Expression Systems for
Peptide Production
Susanna Leong School of Chemical and Biomedical Engineering, Nanyang Technological University, Singapore
(Source: Lonza Ltd., Basel, Switzerland)
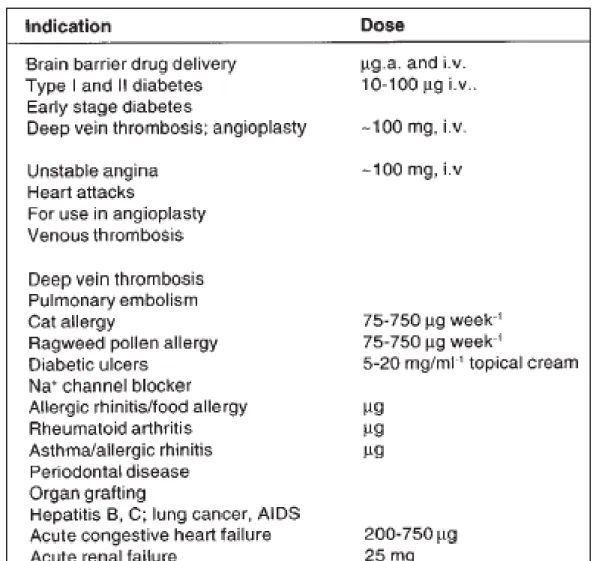
- Demand for peptides to be produced in >100 kgs (industrial).
Challenges in Peptide Production
Table 1: Peptide therapeutics dosage requirement (BioScan).
- Demand for peptides to be produced in >100 kgs (industrial).
- Right strategy of synthesis applicable on all scales. - Quality of peptides.
- Downstream processing and isolation. - Economics/cost-of-goods.
2 main technologies available for peptide production: (i) Solid Phase Synthesis
(ii)Recombinant Technology
Issues with Solid Phase Synthesis - Expensive “raw material”,
- Complex impurity profile,
- Increasing peptide lengths can lead to chain
aggregation, truncated peptides, difficult purification, low yield.
Recombinant Peptide Production
Key advantages:- Relatively cheap and simple to perform. - Easy manipulation of expression host. - Potentially higher yield at a cheaper cost
(with bioprocess-centered peptide design, optimised downstream processing). Raw materials, Waste streams. 35%:65% Solid phase Synthesis Unit operations,
Scale-up, development costs.
5%:95% Recombinant
Technology
Cost-determining Factors Raw Materials:
Production Cost Ratio
Direct secretion – minimise unit operations. Hydrophilic and slightly hydrophobic proteins. With or without 2° or 3° structures. Intracellular or Fermentation medium P. Pastoris Use of secretion to generate biological active conformations Proteins with 2° or 3° structure (S-S bonds). Periplasmic E. coli High product concentrations (5-10 g/L culture) Hydrophilic proteins. No 2° or 3° structure. Intracellular E. coli Advantages Suitable for Product Localisation Host System
E. coli
Vectors for Peptide Expression
- pET: high expression levels and tight control.- pAED - pGEX
Peptide expression in
E. coli
Stability of peptides enhanced if: (i) fused to a carrier protein, OR
(ii) linked together as a large tandem polymer of repeated units.
- Low expression levels,
Tandem Peptide Fusion Protein
Table 2: Production of recombinant KSI-Peptide-His Tag Fusion Proteins (Kuliopulos and Walsh, 1994).
Fusion Protein System for short S-S containing
Peptides
(Fairlie et al., 2002)
SHP2 Met HIS
SS
Peptide
Fig. 1: Intracellular phosphatase (SHP2) fusion construct (Fairlie et al., 2002).
Key advantages of SHP2 as a fusion partner:
(i) Expressed at high levels in E. coli,
(ii) Easily purified via a hexa-histidine tag, (iii) Highly soluble in native buffers,
(iv) Contains a unique Met residue to facilitate cleavage.
