Green Rust: The Simple Organizing ‘Seed’ of all Life?
Michael J Russell
Planetary Chemistry and Astrobiology; Jet Propulsion Laboratory; California Institute of Technology, Pasadena, CA 91109-8099, USA;
E-Mail: [email protected]
The most important problem of synthetic biology is…the reduction of carbonic acid.
and
Without the idea of spontaneous generation and a physical theory of life, the doctrine of evolution is a mutilated hypothesis without unity or cohesion.[1]
Leduc, [1]
Abstract: Korenaga and coworkers present evidence to suggest that 4.3 billion years ago the
Earth’s mantle was dry and water filled the ocean to twice its present volume.[2] CO2 was
constantly exhaled during the mafic to ultramafic volcanic activity associated with magmatic plumes that produced the thick, dense and relatively stable oceanic crust. In that setting two distinct major types of sub-marine hydrothermal vents were active: ~400°C acidic springs whose effluents bore vast quantities of iron into the ocean, and ~120°C, highly alkaline and reduced vents exhaling from the cooler, serpentinizing crust at some distance from the heads of the plumes. When encountering the alkaline effluents, the iron from the plume head vents
precipitated out forming mounds likely surrounded by voluminous exhalative deposits similar to the banded iron formations known from the Archean. These mounds and the surrounding
sediments likely comprising nanocrysts of the variable valence FeII/FeIII oxyhydroxide, green rust. The precipitation of green rust, along with subsidiary iron sulfides and minor concentrations of Ni, Co and Mo in the environment at the alkaline springs may have established both the key bio-syntonic disequilibria, and the means to properly make use of them – those needed to drive the essential inanimate-to-animate transitions that launched life. In the submarine alkaline vent model for the emergence of life specifically it is first suggested that the redox-flexible green rust microcrysts spontaneously formed precipitated barriers to the complete mixing of carbonic ocean and alkaline hydrothermal fluids, barriers that created and maintained steep ionic disequilibria; and second, that the hydrous interlayers of green rust acted as 'engines' that were powered by those ionic disequilibria and drove essential endergonic reactions. There, aided by sulfides and trace elements acting as catalytic promoters and electron transfer agents, nitrate could be reduced to ammonia and carbon dioxide to formate, while methane may have been oxidized to methyl and formyl groups. Acetate and higher carboxylic acids could then have been produced from these C1 molecules and aminated to amino acids, and thence oligomerized to offer peptide nests to phosphate and iron sulfides and secreted to form primitive amyloid-bounded structures, leading conceivably to protocells.
Keywords: Hadean, carbonic ocean, mantle plumes, Banded Iron Formation, green rust,
1. Introduction
Chemistry unbridled, could not have led to life.[1,3,4]. Redox and pH gradients were required, together with 'engines of disequilibria conversion’ to allow these gradients to do the work of surmounting the steep endergonic barriers encountered by C1 molecules that they might react and complexify along the pathways and, eventually, around the autocatalytic cycles of life, just as the autotrophs feeding the base of the food web do today. But what materials and what environments could have allowed these gradients to be used to drive the key endergonic processes needed to launch life? In our original alkaline hydrothermal vent (AHV) model we suggested that life emerged at an alkaline hydrothermal spring on meeting the carbonic Hadean Ocean.[5]). At the macro level the spontaneously precipitated sulfides and hydroxides could induce just the redox and pH gradients, and with the same polarities, as extant life. And the precipitate mounds could provide the conversion engines specific to the transformations that needed to be made in the ambient environment, sufficient to drive life’s emergence.[3,4,6] But it became clear that mere porous cavities could not provide the low water activities and the pumps beyond the merely osmotic, to take on the requirements of an autotrophic metabolism, let alone its need for adaption, guidance and waste disposal. Moreover, a mineral seed has struck many as lacking the versatility of organic molecules whatever their provenance. So, when Arrhenius [7] also called upon a crystal as being a host mineral of biopoesis, it was not obvious then that a genuine epiphany had been realized.[7] Arrhenius’ crystal group of choice were the
Doppelschichtstrukturen mit brucitähnlichen first decribed by Allmann.[8,9] These double layer hydroxides (strictly the DLH though generally and mistakenly referred to as the layered double hydroxides LDH) may be derived out of the brucite (Mg[OH]2) structure, especially significant
in that brucite itself is often the first mineral to be precipitated at submarine alkaline
hydrothermal vents today.[10,11,12] In the DLH proper, two thirds of the cations are divalent (e.g., Mg2+, Fe2+, Ni2+, Zn2+, Ca2+, Co2+, Mn2+) while every third cation carries a +3 charge (e.g.,
Al3+, Fe3+, Cr3+, Mn3+, Ni3+, Co3+). One such DLH, green rust (~FeII
4FeIII2[OH]12CO3·3H2O),
struck Arrhenius [7] as a particularly good candidate to consider in terms of life’s emergence. It is pliant, contains hydrous interlayers that readily persorb anions and is both responsive and resilient to strong variations in pH and redox. We might think of these interlayers as the viscous ‘innards’, or even as proto-cytoplasm within the spontaneously precipitated and rechargeable green rust. Green rust, along with the subordinate iron sulfide mackinawite ([Fe>Ni]S), functioned, it is proposed, as electrochemical nano-engines acting to convert the imposed external proton and redox disequilibria into the internal disequilibria necessary to bring life into being.[6] These engines might effect ‘free energy’ conversions of various types driven by
Brownian motion with built-in escapement mechanisms that prevent back reactions.[3,4,13 ] It is in this light that it should be possible to discern such recognizable mineral precursors in the geologic record to explain the autogenic emergence of metabolic pathways and cycles supplied from the simplest of carbon substrates, carbon dioxide, formate and methane. [14] In the AHV model, the complex molecular engines mediating present-day electron transfer and conversions are a result of evolution from such abiotic mineral ancestors.[3,4,13]
Thus, the nub of this contribution is to suggest how green rust (~[Fe2+
6xFe3+6(1−x)O12H2(7−3x)]2+.[CO2−3.3H2O]2−), nickel-rich mackinawite ([Fe>Ni]S), greigite
(Fe5NiS8), violarite (Fe2Ni4S8) and possibly tochilinite (FeS[Mg,Fe2+][OH]2), precipitated as the
the emergence of life.[15,16] That green rust is capable of conformational changes during oxidative-reductive and protonation-deprotonation oscillations lends itself to such
investigations.[7,17,18,19] We also speculate on the crucial steps taken toward what must have been the break-out metabolism and an organic takeover of the mineral precursor disequilibrium or ‘free energy’ converters, as they acquire organic frameworks in a process that we have called ‘minerobiolization’. We then review the status of efforts to test these and related ideas bearing on the issue and conclude with a rather long list of falsifiable predictions of the submarine AHV as it now stands! The transition from inorganic- toward partly organic-enabled molecular
mechanisms was, we believe, just the very first of many subsequent evolutionary steps made through the adaption of a chance function derived in one context for use in another. In the text, references to analogous modern mechanisms to those considered to have first begat life can be thought of as the direct legacies from, and as being broadly homologous with, their mineral progenitors. With these assumptions in mind we will suggest a testable model whereby the first and simplest pathways and enzymatic mechanisms are assumed to reflect on early beginnings and availabilities of disequilibria, mechanisms for their conversion, as well as on the trace metals and phosphate that contributed to these processes then and now.
2. Model assumptions
The submarine AHV model for life’s emergence focusses on the interaction between C1 molecules, and proton and electron gradients, acting across membranes for autogenesis.[20,21] The iron oxyhydroxides and sulfides comprising the membranes act as disequilibrium
converters.[13,16] However, these and several other key aspects of the AHV theory need
experimental testing, particularly at pressures high enough (~10 bars) to keep hydrogen, methane and CO2 in sterile solution. Nick Lane’s group have had some success testing this autotrophic
(autogenic) hydrothermal model for the emergence of life by attempting to reproduce molecules of the reductive acetyl coenzyme-A pathway (rAcCoA).[22]. This group tested an abiotic CO2
-fixing pathway employing a rig that models the submarine alkaline vent whereby CO2 dissolved
in an acidulous ocean analog on one side of an FeS-bearing membrane is juxtaposed to an alkaline hydrothermal solution on the other.[23,24,25,26,27,28] An endergonic reaction was expected to be driven by the protons pent up in the ocean (the original and natural proton motive force) as they permeated the iron sulfide chimney toward the alkaline interior along a single pathway.[22,29,30] Yields of CO2 reduction products in these experiments were ~50 µM formate
along with a variable and inconsistent formaldehyde concentration of around 100 nM. These authors speculate that the formaldehyde product should be enough to initiate the formose reaction and produce thereby, various sugars. However, formaldehyde is a significant
contaminant in buildings and laboratories, which might explain the non-reproducibility of their results and give pause for thought.[31,32]
Ryuhhei Nakamura and his group have also provided partial experimental support for the AHV theory.[33] Using iron-nickel sulfides they demonstrate the electro-catalytic reduction of CO2, whereby the 3-4 pH units operating as the putative proton motive force (PMF) is
“converted” to the ~200 mV over-potential required in the natural reduction.[33] Yamaguchi and collaborators caution that the PMF is rapidly expended through reduction of H+ unless amines
are to hand to substantially improve the Faradaic efficiency for CO2 reduction.[33,41] Rapid
species, and at the same time increase the durability of the membrane.[20,34,35,36,37,38] Much of any H2 generated in these conditions would be absorbed in the sulfides which could act as H2
stores as well as – through the re-reduction of the greigite to mackinawite – induce a homeostatic pH regulating mechanism.[33,39,40] The two products of these electrochemical reductions are CO and CH4. Beyond the carbon monoxide, no intermediates such as formaldehyde or a methyl
group, from which a metabolic pathway might be fed, are recorded.
