping methods. Res. Microbiol.149:497–507. 28. van Leeuwen, W., H. Verbrugh, J. van der Velden, N. van Leeuwen, M. Heck,
and A. van Belkum.1999. Validation of binary typing forStaphylococcus aureusstrains. J. Clin. Microbiol.37:664–674.
29. van Leeuwen, W., W. van Nieuwenhuizen, C. Gijzen, H. Verbrugh, and A. van Belkum. 2000. Population studies of methicillin-resistant and -sensitive
Staphylococcus aureusstrains reveal a lack of variability in theagrD gene, encoding a staphylococcal autoinducer peptide. J. Bacteriol.182:5721–5729. 30. van Leeuwen, W., C. Libregts, M. Schalk, J. Veuskens, H. Verbrugh, and A. van Belkum.2001. Binary typing ofStaphylococcus aureusstrains through reversed hybridization using digoxigenin-universal linkage system-labeled bacterial genomic DNA. J. Clin. Microbiol.39:328–331.
31. Zadoks, R., W. van Leeuwen, H. Barkema, O. Sampimon, H. Verbrugh, Y. H. Schukken, and A. van Belkum.2000. Application of pulsed-field gel electro-phoresis and binary typing as tools in veterinary clinical microbiology and molecular epidemiologic analysis of bovine and humanStaphylococcus au-reusisolates. J. Clin. Microbiol.38:1931–1939.
VOL. 42, 2004 NOTES 1269