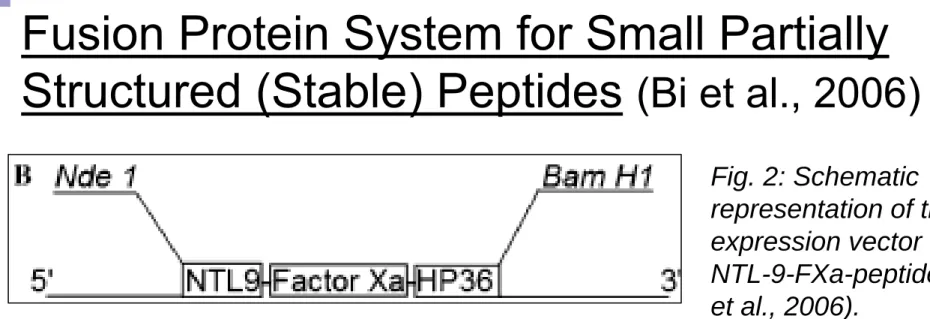
Fusion Protein System for Small Partially
Structured (Stable) Peptides
(Bi et al., 2006)
Key advantages of NTL (N-term domain of L9) as a fusion partner:
(i) Expressed at moderately high levels in E. coli,
(ii) Very soluble and highly stable (melting mid-point of 77 deg C and remains fully folded from pH 1-11),
(iii) Easy purification on ion exchange (protein is very basic).
Purified fusion protein yield: 70 mg/L
Purified peptide yield: ~ 30 mg/L
Fig. 2: Schematic representation of the expression vector for NTL-9-FXa-peptide (Bi et al., 2006).
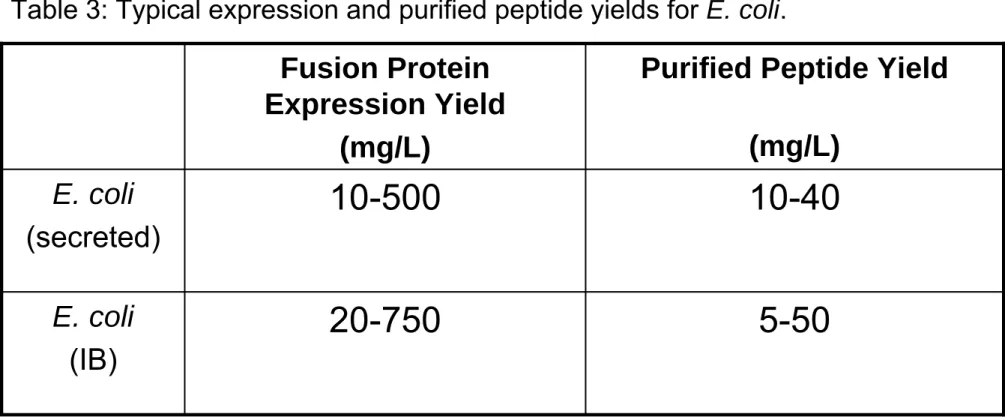
5-50 20-750 E. coli (IB) 10-40 10-500 E. coli (secreted)
Purified Peptide Yield (mg/L)
Fusion Protein Expression Yield
(mg/L)
Yeast Host Systems
Pichia pastoris (P. pastoris)
- Utilises methanol as sole C source (methylotrophic). - AOX1 promoter:
(i) production of alcohol oxidase for methanol oxidation, (ii) over-expression of protein genes.
- Growth to high cell densities on inexpensive, defined media (Typical growth levels: >30% of total protein).
- Foreign protein expression levels range from mg/L to g/L. (10 to 100 times higher protein expression levels than S. cerevisiae).
- Expression levels depend on a.a. sequence, tertiary structure, expression site.
P. pastoris
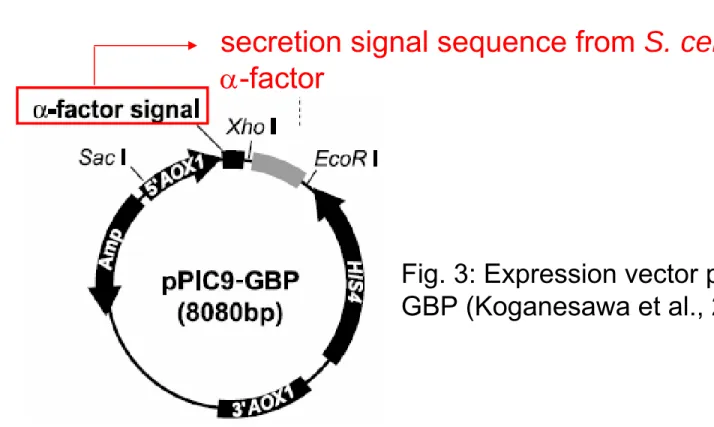
Vector for Peptide Expression
- Allows intracellular expression or secretion into growth medium.