More in line with the AHVT, an early experiment by Huber and Wächtershäuser did demonstrate the production of thioacetate directly from carbon monoxide using iron and/or nickel as catalysts in neutral and alkaline conditions.[42] Their experiment called for the activated-methyl group, methane thiol, generated from CO2 and H2S or FeS/HCl and H2S – an
ostensibly autotrophic reaction.[44] However, yields of the methane thiol were limited to 0.1% of the H2S feed, the initial concentration of which was ~1mM/L at most, suggestive of a ≤1
µM/L thiol concentration — less than one ten thousandth of the concentration of CO2 around the
vent.[43,45,46,47]. That this experiment requires repeating is made the clearer by the fact that thiols are, against expectation, seemingly absent from submarine hot springs.[48] Nevertheless, these experiments do partially address Luduc’s assertion that “The most important problem of synthetic biology is not so much the synthesis of the albuminoids as the reduction of carbonic acid” –anargument echoed by the mineralogist Goldschmidt, and given biochemical foundation by Fuchs.[1,24,49,50]
Thus, in the face of these results and speculations, it might seem perverse to suggest instead a two-route autogenic pathway to the metastable intermediates of life. However, as the submarine alkaline vent environment offers millimolar-levels of both the fully-reduced carbon end member CH4, as well as the stable fully-oxidized carbon end member CO2, we have
formulated a hypothesis involving these feeds from either end of the full C1 redox span to produce activated acetate, the target molecule of the first step to metabolism.
[23,24,51,52,53,54,55] This two-path model to activated acetate – an alternative possibility yet to be experimentally tested – has been termed ‘denitrifying methanotrophic acetogenesis’.[54] In this model CO2 is reduced to formate or CO with electrons either initially provided by oxidation
of FeII as in the experiments mentioned above, or by a reversal of the formate hydrogen lyase reaction.[15] At the same time the hydrothermal CH4 is converted to a methyl group in complex
reactions involving molybdenum in a 2-electron redox cluster that have its electrons drawn, one to the high electron acceptor nitrite generated in green rust, converting it to nitric oxide, while the other, now more strongly reducing electron, is accepted by a greigite (Fe5NiS8) cluster (cf.
CO-dehydrogenase).[54] The oxygen atom of the nitric oxide is activated by the high potential electron from the molybdenum center which now has the power to oxidize the methane to a methyl group.[54] The two lost electrons are replaced by those released on the oxidation of hydrothermal hydrogen at a NiFe (Ni-bearing greigite) center acting as a hydrogenase. The resulting methyl group is further oxidized to a formyl group and then sulfidized to methane thiol, the two uniting to form the activated acetate as in the Huber-Wächtershäuser
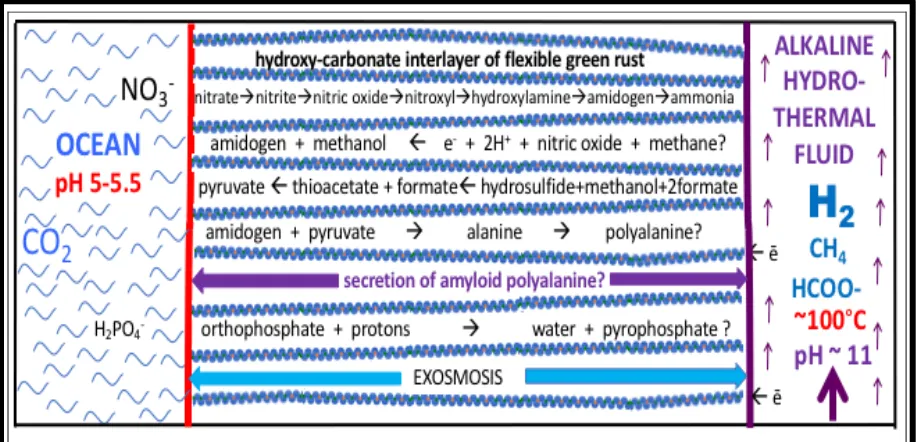
experiment.[15,43,54] The electron acceptors in this modified AVT are nitrate and nitrite (the first ‘breathing’ entities of life) – derived from the nitrogen oxides produced mainly through cloud-to-cloud lightning and rained into the Hadean ocean.[54,56,57,58] Eventually reaching micromolar concentrations this oxidant, along with ≤350 mM of carbon dioxide are carbureted to the mound’s margins through secondary convection and entrainment (Figure 1, Table 1).
Table 1. Steps toward ligand-accelerated autocatalytic denitrifying methanotrophic acetogenesis: demonstrated, probable, possible and predicted, with comparisons to enzymes.
Biosyntonically ‘engineered’ steps
cf. prebiotic enzyme analogues
Mineral barriers, engines, catalysts
Abiotic reaction coupling and/or
gradient
References
0. {5OH- + HS-} + 2Fe2+ + Fe3+ + Ni2+
à{FeS + Fe2(OH)5} + ē membrane
Green Rust & [FeNi]S set in SiO2?
spontaneous barrier
precipitation 40,45,299
1. H2à 2H• à 2H+ + 2e- NiFe[Mo]-H2ase GR>FeS>NiS>MoSchimneys 2/ redox gradient 126,129
1a. proton-coupled electron transfer processes fd GR, mackinawite, greigite proton gradient 103
1b. electron bifurcation, conformation plasticity,
electron and proton transfer, gating MMO,Nir GR, mackinawite, greigite, MoSx redox gradient 6,18,28,123,
2. H+ + 2ē + CO
2à HCOO- + H2O FHL Ni3Fe, or [FeNi]S or MoS redox, pH gradient, serpentinization, or 22,141,285, 300,301
2a. CO2 + 2ē + H+à CO + OH- CODH violarite electron conduction 33,297
3. CH4 + NO3- + H2 + H+ à •CH3 + 2H2O + NO MMO
GR & Mo-dosed greigite (redox/
pH gradients)
redox and pH gradient undemonstrated
103,219,287, 293,294
4. NO3- + 4H2 + 2H+à NH4+ + 3H2O
Nar/Nir/NOR GR (redox/ pH gradients) redox (~180 mins) 161,168;
5. •CH3 + OH-/SH-?) à CH3OH/ CH3SH) + ē
MMO? GR? high T undemonstrated None
6. CH3OH + [2FeIII] à HCHO + [2FeII] + 2H+
MDH GR [FeNi]S? Fe2(MoO4)3 undemonstrated 192
7. HCHO+HP2O73- + [OH-] à [HCOPO4]2- +HPO42-
FK ? undemonstrated exergonic but 133
8. HCOPO42- + HS- + 2H+ + 2ē à CH3S- + HPO42- ? undemonstrated exergonic but 133,296
9. CH3S- + HCOO- + H+à CH3COOH + HS-
ACS Fe4NiS9(HN)2 cf. Reppe chemistry, 29,43,288
9a. CH3S- + CO à CH3COS- Fe4NiS9(HN)2 High yield (20 hours) 42
9b. (CH3COS- + HPO42-à CH3COPO42- + HS-) ? low yield 302
10. HCOO- + CH
3CO~SCH3 + ē à CH3COCOO- +
HSCH3- PFL Fe2(RS)2(CO)6 undemonstrated 217,289
10a. CH3COCOO- + (HP2O7)3- + CO2à
10b. CH2=C(OPO3)2-COO- + CO2 + H2O à -OOCCH
2COCOO- + HPO42- + H+ ACC GR/mackinawite? predicted 16
11. CH3COCOO- + NH4+ + 2ē + 2H+à
CH3CH(NH2)COO- + H2O ALT GR/mackinawite? 24 hours 38
12. (CH3CH(NH2)COOH)4 + CH3CH(NH2)COOHà
CH3CH(NH2)CO-CH3CH(NH)CO-CH3
CH(NH)CO-CH3CH(NH)CO-CH3CH(NH)COOH + 4H2O DLH [cf. “DNA pol”!]
dolomite (ab initio simulations
mackinawite and double layer hydroxide)
spontaneous (dolomite)
199,290,291, 292,298
13. Pi + Pi à PPi by GR H+-PPase FeS, GR only at equilibrium 16,181
ligand-assisted recapitulation?
{13} poly-alanine peptide-strengthened
membrane? mineral-organic framework spontaneous 45,125
{14} SGAGKT peptide + Pi à P-loop 6mer peptide spontaneous 239
{15} CH3CH(NH2)CO-CH3
CH(NH)CO-CH3CH(NH2)CO-CH3CH(NH)COOH+ Ni2+ à Ni-CH3CH(NH2)CO-CH3
CH(NH)CO-CH3CH(NH)CO-CH3CH(NH)COOH ATCUN motif
4mer peptide spontaneous 235,236,238,295
{16} (Fe4NiS) + CH3CH(NH2)CO-CH3
CH(NH)CO-CH3CH(NH)CO-CH3
CH(NH)CO-CH3CH(NH)COOH
à [Fe4NiS]-CH3CH(NH2)CO-CH3
CH(NH)CO-CH3CH(NH)CO-CH3
CH(NH)CO-CH3CH(NH)COOH
Proto-fd, ACS, CODH
6mer peptide partial demonstration 189
à{16}{1}{2}{3}{4}{5}{6}{7}{8}{9}{10}{11}{12}{13}à
repeat GR breakout metabolism? Figure 1
ACC = Acetyl-CoA carboxylase; ACS = Acetyl-CoA synthase; ALT = Alanine
transaminase; ATCUN motif= Amino terminal Cu(II) and Ni(II) bindingmotif; CODH = Carbon monoxide dehydrogenase; (DLH = Double Layer Hydroxide; DNA pol = DNA polymerase); Fd = Ferredoxin; FHL = Formate hydrogen lyase; FK = Formate kinase; H+
-PPase = proton pyrophosphatase; MMO = methane monooxygenase; MDH = methanol
dehydrogenase; Nar = nitrate reductase; Nir = nitrite reductase; NOR = nitric oxide reductase;
NiFe[Mo]-H2ase = NiFe[Mo]-hydrogenase; PFL = Pyruvate formate lyase.
3. The Hadean water world
Prior to the advent of plate tectonics and following a rapid rain-out at ~4.4 Ga, about two present-day ocean volumes likely enveloped an entirely submerged mafic Hadean crust
comprising extensive igneous provinces fed by large mantle plumes.
[58,59,60,61,62,63,6465,66,67] Judging from a modern example, the Ontong Java Plateau (that portion distant from the collision zone with the Solomon Island Arc), Hadean crust would have lain several kilometers below sea level, well out of the way of the so-called Late Heavy
strictly chondritic (i.e., “primitive”) magma reservoir through the ‘heat-pipe’ tectonics of mantle plumes.[72] And these zircons have none of the indicators for fractionation that might otherwise suggest the development of an emergent Hadean continental crust.[73,74] There really was no fresh water to be had.
Furthermore, conditions at the Hadean ocean surface were certainly less than clement! The length of the day was shorter, the UV flux destructive to molecules of any length or
intricacy, the moon close, the tides twenty times stronger, and the weather an unchecked roiling maelstrom (Figure 1).[76,77] Thus, there were no candidate land-based ‘birthing’ pools to periodically dry out and ‘drive’ polymerization of ‘in-falling organic compounds’ to produce at best, a picomolar organic soup, which anyway would be of doubtful nutritious value. Such conditions lay nearly a billion years into the future, well after life’s onset, and are in any case irrelevant to the emergence of life.[6,78,79]
Extrapolating backwards from early Archean exposures in Greenland, which may evidence the onset of plate tectonics, the Hadean ocean crust of this water-world was likely covered in green rust (~[Fe2+6xFe3+6(1−x)O12H2(7−3x)]2+.[CO2−3.3H2O]2−) and silica gels derived
from a myriad of sulfate-poor acidic ~400°C hydrothermal springs.[80,81,82] These very hot springs were driven by heat from the mantle plumes and bore many tens of mM iron Fe into the Hadean carbonic ocean.[82] The pH of this ocean, as back-extrapolated and assuming
equilibrium with CO2 and metal solubilities, has generally been over-estimated, a consequence of
failing to factor in the 50-fold supersaturation of iron in carbonic fluid.[83] However, empirical evidence from iron-rich carbonic lakes in Cameroon suggest instead, a pH of 5 to 5.5, although such a low pH is not critical to the AHV theory.[83,84, 85] Green rust would only have
precipitated from this acidulous ocean on meeting the alkaline hydrothermal
fluids.[7,76,81,82,85,86,87,88,89,90] On burial, this green rust converted to the hematite and magnetite constituting the thinly laminated banded iron formations (BIFs) comparable to those we see today.[89,90] With a specific gravity of around 5 and interlayered with basalt/komatiite and chert with specific gravities of ~2.9 and 2.5 respectively, the Hadean crust was ‘doomed’ to founder back into the hot, dry mantle, so explaining its absence.[80]
4. The precipitate mound at the submarine alkaline vent
As noted, green rust would have been precipitated from iron derived ultimately from high temperature acidic springs. This iron had remained supersaturated until interfacing alkaline fluids.[83,89] A major source of such alkaline waters would have been “Lost City-type” moderate temperature alkaline springs.(5,12, 89,90,91,92,93] Subsidiary hydrothermal mackinawite [Fe>Ni]S, greigite ~Fe5NiS8), amorphous silica (SiO2) and phosphates
(~FeII
3[PO4]2·8H2O) along with a few scattered grains of molybdenite, were likely to have been
co-precipitates (Table 1, Figure 1). [20,28,34,45,85,93,94,95,96,97,98,99,100,101,102,103] At alkaline hydrothermal vents in today’s ocean, brucite (Mg[OH]2) – a single layer
hydroxide – is an early precipitate.[12,92] Such a wet environment is seen by some as a fatal flaw in Submarine Alkaline Hydrothermal Vent (AHV) theory and they instead call upon wet-dry cycling in surface pools and ponds. However, as Philip Ball reminds us, water is not only “life’s solvent”, but acts as a shell to, and interacts with, both proteins and their
substrates.[104,105,106,107,108] Indeed, water composes 70–80% of the cell mass where it is deeply involved in the orchestration of cellular machinery.[109] Furthermore, all of the
mediation of protein machines – both to control the water activity at the active site, but also to provide a means by which these inherently endergonic processes can be thermodynamically driven – merely brings forward what the 'mineral machines' of 'first life' must also be capable of doing (and must be shown to be capable of doing) (Figures 1 and 2).
The rapidly precipitated nano-crystalline green rust and mackinawite comprising the alkaline mounds would also have included ambient water – confined and somewhat immobilized through hydrogen bonding in galleries 0.3 nm deep between the ‘brucite’ layers (e.g., in
[FeII4FeIII2(OH)12][CO3].3H2O). Thus, far from being a negative factor in the AHV theory for
life’s emergence, the intimate polar interfacing with constitutional water in green rust is just one of the many common features that could link such minerals to emergent
life.[7,109,110,111,112,113] And the anions required for charge parity in green rust were engaged through ion-dipole interactions with FeIII/FeII cations.[114,115] In strong contrast, nano-confinement of water in mackinawite greatly enhances its self-dissociation, entailing no entropic cost and permits almost frictionless flow between the tetrahedral sheets.[116] Proton flow comparable to that in bulk water in these circumstances would have been via the Grotthuss mechanism at a rate of around 25 nm ns−1.[117] This is one of the reasons we are now persuaded
that it was the green rust and mackinawite micro- and nano-crysts and their associated interlayer galleries filled with structured or ‘bulk’ water respectively that were life’s cellular precursors, rather than the fluid-filled chambers or gels that we, and others, first imagined to have borne the first cytoplasm.[5,45]
In the living cell, for example, water molecules are hydrogen-bonding to iron in heme and iron hydroxylase, to magnesium in the ribosome, and, for a pertinent organic example, water molecules stabilize and activate C-methyltransferase.[118,119,120,121] So, although the
founding submarine alkaline mound was completely immersed in water, bulk water was
excluded at the site of some reactions in the interlayers of green rust and mackinawite, just as it is by proteins, or in lipid membranes. And these are the minerals that not only provided the spontaneously deposited barrier that kept the two contrasting fluids from immediate mixing, but also, dosed with trace elements Ni, Zn and Co, they constitute the inventory from the minéral trouvé that could have acted as nano-engines and electron transfer agents (i.e., disequilibrium converters and ‘free energy’ transformers) at life’s emergence.[13,122,123] In the following we will examine the chief of these – the layered nanocrysts, green rust and mackinawite. Notably, these can be produced in the lab with 10 mmol FeII iron concentrations, well below the many tens of mmol of iron in the ~ 400ºC spring sources mentioned above (Figures 1 and
2).[101,124,125] We will also touch on the likely, or possible, accompanying minor minerals, greigite, violarite and tochilinite. But we first consider the implication of a foundational
presumption of the AHV theory: that life itself was driven into being by thermodynamic forces - just as has also been, and by absolute necessity, every living thing since. What can we
reasonably conjecture about what those original forces were that could have had that consequences?