Fig. 3: Expression vector pPIC9-GBP (Koganesawa et al., 2002). secretion signal sequence from S. cerevisiae α-factor
Table 4: Effect of signal sequences on secretion levels and activities of recombinant xylanases produced in P. pastoris (Korona et al., 2006)
Other leader peptides (e.g., signal sequences from S.
cerevisiae secretory proteins) which enhance secretory yields: (i) invertase,
(ii) acid phosphatase, (iii) killer toxin.
- Secretion efficiency of yeast cells influenced by: (i) Host strain and product characteristics,
(ii) Signal/leader sequence, (iii) Promoter strength,
(iv) Expression vectors, (v) Chaperon availability,
(vi) Environmental factors (bioreactor operations, media composition).
Saccharomyces cerevisiae (S. cerevisiae)
- Galactose-inducible promoter (GAL) most efficient, compared to constitutive promoters (e.g., TPI, ADHI).
- GAL: induced by galactose, repressed by glucose.
Advantage over P. pastoris:
- Have stable multicopy vectors (e.g., episomal plasmids) – no screening of multi-copy transformants needed.
Disadvantage over P. pastoris:
- Generally lower secretion efficiency. - Grown at lower cell density.
Case Study: mEGF secretion in P. pastoris vs. S. cerevisiae (Clare et al., 1991) -N.D. 7.7 22.4 13-copy 1.8 6.0 0.6 1.9 Single-copy P. pastoris 3.2 7.4 3.7 0.6 Multi-copy 0.07 0.02 -<0.001 Single-copy S. cerevisiae mg/1011 cells mg/L mg/1011 cells mg/L YP Medium** YNB Medium* Secretion Level Host
mEGF secretion by multi-copy P. pastoris (Clare et al., 1991) 447 402 355 N.D. N.D. 33.6 Fermenter (mg/L) 48.7 35.6 23.2 8.5 2.5 1.5 Shake flask (mg/L) 19 13 9 4 2 1 Copy No.
• Increase in mEGF levels with increased gene dosage was reduced in the fermenter c.f. shake flasks.
• Saturation of secretion pathway was not observed even at high gene dosage.
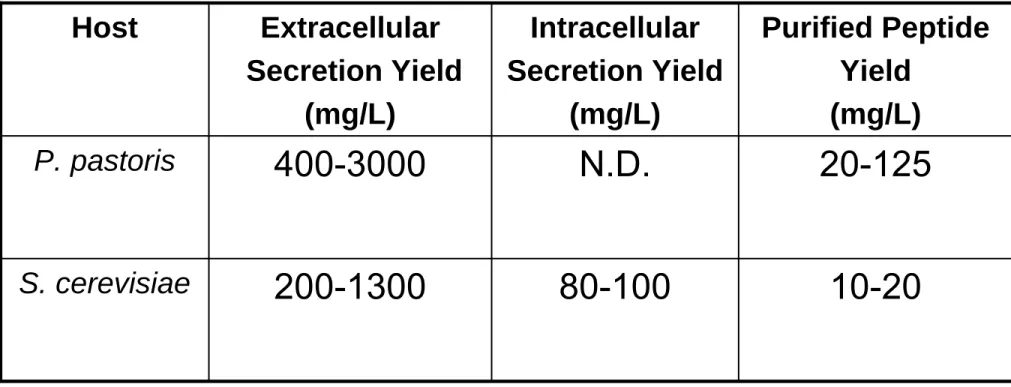
80-100 N.D. Intracellular Secretion Yield (mg/L) 10-20 200-1300 S. cerevisiae 20-125 400-3000 P. pastoris Purified Peptide Yield (mg/L) Extracellular Secretion Yield (mg/L) Host
Table 5: Typical expression and purified peptide yields for P. pastoris and S. cerevisiae.
Fusion systems for peptide expression
Choice of fusion method depends on cleavage methods (chemical or enzymatic).
Chemical Cleavage (CC)
- CC reagents generally recognize single or paired a.a. residues (Useful for short peptides).
- Removal of reagents via a dialysis/buffer exchange step.