5. The disequilibria imposed across the mineral barrier
Given that life, in its bid to hydrogenate carbon dioxide and, perhaps, oxidize methane to a methyl group, takes its power to do so from thermodynamically 'downhill' electron flow, it is reassuring to note demonstrations of electron generation sufficient to light LED bulbs at both actual and experimental hydrothermal vents.[105,126, 127,128 ,129] But whether electrons can also be harvested directly from hydrothermal H2 across an abiotic mineral membrane or more
mackinawite or white rust, or iii) directly through the self-oxidation of nano-crysts of these metastable minerals (Table 1).[45,130]
It is well established that the tensions between volcanic carbon dioxide against the hydrogen exhaling into the acidulous ocean of young wet rocky planets, cannot be released through mere geochemical reactions beyond formate because of thermodynamic
barriers.[29,58,131] Even electrochemical experiments involving steep over-potentials do not produce the chemical intermediates that populate autotrophic metabolic pathways such as methyl thioacetate (CH3COSCH3) which requires assailing the thermodynamic barrier posed by
formaldehyde.[131] However, the endergonic reduction of carbon dioxide through formate to a formyl group is achieved in biology by coupling it to any one of three types of exergonic driving reactions: 1) the relaxation of a proton gradient, 2) the hydrolysis of polyphosphates or 3)
relaxing a redox gradient via electron bifurcation.[19,22,132,133,134,135] All of these disequilibria conversions must be, of course, carried out by macromolecular nano-engines (disequilibrium converters) and, as we have intimated, we have every reason to assume that nano-engines were coopted from the mineralized environment obtaining in the alkaline
hydrothermal mound.[136,137,138] For reasons stated in section 2, we appeal to hydrothermal methane to provide reduced carbon from the other end of the redox scale to the system along with formate and now consider how this model corresponds to what is now known of the Hadean world.
The iron-rich mound generated at the alkaline vent offers an ambient trans-membrane proton motive disequilibrium or force (DpH 4 to 5) as emphasized by Herschy and his
colleagues.[22] It also provides a redox gradient of 500mV or so comprising hydrogen plus methane versus nitrate and FeIII, MnIV plus carbon dioxide. At the same time hydrothermal
hydrogen at ~15 mmol/kg and formate at ~150 µmol/kg are born towards, and focused at, the mound.[139,140,141,142] Hydrogen is the sine qua non electron donor used by all life still in the form of NADH.[143,144] Indeed, in this way billions upon billions of electrons are transported to the vent in hydrothermal solutions exhaling at ~1000kg/sec in the present day.[145,146] Temperatures too were appropriate (30° to 100°C), as were pressures (≥100 bars), and longevity (~1020 nanoseconds).[93] The trace metals required for catalysis would be precipitated with the
sulfides and oxides on meeting the margins of the alkaline vent fluid.[125,147] However, as we have argued, endergonic reactions (a positive DG) can't be made to happen by mere catalysis, which can only reduce an activation energy, so mere catalysis cannot explain the emergence of life.[3,6] We need instead, nanoscale disequilibrium converters situated at the margins of the alkaline hydrothermal mound to take advantage of the various gradients imposed by the
precipitation of relatively insoluble inorganic entities. This seems a tall order: that one or more of such mineral microcrystals might also act as the thermodynamic conversion engines driving emergent metabolism in the sense of Cottrell seems counter intuitive, viz., "The most promising path to solving this riddle appears at present to be through investigating whether” their local redox-state effects on gallery conformation, especially in green rust, could drive the needed proto-biosynthetic reactions while retaining their overall integrity.[148]
We hypothesize that the proton and electron gradients, as in life, have the theoretical potential to drive endergonic reactions toward a break-out metabolism.[16] For one thing, the disequilibrium between the redox couples H+/H2 and NO3-/NO2- of ~ 800 millivolts is also now
well established.[58,150] H+/H2 and the NO3-/NO2-. Involved in electron bifurcation, the
likely oscillated around 5 pH units given the continual escape of CO2 from the so-called
“popping rocks” comprising the ubiquitous mafic lava flows, and the empirical values reported from Lakes Nyos and Monoun in the Cameroon, as against the alkaline (pH ~ 11) hydrothermal solution, producing a gradient of 250 to 300 millivolts (1 pH unit being equivalent to 59
mV).[68,84,149] Thus the exact pH disequilibrium is not critical beyond offering a potential of ~200 mV.
It should be noted here that ferrous carbonate remains super-saturated in these conditions.[83] The fact that both the oxyhydroxides and the sulfides can house substantial nickel concentrations in structures affine with active sites in the metalloproteins makes them promising promotors, disequilibrium converters (‘free energy transducers’) or nanomotors and electron transfer agents (protoenzymes) at life’s emergence. However, the big question remains as to how these disequilibria were coupled to surmount those endergonic reactions along the pathways to metabolism.
Of course, a detailed research effort is required to reveal how protonic and electronic disequilibria could have been converted abiotically, presumably by the minerals discussed above, to do their work (Figure 2). One obstacle to understanding how exergonic processes are able to drive endergonic ones is quite counter intuitive.[13] For example, such driving does not involve the transfer of energy (even the 'free part' of a literal energy) from the driving process to the driven one. But rather. the engine managing the conversion makes individual fluctuation events dissipating the driving reaction contingent on the coincident occurrence of an endergonic
("reverse") fluctuation in another thermodynamically weaker process, thereby creating a disequilibrium in that process.[13,151,152].
6. Green rust – the first organizing nanoengine of life?
Carbonate green rust (GRcarb) nucleates rapidly to give a plethora of equi-dimensional
planar micro- to nano-crysts generally measuring from 200 nm to ~2 µm across with a a relatively constant aspect ratio of ~1:10.[153,154] So, a 1 µm GRcarb would be about 100 nm
thick and comprise ~130 double layers overall.[154,155,156,157,158] The height of the hydroxyl interlayer galleries is ~0.30 nm, allowing them perhaps to act as anion and proton channels. And the Fe2+-Fe3+ distance of ~0.33 nm along the iron layers would allow electron hopping by
quantum tunneling, though a small proportion of electrons could also jump across the
layers.[18,19] The brucite layers trap a monomolecular interlayer of viscous solution producing solutions concentrated to around 14 molar![7]
Green rust is redox active and can rapidly reduce nitrate to the ammonium ion as well as the redox sensitive ions such as MoVI to MoIV.[159,160,161,162,163,164,165,166,167,168,169]
Such redox-sensitive trace metal ions tend to be reduced and precipitated from their soluble source at, and as, the outer edges of green rust are oxidized.[166] Moreover, such metastability would threaten the integrity of GRs were it not for their partially oxidized margins being highly attractive to orthophosphate (HPO42-). That the ionic diameter of the phosphate ion at 0.48 nm so
Phosphorylation of organic products of redox reactions involving carbon might then ensue.[16] The resulting gaps in the phosphate surrounds to the green rust will be instantly healed as more phosphate is attracted to the positively charged exterior.
To make green rust work as a thermodynamic engine to drive reductions, it would itself have to be continuously re-reduced. In this regard it has been demonstrated that green rust can also be reduced by hydrocarbons without degrading its structure, but whether methane could also reduce relatively oxidized green rust (via activation of NO) as we have suggested, must await experimental test.[54,173,174,175] Another possible, though undemonstrated, method would be the oxidation of hydrothermal hydrogen with nickel-dosed mackinawite or greigite, as in a Ni-Fe hydrogenase.[14,34,176,177] The resulting protons would be immediately neutralized by
reaction with the hydroxyls in the alkaline hydrothermal flux. In other words, it is reasonable to assume that the redox states could be reversed (oscillate) depending on which way the
disequilibria were stacked. Green rust has a low redox potential, being almost as reducing as native iron (Fe0) in anoxic alkaline conditions.[178] Indeed, as we have noted, GRs can compete
with biotic pathways in the reduction of, for example, selenate and nitrate.[160,179] Moreover, we have seen how green rust, with its well-ordered hydrous innards, can act as an aminase. For example, Barge and colleagues have also demonstrated the amination of pyruvate to alanine.[38]
There is an interdependence between the redox potential and the pH of green rusts: oxidation is promoted by deprotonation in alkaline conditions, while the reverse is true of acid conditions where protonation supports reduction.[17] These factors prompted Génin and his collaborators to remark, “Among all compounds containing FeII ions that can play the reservoir of reducing species, the FeII−III oxyhydroxysalts commonly denominated green rusts … constitute an unmatchable compound due to their redox flexibility.[180] As might be expected, solid state reductions of oxidized green rusts can also be driven electrochemically, resulting again in a highly reduced but still morphologically recognizable double layer hydroxide ([FeII
6(OH)10][CO3.2H2O]) (Figure 2).[88,180] Nevertheless, without the protection of organic
intercalates there is a tendency for the Fe2+ to dissolve, thereby threatening the integrity of the
mineral.[20,125,182]
Although these green rust-comprising barriers would have been leaky, individual
crystallites would have frustrated immediate dissipation.[28] Green rusts comprising this barrier would be subject to large redox and pH gradients with ranges that had the potential
commensurate with that required for the onset of metabolism. Electrons could have been
supplied from hydrothermal formate, hydrogen, or from oxyhydroxides and sulfides, dosed with other transition metals comprising the walls of hydrothermal chimneys, as they
oxidize.[126,127,128,129,142] Redox oscillations may have oxidized and re-reduced iron ions between Fe2+ and Fe3+ much as our hemoglobin is cycled between 0 and 4 oxygens many times
over as the red cells are pumped from lung to periphery and back with a cycling time of around 1 minute and with an average life span of 1,000 to 2,000 hours. At the same time the pH gradient would have contributed to protonation and deprotonation of green rust, tending to hold the mineral between the bounds of 2Fe2+/Fe3+ and Fe2+/2Fe3+[167,183,184] The mineral so poised
would have remained metastable, i.e., could have been party to an emergent homeostasis.[185] Such metastability may have been strongly supported by orthophosphate (HPO42-) which, having
compartmental systems with flexible membranes and what may be called primitive cellular metabolic function. Like cells, they retain phosphate-charged reactants against high
concentration gradients and exchange matter with the surroundings by controlled diffusion through the ‘pores’ provided by the opening of the interlayers at the crystal edges. Here, the exposed negative charge on the interrupted metal hydroxide ‘membrane’ leads to sorption of cations as ‘gatekeepers’”[7] He even goes on to speculate that the origin of information might lie in “the variable pattern of cations, conserved within individual (DLH) crystals,” and further, that “such an animation process … would be guided by the subtle electronic forces that metal cations exert on their environment,” while opining “the progress so far in prebiotic chemistry merely suggests that it may be possible to synthesize the tape on which the information for life could be recorded, if we only knew how.”[7]
The margins of green rust would be continually ‘rammed’ by hydronium ions from the ~5 pH ocean as the protons themselves were drawn down-gradient to the pH 11 interior.[4] And their path would be vectored by the natural orientation of these layered minerals normal to the barrier’s surfaces with the hydrous interlayers acting as channels.[117] And it may have been within the confines of green rust at high pH that condensation reactions could have been driven. Erastova and her collaborators – noting how mineral surfaces “may have very high enthalpies of rehydration” and thereby provide “a driving force for condensation reactions” – have suggested, through molecular dynamics simulations, how layered hydroxides can concentrate, align and act as adsorption templates for amino acids, and promote peptide bond formation.[186] Moreover, following Amend and Shock, Kitadai has found that – in regard to the thermodynamics of amino acid synthesis and polymerization – “hot, alkaline hydrothermal systems beneath cool, slightly acidic Hadean ocean are an energetically excellent setting among possible vent-ocean
combinations”.[187,188]
It might be argued that the water generated in these protobiochemical processes would tend to lyse any product. However, we argue that because the founding submarine alkaline mound was completely immersed in salty water both inside and out, bulk-water itself was minimized at the site of these and other reactions as a result of exosmosis from the hydrous interlayers (Figure 2).
7. Iron sulfides: The supporting cast
7.1. Mackinawite [Fe(Ni,Mo)S]: Electron transfer agent, amino acid polymerase and possible hydrogenase
Mackinawite has the extremely low redox potential of -1,010 mV at pH 12 at 25°C , lower even than green rust[189]. And 20% of the total iron in mackinawite can be oxidized before its layered conformation is lost and it converts to the water-absent inverse spinel, greigite (Fe3S4).[101,190] However, both nickel concentrations and the infusion of molecular H2 can
inhibit this transition [101,191,192,193] Indeed, up to ~ 50% of the iron can be replaced by nickel without changing the mackinawite conformation although the solid solution is non-ideal.[193] And Mo:Fe molar ratios in mackinawite precipitated in the presence of
thiomolybdate, can reach ~ 0.05.[194] Both nickel and molybdenum are likely to promote catalytic activity.[103,195]
viscosity therein is similar to that of bulk water – strongly favoring the rapid diffusion of excess protons by the Grotthuss mechanism (116,117,172 ,196,197,198,199,200,201] Mackinawite is also known to be an excellent conductor, suggesting that electron-hopping along the iron layers in mackinawite could be coupled to proton transfer using intermolecular hydrogen-bonds through fluctuation and sequential H-bonding, as in peptides.[19,203] Such coupling of proton flux to electron flow, and vice-versa, would be comparable to the proton-coupled electron transfer (PCET) reduction of nitrite to NO reaction involving cytochrome cd1 in nitrite
reductase.[202,203,204,205,206] Indeed, we can imagine the aqueous interlayers within
mackinawite acting as a proton wire with a rate of proton transfer over 25 nm ns−1 (about a tenth
of the speed of sound) driven by a redox disequilibrium of ~1 eV .[117, 200,207] Yet the hydroxyl ions (or proton holes) tend to be immobilized by hydrogen-bonding to sulfur and the dangling oxygen is partially hydrated, thus inhibiting the diffusion of hydroxyls through the galleries from the alkaline hydrothermal fluid, and thereby favoring proton transfer.[116].
Another biotic-like feature of these nano-confined conditions between the tetrahedral FeS sheets of mackinawite microcrysts is suggested by the ‘nano-solvation’ of glycine and its
polymerization in the presence of carbonyl sulfide (Table 1).[116,199,208,209,210]
7.2. Greigite [Fe3S4]
In the experimental growth of iron sulfide chimneys White and her colleagues found that mackinawite was partially oxidized to greigite [Fe2+Fe3+2S4] between 70° and 75°C.[101] We
assume, after Rickard and Luther, that the reaction is autocatalytic with water as the
oxidant.[213] The hydrogen produced in this reaction maybe occluded or trapped perhaps in the cuboidal entity of the greigite structure where it may be split, with electrons delocalized such that the average charge on the metals is decreased in the cuboid while the sulfides are protonated.[14] The sulfur atoms would remain in place during the transition, though the mackinawite a axis is at a 45º angle to that of greigite.[211] Microcrysts of greigite are magnetic and have low
electrochemical resistance.[220,221,222] Greigite is an anhydrous inverse spinel wherein the apical iron (or nickel) is in the 2+ state, while the delocalized electrons produce an average
charge of 2.5+ on the 4 iron ions in the cuboid.[212] Although the structure of greigite is afine
with the active centers of ferredoxins (e.g., in molybdenum-bearing formate hydrogenlyase and radical SAM, and is comparable to the cuboids in the active centers of carbon monoxide
dehydrogenase and acetyl coenzyme synthase, no passive ‘catalytic’ activity in these minerals has (yet) been demonstrated [28,122, 214, 215, 216,217,218,219]).
Nickel can deputize for iron in greigite, generally in ~Fe5NiS8 or as violarite
~Fe2Ni4S8.[33,212] Yamaguchi and coworkers have demonstrated the electrochemical reduction
of carbon dioxide to formate and methane by coating a carbon paper electrode with violarite.[33] Efficiency, as we might expect, was substantially increased on addition of amine
compounds.[33] Yet, in contrast to its occlusion in mackinawite, molybdenum appears to be excluded from the inverse spinel, though Ni-bearing greigite can be coated with layered MoS2 in
the lab.[223] Dosed thus, Sharifvaghefi and Zheng show that it can, while remaining stable, promote both hydrogenation and desulfurization reactions of, for example, dibenzothiophene (DBT, H4C6S C6H4).[223]
We note here that though never identified as a precipitate in the AHV conditions, the mixed hydroxide and sulfide minerals tochilinite (FeS[Mg,Fe2+][OH]
2) and ferrotochilinite
(6FeS·5Fe(OH)2) could also have been represented in the hydrothermal mound where it might
have played a part in early metabolism.[224,225,226,227] These composite minerals comprise 6 mackinawite-like layers alternating with 5 brucite-like layers.[228] They can be grown below 200°C in alkaline hydrothermal solution, but as a result of the two-dimensional
incommensurability of their sub-structures, they tend to curl into nanotubes.[229] Tochilinite occurs in serpentinized mafic and ultramafic rocks and in CM carbonaceous chondrites where it appears to result from moderate temperature hydrothermal alteration of olivine- and sulfide-rich precursors.[139,228,229,230,231,232,233]