Asparagine-glycine bonds at pH 9 Hydroxylamine
Aspartyl-prolyl (or glycine) bond Dilute acid Tryptophan residues N-chloro succinimide, BNPS-skatole Methionine-Xaa bond Cyanogen bromide Cleavage site CC Reagent
Table 6: Purification of anti-fungal peptide (Gavit et al., 2000)
Optimising cleavage site for high recovery
Simultaneous cell lysis and IB fusion protein hydrolysis
(without prior IB solubilisation) with 10% acid (Gavit et al., 2000).
Enzymatic Cleavage
Table 7: Enzymatic
methods for fusion protein cleavage (Flaschel and Friehs, 1993).
Thioredoxin (Trx)
- 11 700 Da, soluble cytoplasmic protein.
- structural similarity to mammalian protein disulfide isomerase (PDI) → folding partner for IB-prone proteins.
- can be expressed as a soluble form to 40% of total bacterial protein.
Example of a fusion thioredoxin-peptide gene:
product factor Xa
cleavage site
- typical soluble yield: 10-20% of total protein (LaVallie et al., 1993; Wilkinson et al., 1995).
Ubiquitin (Ub)
- 76 a.a. residue eukaryotic protein (with roles in protein turnover and stress response).
- Ub fusion proteins have been reported in yeast (McDonnell et al., 1989; Poletti et al., 1992) and E. coli (Butt et al., 1989; Cherney et al., 1991).
- Ub-specific proteases cleave fused proteins from Ub C-term. - Ub fusion partner enhances overall yields and exhibits
Ubiquitin (Ub) fusion systems in
E. coli
Expression level so far:- 20 to 50% of the total cell protein in E. coli.
- Peptide libraries in E. coli as Ub-fusions – 60 mg/L culture (Labean et al., 1992).
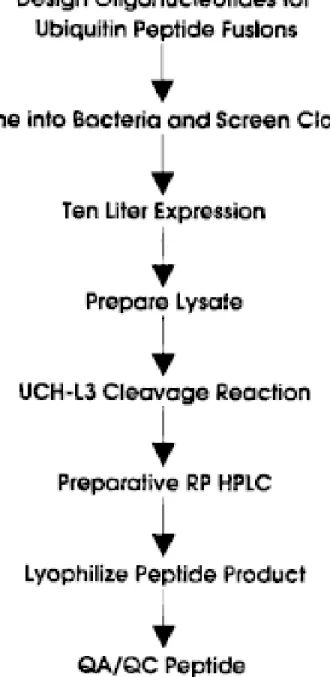
- Multigram peptide yields for 10 L fermentation reported: ~ 360 mg/L pure peptide obtained
Fig. 4: Flow diagram of the Ub fusion process for making recombinant peptides in bacteria (Pilon et al., 1997).
SUMO (
S
mall
U
biquitin-like
MO
difier)
- ~ 100 a.a. residues, heat-stable, highly compact globular structure.
- Enhances peptide expression level, improves solubility and biological activity.
- Addresses problems encountered in most fusion systems (MBP, GST, Trx) where:
(i) Cleavage proteases require linkers between fusion tag and peptide, leading to artificial N-terminus after cleavage.
(ii)Recognition sequences for proteases in linker is small, hence identical a.a. sequences in peptide may also be cleaved.
- highly efficient and recognises the tertiary sequence of SUMO.
SUMO Proteases
SUMO Fusion Systems
Fig. 5: SDS PAGE analysis showing the cleavage of 20 fusion peptides, each
having a different amino acid residue following the cleavage site (LifeSensor Inc.).
- Releases peptides with any desired N-term residue (except proline) → gives desired N-term for enhanced bioactivity.
SUMO Fusion Systems
SUMO proteases cannot tolerate high chaotrope concentrations.
Fig. 6: SDS PAGE analysis showing SUMO protease tolerance against different chemicals (LifeSensor Inc.).
SUMO Fusion System Recovery Flowsheet
Cell lysis Cell debris
Ni- chromatography purification Elute fused peptide
Fused Peptide Cleavage (with SUMO protease)
Ni chromatography purification Remove fusion tag, protease
Concluding Remarks
- In general, mg amounts of purified peptides achievable to date in laboratories.
- P. pastoris can be favourable over E. coli for industrial
production of peptides due to ease of scale up to very high cell densities, high volumetric yields and the absence of endotoxins.
- Characteristics of each peptide determine the optimal host and production strategy.