8. The peptide and amyloid takeover?
Barge and her collaborators have demonstrated the amination of pyruvate to alanine in alkaline conditions within moderately reduced green rust.[38] Nevertheless, amino acids have not been polymerized in green rust or mackinawite to date. However, short peptides have been generated in conditions corresponding to the alkaline vent environment.[208,234] And once formed, even six-mer peptides spontaneously and promiscuously sequester inorganic
Fe(Ni)sulfides, metal ions and pyrophosphate (PPi) clusters.[36,189,235,236,237,238,239] Such spontaneous folds around phosphate survive today as the GxxxxGKS/T motif of P-loop
proteins.[240,241,242] The achiral glycines contribute the universal joints required for this coordination, generally referred to as the Walker motif, possibly a reminder of the emergence of an heterochiral world before the left-handed chirality of peptides became dominant?[243,244]
As it is the backbone amide hydrogen bonds that coordinate to the clusters, in these early times the side chains of amino acids played little part beyond, perhaps merely serving to bind two essential metal ions and orient substrates in the so-called “the two-metal-ion catalytic site”.[35,189,245,246,247] The nesting itself renders the clusters more stable, more catalytically active and, in the case of the sulfides, significantly reduces their redox potential. These
properties may have both quickened and directed the early evolution of metabolism. Indeed, these are the structures that were locked into many of the proteins of life as they evolved and complexified.
Many oligopeptides whose residues are sufficiently hydrophobic, will aggregate in water into so-called ‘amyloid’ fibrils. These structures stabilize the oligopeptides against proteolysis and can be capable of conformational self-replication. For example, oligopeptides comprising ala-ile-leu-ser-ser-thr pack as antiparallel beta-sheets that, in spite of having a dry hydrophobic core, can offer water-mediated interfaces, cf. the aquaporin channel.[238,248,249,250]
Greenwald and Reik speculate that this was the common ancestor fold.[249] The sheets are capable of propagation, carrying information, and can be conformationally
self-replicating.[251,252,253,254] Amyloid has also been suggested by many as the likely first organic cell wall and membrane, favored over the leaky and inefficient iron mineral barriers.[238,255] Superior to lipids – which anyway are ‘costly’ to make, suffer the
disadvantage of their impermeability, and likely evolved later – they maintain the capability of proto-enzymetic activity.[133,238,242,256,257]
5 to 6-mer alanine nests sequestering anionic clusters of the iron oxyhydroxides and sulfides, could also have played an essential role in mineral redox-powered disequilibria-converting engines driving ligand-accelerated autocatalytic reactions. In this way
geo-mechanochemical reactions could have been driven along the mineral-enabled pathways toward methanotrophy and acetogenesis, ever improving as peptides sequester active clusters,
eventuating in an incomplete reverse tricarboxylic acid cycle before biochemistry took over proper. Table 1 indicates the possible steps in the putative ligand accelerated pathway to life’s emergence, and Figure 1 details the possible transition from the earth to the life sciences.
10. Emergence of life as a biofilm
We have suggested here that amyloid-forming peptides would be the first organic polymers to be produced within the interlayers of green rust and that these would be secreted to envelop their layered iron mineral parents (Figure 2). We further speculate that pods of amyloid-bearing nanocrysts and peptide nests may have differentiated to protocells of a common size dictated by water content and binding forces. Such cells may have developed different functions, perhaps forming a symbiotic cooperative. In this scenario we might imagine that the intercellular spaces being occupied by amyloid would bind the individual proto-cells together to give
individuated cellular cooperatives.[28,258] Thus, the first palpable living communities, still at the hydrothermal mound, could have been biofilms, adherence then, as it is often now, provided by ‘excess’ amyloid, probably augmented with carboxylates.[259,260,261,262,263,264,265,266] These putative biofilms comprising ‘minimal organic cells’ may have resembled iron-bearing microbialites.[267]
11. Discussion of method and approach
We began this article with a quote from Leduc as a basis for the quest to understand life’s emergence.[1] We quote him again here to give context for this ‘unending’ quest – to remind us that some provisional explanation is required for the emergence of life to give root to the
Darwinian program.[177,268] Leduc insists that “The chain of life is of necessity a continuous one, from the mineral at one end to the most complicated organism at the other. We cannot allow that it is broken at any point, or that there is a link missing between animate and inanimate nature (viz. the missing link between the inorganic and the organic kingdoms). Hence the theory of evolution necessarily admits the physico-chemical nature of life and the fact of spontaneous generation. Only thus can the evolutionary theory become a rational one, a stimulating and fertile inspirer of research."[1] Or more bluntly “Without the idea of spontaneous generation and a physical theory of life, the doctrine of evolution is a mutilated hypothesis without unity or cohesion.”[1]
unknown process. It is mere rubbish thinking, at present, of origin of life; one might as well think of origin of matter” (https://www.darwinproject.ac.uk/letter/DCP-LETT-4065.xml).
It is often overlooked that Darwin’s approach was steeped in the geological milieu he shared with his mentors, Adam Sedgwick and Charles Lyell, two of the founders of modern geology. Indeed, he was a self-proclaimed geologist himself, so it is all the more surprising that earth science is given such short shrift by chemists who rarely consider the initial conditions offered to them by geology.[270,271] Just to report experiments that happen to work in the lab, that seem to mimic certain of life’s pathways, but using unverifiable, indeed, implausible, prebiotic ferrocyanide ‘feedstocks’, leads us to wander into a miasma of uncertainties. It is time to reflect on Polanyi’s premonition: “it is only when we are confronted with the anxious dilemma of a live scientific issue, that the ambiguity of the formal processes and of various attenuated criteria of scientific truth becomes apparent, and leaves us without effective guidance.[272] In other words, we have to think widely and beyond our disciplinary boundaries!
It is true that the notion of‘spontaneous generation’ had fallen into disfavor, where it still resides, since Pasteur’s famous experiment, andDarwin himself sought to distance himself from the idea.[273] Indeed, Deane Butcher recounts how Pasteur’s acolytes prevented publication of Leduc’s hypotheses for some time in France – hypotheses that remain disparaged even in this new century.[1,274] However, it should be remembered that the phrase tends to be understood to support a system supplied with the 'organic building blocks of life’, whereas Leduc’s notion assumed the spontaneous generation to have been in an inorganic milieu.[1] The AHV theory reiterated here also assumes a spontaneous inorganic emergence of life – a theory yet considered as inconsistent with advances made in the laboratory.[275] But again, this is to misunderstand how completely new knowledge is come by.[276] The outcomes of theory and experiments should lead toward palpable goals. In this regard the AHV theory did predict the presence of off-ridge submarine alkaline vents in the present oceans, a prognosis met by the discovery of the Lost City submarine alkaline vents in 2000.[5,12,277] It also explains, for example, why early life did not have to invent such a counterintuitive mechanism as that entailed in Mitchell’s proton motive force, how it was supplied with the necessary low entropy C1 feed, how biosynthesis could proceed in a highly radiated and neutral atmosphere, and why it wasn’t anyway destroyed by surface catastrophes in the Hadean. Here we not only list experiments favoring the theory overlooked by, for example, Ritson and his coworkers, but detail further predictions, some of which could be falsified, reveal embarrassing missing links or even leave the AHVT as just one more casualty of the general theory of ‘natural rejection’! (Table 1).[275,278]. In Feynman’s exacting dictum, here “we are trying to prove ourselves wrong as quickly as possible, because only in that way can we find progress”.[279]
12. Conclusions
monoxide, iv) oxidize hydrogen and methane to methyl groups to react with formate or CO and thereby, v) produce acetate and pyruvate, vi) reduce nitrate, vii) aminate carboxylic acids to the simple amino acids, viii) polymerize these to heterochiral peptides to protect the evolving system at its various scales – all-in-all resulting in the germination and first flowering of the organic evolutionary tree as it emerges from the hydroponically-fertilized green rust seed. The activity of the water generated in these proto-biosynthetic reactions may have been kept low in the hydrous innards of green rust and mackinawite through exosmosis to the salty surrounds.[93,186]
The AHV theory as it stands, focuses on how the hydrous interlayers or channels in green rust, and to a lesser degree in mackinawite, likely mediated the imposed proton gradient and, through the bifurcation of electron pairs, effectively stepped up the redox gradients to the effect of driving an organic takeover.[4] We are conscious that the relative contributions of green rust, mackinawite and associated minerals, and how they may have cooperated in Leduc’s ‘synthetic biology’, has been left unresolved. And we alert the reader to a significant revision of the AHV theory since its first proposal, namely that although we still recognize that the job of life overall is to hydrogenate carbon dioxide, it may be that life first captured both the partially and fully reduced forms of C1 carbon as hydrothermal formate and methane, and only later (though well before LUCA) ‘learnt’ to reduce CO2 through all the required intermediates for CO2 autotrophy
to emerge from its mineral placenta.[50,54,55]
At bottom is the assumption that the protonically and electronically powered nano-engines and disequilibrium conversions needed to drive those endergonic reactions required to produce life’s many processors and its superstructures today were initially coopted from iron oxyhydroxides and sulfides, dosed with transition metals and phosphate, precipitated at the submarine alkaline vent.[13,280,281,282] However, even in the light of some experimental support we can see no clear path to the nucleotide world and, beyond intriguing suggestions of Greenwell and Coveney and others, the theory dissipates in hypotheses, ideas and
speculation.[7,34,283] Nevertheless, “that living organisms are distinguished, not by their momentary appearance, but by their behaviour and by their relationship to their environment” in Mitchell’s memorable phrase, would also be true for the putative layered hydrated seed crystals to life.[284]
So, maybe it all began with the selection of thermodynamically-driven microcrysts which could ‘improve’ (increase the system's dissipative flow) by becoming more organized and
complicated, and/or by growing or declining in ‘size’, and/or by replicating, and, in doing any or all of these the more efficiently, they could have ‘captured’ the driving disequilibria, to the effect of converting those disequilibria ever faster and more efficiently on the path to subjugation by the organic world. But showing how selection for the 'fittest' in this sense can happen, or does happen, in the mineral precipitates is the major challenge?[3,4,55,177]
Acknowledgements
I thank Elbert Branscomb, Wolfgang Nitschke, Grayson Chadwick, Laurie Barge, Stuart Bartlett, Julyan Cartwright, Leon Alkali, Jan Arends, Patrick Beckett, Isik Kanik, Adrian Ponce and Adrian Stoica for discussions. I am also grateful to two anonymous referees‘ for their generous suggestions and corrections. This research was supported by NASA, through the NASA
Conflicts of Interest
The author declares no conflict of interest.
References
1. Leduc, S. The Mechanism of Life, Rebman Ltd., London, 1911.
2. Korenaga, J.; Planavsky, N.J.; Evans, D.A. Global water cycle and the coevolution of the Earth’s interior and surface environment. Phil. Trans. R. Soc. A2017, 375, 20150393. 3. Branscomb, E.; Russell, M.J. Frankenstein or a submarine alkaline vent: Who is responsible
for abiogenesis? Part 1: What is life–that it might create itself? BioEssays2018, 40, 700179. 4. Branscomb, E.; Russell, M.J. Frankenstein or a submarine alkaline vent: Who is responsible
for abiogenesis? Part 2: As life is now, so it must have been in the beginning. BioEssays2018, 40, 700182.
5. Russell, M.J.; Hall, A.J.; Turner, D. In vitro growth of iron sulphide chimneys: possible culture chambers for origin-of-life experiments. Terra Nova1989, 1, 238-241.
6. Branscomb, E.; Russell, M.J. Turnstiles and bifurcators: the disequilibrium converting engines that put metabolism on the road. Biochim. Biophys. Acta2013, 1827, 62-78.
7. Arrhenius, G.O. Crystals and life. Helv. Chim. Acta.2003, 86, 1569-1586. 8. Allmann, R. Doppelschichtstrukturen mit brucitähnlichen Schichtionen [Me(II)1−
xMe(III)x(OH)2]x+. Chimia1970, 24, 99-108.
9. Allmann, R. Das Bindungsprinzip der Doppelschichtstrukturen mit brucitähnlichen Schichtionen. Fortschr. Miner.1971, 48, 24-30.
10. Okumura, T.; Ohara, Y.; Stern, R.J.; Yamanaka, T.; Onishi, Y.; Watanabe, H.; Chen, C.; Bloomer, S.H.; Pujana, I.; Sakai, S.; Ishii, T. Brucite chimney formation and carbonate alteration at the Shinkai Seep Field, a serpentinite-hosted vent system in the southern Mariana forearc. Geochem. Geophys. Geosyst. 2016, 17, 3775-3796.
11. Pisapia, C.; Gérard, E.; Gérard, M.; Lecourt, L.; Lang, S.Q.; Pelletier, B.; Payri, C.E.;
Monnin, C.; Guentas, L.; Postec, A.; Quéméneur, M. Mineralizing filamentous bacteria from the Prony Bay Hydrothermal Field give new insights into the functioning of serpentinization-based subseafloor ecosystems. Front. Microbio. 2017, 57.
12. Kelley, D.S.; Karson, J.A.; Blackman, D.K.; Früh-Green, G.L.; Butterfield, D.A.,; Lilley, M.D.; Olson, E.J.; Schrenk, M.O.; Roe, K.K.; Lebon, G.T.; Rivizzigno, P.; and the AT3-60 Shipboard Party. An off-axis hydrothermal vent field near the Mid-Atlantic Ridge at 30ºN. Nature2001, 412, 145-149.
13. Branscomb, E.; Biancalani, T.; Goldenfeld, N.; Russell, M.J. Escapement mechanisms and the conversion of disequilibria: The engines of creation. Phys. Rep. 2017, 677, 1-6.
14. Russell, M.J.; Hall, A.J. 2006, The onset and early evolution of life. In Kesler, S.E., and Ohmoto, H., eds., Evolution of Early Earth’s Atmosphere, Hydrosphere, and Biosphere— Constraints from Ore Deposits, Geol. Soc. Am. Mem. 2006, 198, 1-32.
15. Nitschke, W.; Russell, M.J. Hydrothermal focusing of chemical and chemiosmotic energy, supported by delivery of catalytic Fe, Ni, Mo/W, Co, S and Se, forced life to emerge. J. Mol. Evol.2009, 69, 481-496.
17. Génin, J.-M.R.; Aïssa, R.; Géhin, A.; Abdelmoula, M.; Benali, O.; Ernstsen, V.; Ona-Nguema, G.; Upadhyay, C.; Ruby, C. Fougerite and FeII–III hydroxycarbonate green rust;
ordering, deprotonation and/or cation substitution; structure of hydrotalcite-like compounds and mythic ferrosic hydroxide Fe(OH)(2+x). Solid State Sci. 2005, 7, 545-572.
18. Wander, M.C.; Rosso, K.M.; Schoonen, M.A. Structure and charge hopping dynamics in green rust. J. Phys. Chem. C2007, 111, 11414-11423.
19. Page, C.C.; Moser, C.C.; Chen, X; Dutton, P.L. Natural engineering principles of electron tunnelling in biological oxidation–reduction. Nature1999, 402, 47-52
20. Russell, M.J.; Daniel, R.M.; Hall, A.J.; Sherringham, J. A hydrothermally precipitated catalytic iron sulphide membrane as a first step toward life. J. Mol. Evol. 1994, 39, 231-243. 21. Mitchell, P. Keilin’s respiratory chain concept and its chemiosmotic consequences. Science
1979, 206, 1148-1159.
22. Herschy, B.; Whicher, A.; Camprubi, E.; Watson, C.; Dartnell, L.; Ward, J.; Evans, J.R.; Lane, N. An origin-of-life reactor to simulate alkaline hydrothermal vents. J. Mol. Evol. 2014, 79, 213-227.
23. Ljungdahl, L.G.; Irion, E.; Wood, H.G. Total synthesis of acetate from CO2. I.
Co-methylcobyric acid and co-(methyl)-5-methoxy-benzimidizolycobamide as intermediates with Clostridium thermoaceticum. Biochem. 1965, 4, 2771-2779.
24. Fuchs, G. Alternative pathways of autotrophic CO2 fixation. In Autotrophic Bacteria, Eds
H.G. Schlegel and B. Bowen, Science Technology, Madison, WI, 1989, 365-382.
25. Fuchs, G. Alternative pathways of carbon dioxide fixation: insights into the early evolution of life. Annu. Rev. Microbiol. 2011, 65, 631-658.
26. Ragsdale, S.W. The Eastern and Western branches of the Wood/Ljungdahl pathway: how the East and West were won. BioFactors1997, 6, 3-11.
27. Russell, M.J.; Martin, W. The rocky roots of the acetyl coenzyme-A pathway. TIBS, 2004, 24, 358-363.
28. Russell, M.J.; Hall, A.J. The emergence of life from iron monosulphide bubbles at a submarine hydrothermal redox and pH front. J. Geol. Soc.1997, 154, 377-402.
29. Windman, T.; Zolotova, N.; Schwandner, F.; Shock, E.L. Formate as an energy source for microbial metabolism in chemosynthetic zones of hydrothermal ecosystems. Astrobiology 2007, 7, 873-890.
30. Sojo, V.; Herschy, B.; Whicher, A.; Camprubi, E.; Lane, N. The origin of life in alkaline hydrothermal vents. Astrobiology, 2016, 16, 181-197.
31. Marchand, C.; Le Calve, S.; Mirabel, P.; Glasser, N.; Casset, A.; Schneider, N.; De Blay, F. Concentrations and determinants of gaseous aldehydes in 162 homes in Strasbourg (France). Atmos. Environ. 2008, 42, 505-516.
32. Jiang, C.; Zhang, P. Indoor carbonyl compounds in an academic building in Beijing, China: concentrations and influencing factors. Front. Environ. Sci. Eng. 2012, 6, 184-194.
33. Yamaguchi, A.; Yamamoto, M.; Takai, K.; Ishii, T.; Hashimoto, K.; Nakamura, R.
Electrochemical CO2 reduction by Ni-containing iron sulfides: How is CO2 electrochemically
reduced at bisulfide-bearing deep-sea hydrothermal precipitates? Electrochim. Acta2014, 141, 311-318.
Thermal Geochemical Processes, R. Ikan ed. Dordrecht, Kluwer Academic Publishers 2003, 325-388.
35. Milner-White, E.J.; Russell, M.J. Nests as sites for phosphates and iron-sulfur thiolates in the first membranes: 3 to 6 residue anion-binding motifs (nests). Orig. Life Evol. Biosph. 2005, 35, 19-27.
36. Milner-White, E.J.; Russell, M.J. Functional capabilities of the earliest peptides and the emergence of life. Genes2011, 2, 671-688.
37. Falkowski, P.G.; Fenchel, T.; Delong, E.F. The microbial engines that drive Earth's biogeochemical cycles. Science2008, 320, 1034-1039.
38. Flores, E.; Barge, L.; VanderVelde, D.; Kallas, K.; Baum, M.M.; Russell, M.J., Kanik, I. Amino acid synthesis in seafloor environments on icy worlds. Am. Astron. Soc. DPS Meeting 2016, 48, id.323.02.
39. Bungs, M.; Tributsch, H. Electrochemical and photoelectrochemical insertion and transport of hydrogen in pyrite, Ber. Bunsenges. Phys. Chem. 1997, 101, 1844.
40. Russell, M.J.; Hall, A.J.; Fallick, A.E.; Boyce, A.J. On hydrothermal convection systems and the emergence of life. Econ. Geol. 2005, 100, 419-438.
41. Huber, C.; Wächtershäuser, G. Primordial reductive amination revisited. Tetrahedron Letters, 2003, 44, 1695-1697.
42. Huber, C.; Wächtershäuser, G. Activated acetic acid by carbon fixation on (Fe,Ni)S under primordial conditions. Science1997, 276, 245-247.
43. Heinz, W.; Lauwers, A.M. Organic sulfur compounds resulting from the interaction of iron sulfide, hydrogen sulfide and carbon dioxide in an anaerobic aqueous environment. Orig. Life Evol. Biosph. 1996, 26, 131-150.
44. Heinen, W.; Lauwers, A. The iron-sulfur world and the origins of life: abiotic thiol synthesis from metallic iron, H2S and CO2; a comparison of the thiol generating FeS/HCl (H2S)/CO2
-system and its Fe0/H
2S/CO2-counterpart. Proc. Koninklijke Nederlandse Akademie van
Wetenschappen, Amsterdam1997, 100, 11-25.
45. Mielke, R.E.; Russell, M.J.; Wilson, P.R.; McGlynn, S.; Coleman, M.; Kidd, R.; Kanik, I., Design, fabrication and test of a hydrothermal reactor for origin-of-life experiments, Astrobiology2010, 10, 799-810.
46. Shibuya, T.; Yoshizaki, M.; Masaki, Y.; Suzuki, K.; Takai, K.; Russell, M.J. Reactions between basalt and CO2-rich seawater at 250 and 350° C, 500bars: Implications for the CO2
sequestration into the modern oceanic crust and the composition of hydrothermal vent fluid in the CO2-rich early ocean. Chem. Geol. 2013, 359, 1-9.
47. Shibuya, T.; Russell, M.J.; Takai, K. Free energy distribution and hydrothermal mineral precipitation in Hadean submarine alkaline vent systems; Importance of iron redox reactions under anoxic conditions. Geochim. Cosmochim. Acta. 2016, 175, 1-19.
48. Reeves, E.P.; McDermott, J.M.; Seewald, J.S. The origin of methanethiol in midocean ridge hydrothermal fluids. Proc. Natl. Acad. Sci. USA2014, 111, 5474-5479.
49. Goldschmidt, V.M. Geochemical aspects of the origin of complex organic molecules on Earth, as precursors to organic life. New Biol.1952, 12, 97-105.
50. Say, R.F.; Fuchs, G. Fructose 1,6-bisphosphate aldolase/phosphatase may be an ancestral gluconeogenic enzyme. Nature2010, 464, 1077-1081.
52. Shock, E.L. Geochemical constraints on the origin of organic compounds in hydrothermal systems. Orig. Life Evol. Biosph. 1990, 20, 331–367.
53. Williams, R.J.P.; Frausto da Silva, J.J.R. The natural selection of the chemical elements.
1996, Clarendon. Oxford.
54. Nitschke, W.; Russell, M.J. Beating the acetyl coenzyme-A pathway to the origin of life. Phil. Trans. R. Soc. Lond. B. Biol. Sci. 2013,368, 20120.
55. Russell, M.J.; Nitschke, W. Methane: Fuel or exhaust at the emergence of life. Astrobiology 2017, 17, 1053-1066.
56. Nealson, K. H.; Belz, A.; McKee, B. Breathing metals as a way of life: geobiology in action. Antonie Leeuwenhoek Int. J. Gen. Mol. Microbiol. 2002, 81, 215-222.
57. Ducluzeau, A-L.; van Lis, R.; Duval, S.; Schoepp-Cothenet, B.; Russell, M.J.; Nitschke, W. Was nitric oxide the first strongly oxidizing terminal electron sink? TIBS 2009, 34, 9-15. 58. Wong, M.L.; Charnay, B.D.; Gao, P.; Yung, Y. L.; Russell, M.J. Nitrogen oxides in early
Earth's atmosphere as electron acceptors for life's emergence. Astrobiology2017, 17, 975-983.
59. Bounama, C.; Franck, S.; von Bloh, W. The fate of the Earth's ocean. Hydro. Earth Syst. Sci. 2001, 5, 569-575.
60.Mojzsis, S.J.; Harrison, T.M.; Pidgeon, R.T. Oxygen-isotope evidence from ancient zircons for liquid water at the Earth's surface 4,300 Myr ago. Nature 2001, 409, 178-181.
61. Valley, J.W.; Lackey, J.S.; Cavosie, A.J.; Clechenko, C.C.; Spicuzza, M.J.; Basei, M.A.S.; Bindeman, I.N.; Ferreira, V.P.; Sial, A.N.; King, E.M.; Peck, W.H. 4.4 billion years of crustal maturation: oxygen isotope ratios of magmatic zircon. Contr. Mineral Petrol.2005, 150, 561-580.
62. Bédard, J.H. A catalytic delamination-driven model for coupled genesis of Archaean crust and sub-continental lithospheric mantle. Geochim. Cosmochim. Acta. 2006. 70, 1188-1214. 63. Bédard, J.H. Stagnant lids and mantle overturns: implications for Archaean tectonics,
magmagenesis, crustal growth, mantle evolution, and the start of plate tectonics. Geosci. Front.2018, 9, 19-49.
64. Zahnle, K.; Arndt, N.; Cockell, C.; Halliday, A.; Nisbet, E.; Selsis, F.; Sleep, N.H. Emergence of a habitable planet. Space Sci. Rev.2007, 129, 35-78.
65. Greenwood, R.C.; Barrat, J.A.; Miller, M.F.; Anand, M.; Dauphas, N.; Franchi, I.A.; Sillard, P.; Starkey, N.A. Oxygen isotopic evidence for accretion of Earth’s water before a high-energy Moon-forming giant impact. Sci. Adv.2018, 4, eaao5928.
66. Beall, A. P.; Moresi, L.; Cooper, C.M. Formation of cratonic lithosphere during the initiation of plate tectonics. Geology2018, 46, 487-490.
67. Daly, R.T.; Schultz, P.H. The delivery of water by impacts from planetary accretion to present. Sci. Adv.2018, 4, eaar2632.
68. Mann, P.; Taira, A. Global tectonic significance of the Solomon Islands and Ontong Java Plateau convergent zone. Tectonophys. 2004, 389, 137-190.
69. Roberge, J.; Wallace, P.J.; White, R.V.; Coffin, M.F. (2005). Anomalous uplift and subsidence of the Ontong Java Plateau inferred from CO2 contents of submarine basaltic
glasses. Geology2005, 33, 501-504.